
Аскомицетов это паразиты или нет
Обновлено: 19.04.2024
Возбудителями отомикозов. Классификация грибов поражающих уши
Грибки — это большая группа бесхлорофильных низших растений, которые в силу этого свойства не способны ассимилировать углекислоту воздуха и получают углерод в виде готовых органических соединений. В природе грибки вместе с бактериями участвуют в круговороте веществ, разлагая некоторые органические вещества. Вегетативное тело грибков, называемое грибницей, или мицелием, состоит из бесцветных или окрашенных нитей — гифов. Гифы представляют собой протоплаз-матическую массу, покрытую оболочкой. В оболочках клеток лишь у очень немногих грибков (оомицетов) содержится целлюлоза. Интересно, что хитин больше ни у каких растений не встречается. Протоплазма мицелия содержит ядра, вакуоли, метахроматические зерна, гликоген, волютин.
Гифы на своем протяжении обычно разделены поперечными перегородками на клетки. Такой мицелий носит название многоклеточного, септированного, в отличие от одноклеточного, нессптированного. Нити мицелия имеют различную толщину и длину, бывают прямыми и волнистыми, нередко параллельно идущие и сближенные гифы образуют так называемые мицелиальные тяжи. Нити мицелия могут давать боковые выросты и соединяться (анастомозировать) между собой.
У некоторых грибков на мицелии иногда имеются специфические образования для прикрепления к субстрату — ризоиды (rhizoides), а также специальные присоски гаустории (haustoria) для высасывания питательных веществ из субстрата.
Размножение грибков может осуществляться вегетативным (бесполым) и половым путем. Споры образуются или в специальных клетках — спорангиях (эндоспоры), или на специализированных гифах — конидиеносцах (экзоспоры).
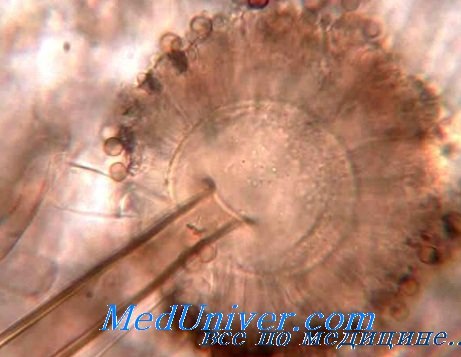
Большинство ботанических классификаций грибов базируется в основном на естественном принципе строения органов полового спороношения. При этом различают пять классов грибов:
1) архимицеты (Archimycetes);
2) фикомицеты (Phycomycetes);
3) аскомицсты (Askomycetes);
4) базидомицеты (Basidiomycetes);
5) несовершенные грибы (Fungi imperfecti).
По способу своего существования все грибки делятся на сапрофитов и паразитов. В настоящее время в мировой литературе известно более 500 видов грибков, патогенных для человека и животных (Е. Gill, 1960). Но это число в значительной мере условно, так как, кроме постоянных паразитов, например дерматофитов, в него входят такие грибки, которые во внешней природе существуют как сапрофиты. При определенных условиях, попадая в организм человека или животного, эти грибки становятся патогенными, так как, приобретая определенную вирулентность, переходят в паразитическое состояние.
Чаще других патогенными являются те грибки, которые входят в состав нормальной микробной флоры кожи и слизистых оболочек человека или животных, а также те, которые находятся в ближайшем их окружении — в воздухе, почве, на фруктах, овощах, стенах сырых помещений и пр.
Возбудителями отомикозов обычно являются именно эти факультативные паразиты и значительно реже — дерматофиты или другие паразитические грибки. Из факультативных паразитов чаще других встречаются представители рода Candida, относящиеся к дрожжеподобным грибкам, а также разные виды аспергиллов и пенициллов, из которых одни относятся к высшим грибам класса Ascomycetes, другие — к несовершенным — Fungi imperfecti, третьи (мукоровые) — к классу низших грибов — Phycomycetes.
В следующих статьях на нашем сайте МедУнивер приводим краткую характеристику тех грибков, которые играют определенную роль в патологии уха.
Аскомицеты (Ascomycetes) (от греч. askos – мешок, сумка и mykes – гриб), один из крупных классов высших грибов, половое размножение которых происходит посредством спор, развивающихся в сумке (аске); то же, что сумчатые грибы.
В учебном пособии приводятся сведения об использовании грибов в промышленности, значении их для человека и в природе.
Раскрыто три типа плодовых тел аскомицетов, на основе которых в классификации выделены 3 подкласса: Голосумчатые, Эуаскомицеты и подкласс Асколокулярные
Подкласс Голосумчатые или гемиаскомицеты – Hemiascomycetidae включает описание, экологическое значение и строение плодовых тел грибов разных порядков и семейств с конкретизацией характеристика разных видов грибов.
В данный подкласс входят грибы Порядка: Эндомицетовые – Endomycetales, рассмотрение которых предложено на примере разных грибов, например, Эндомицетовых, Сахаромицетовых (к которым относятся дрожжевые грибы), Спермофторовых.
Порядок Протомицетовые – Protomycetales – паразиты растений, не имеющие хозяйственного значения. Развиваясь на стеблях, черешках и жилках листьев, они вызывают образование небольших галлов, опухолей или вздутий. Грибы из родов протомицес (Protomyces), тафридиум (Taphridium) и протомикопсис (Protomycopsis) паразитируют исключительно па зонтичных и сложпоцветных, а миксия (Mixia) – на папоротниках.
Порядок Аскосферовые – Ascosphaerales – паразиты насекомых. Грибы единственного вида второго рода – ульевая беттсия (Bettsia alvei) – развиваются в ульях на перге. Широко распространенная в Европе пчелиная аскосфера (Ascosphaera apis) – обычный паразит на личинках медоносной пчелы.
Подкласс Эуаскомицеты – Euascomycetidae образуются в настоящих плодовых телах – аскокарпах, развивающихся по аскогимениальному типу.
Группа порядков Плектомицеты – группа примитивных аскомицетов, имеющих ангиокарпные плодоношения без остиоли, вся полость которых выполнена аскогенными гифами, дающими шаровидные аски, без парафиз и других нитей. Аски неправильно разбросаны в псевдопаренхиматозной ткани. Группа объединяет сумчатые грибы пор. Eurotiales, Microascales, Onygenales.
Порядок Эуроциевые – Eurotiales – объединяет несколько сотен видов. Плодовые тела эуроциевых – клейстотеций с беспорядочно расположенными внутри прототуникатными сумками – образуются обычно на мицелии на поверхности субстрата или бывают погружены в него. Лишь у немногих представителей этой группы они развиваются в стромах, обычно напоминающих склероции. Достаточно полно характеризуется гриб пеницилл, имеющий большое значение и в природе, и в жизни человека.
Порядок Онигеновые – Onygenales – сапрофиты, обитающие на остатках животного происхождения, содержащих кератин, например онигена (Onygena), или на древесине – трихокома (Trichocoma) и дендросфера (Dendrosphaera).
Порядок Микроасковые – Microascales относится к небольшому порядку грибов с темноокрашенными перитециями, содержащими беспорядочно расположенные прототуникатные сумки. Перитеции образуются на поверхности субстрата или частично в него погружены. Они мелкие, шаровидные или грушевидные, часто с длинным хоботком, в несколько раз превышающим по длине диаметр перитеция (Ceratocystis).
Группа порядков Пиреномицеты. Плодовые тела грибов этой группы порядков – перитеции, реже клейстотеции. Освобождение аскоспор активное у перитециальных грибов и пассивное – клейстотециальных.
Группа подпорядков Пиреномицеты включает 5 порядков, среди которых мучнисторосяные, сферейные, спорыньевые и др. Порядки вбирают в себя 640 родов и до 10 тысяч видов, являющихся сапрофитами или паразитами.
Порядок Эризифовые, или мучнисторосяные, грибы – Erysiphales своеобразная группа грибов-паразитов. Большинство их живет на поверхности различных органов растений, где развивается их белая, часто затем темнеющая, грибница со спороношениями. Питание мучнисторосяных грибов осуществляется при помощи гаусториев, проникающих в ткани растений. В этот порядок относятся мучнисторосые грибы. Мучнистая роса яблони, редко груши (Podosphaera leucotricha) поражает все органы растений, на которых появляется белый, затем чуть желтеющий налет. При сильном развитии болезни листья становятся значительно меньше здоровых, буреют и засыхают. Рост пораженных побегов резко замедляется.
Порядок Сферейные – Sphaeriales часто встречаются в огромном количестве в самых различных местообитаниях. Преимущественно это сапрофиты, хотя встречаются и паразиты, иногда даже очень опасные. Однако большинство из этих столь нередких грибов малозаметны.
Порядок Гипокрейные объединяет более 100 родов. Для них характерны мягкие или мясистые перитеции, светлые или ярко окрашенные. Они образуются на мицелии на поверхности или внутри субстрата, на мицелиальном сплетении (субикулюме) или на стромах такого же характера, как и перитеции.
Порядок Гелоциевые – Helotiales характеризуются сумками, которые открываются не в виде крышечки на верхушке, как сумки пецицевых грибов, а в виде трещины или поры.
Порядок Фацидиевые – Phacidiales развивается на растительном опаде, засохших ветвях и листьях древесных растений, кустарников и кустарничков, а также на травянистых и высших споровых растениях. Некоторые грибы поражают зеленые органы растений, а также луб коры и являются причиной их отмирания, что часто приводит к массовой гибели молодых растений в питомниках, культурах и в лесу.
Порядок Циттариевые – Cyttariales- маленький порядок циттариевых объединяет дискомицеты, апотеции которых погружены в студенистые или мясистые стромы. Обычно считают, что сумки у представителей этой группы иноперкулятные, как у фацидиевых и гелоциевых, но у одного из них на сумках обнаружена крышечка, как у саркосцифовых грибов (семейство Sarcoscyphaceae) из порядка пецицевых.
Порядок Пецицевые – Pezizales. Для пецицовых характерны оперкулятные сумки, открывающиеся на вершине крышечкой. Плодовые тела пецицевых – апотеции типичного строения, от очень мелких, не превышающих в диаметре 1 мм, до крупных, размером около 10 см.
Порядок Трюфелевые – Tuberales относятся к группе гипогейных (подземных) аскомицетов. Этот порядок включает около 100 видов, характерный признак которых – подземные плодовые тела, называемые обычно трюфелями. Особое положение среди эуаскомицетов занимает порядок лабульбениевых.
Подкласс Асколокулярные, или локулоаскомицеты – Loculoascomycetidae. Асколокулярные грибы отличаются от других аскомицетов тем, что сумки у них развиваются не в типичных перитециях, а в стромоподобных образованиях, называемых аскостромами или псевдотециями. Формирование сумок у них происходит среди ткани плодовых тел в полостях (локулах), которые возникают частично за счет разрушения внутренней псевдопаренхимы этой стромы, частично за счет вытеснения ее аскогенными гифами и сумками
Порядок Капнодиальные – Capnodiales вызывают явление черни, которая образует черный налет мицелия на листьях и стеблях растений. Капнодиальные грибы развиваются за счет выделений тлей и поэтому не являются паразитами растений, на которых они живут. Однако черный налет мицелия вызывает уменьшение фотосинтетической деятельности соответствующих высших растений. Большинство представителей порядка капнодиальных произрастает в странах теплого климата. В умеренной зоне земного шара встречаются лишь некоторые грибы, относящиеся к роду капнодиум (Capnodium).
Порядок Гистериальные – Hysteriales. Плодовые тела представителей порядка гистериальных – гистеротеции, вытянутые, почти эллипсоидальные, открывающиеся продольной щелью. Сумки булавовидные или цилиндрические, окруженные псевдопарафизами. Споры эллипсоидальные или веретеновидные, двух или многоклеточные, в некоторых родах с продольными и поперечными перегородками, бесцветные или бурые. Порядок гистериевых подразделяют на 4 семейства: гистериевых (Hysteriaceae), графидовые (Graphidaceae), артониевые (Arthoniaceae), рокцелловые (Roccellaceae). Из перечисленных семейств наиболее изучено семейство гистериевых, представители которого относятся к числу весьма распространенных. Представители порядка гистериальных – сапрофиты, развивающиеся, главным образом, на отмерших веточках древесных и кустарниковых растений, а изредка и на стеблях травянистых растений.

Основной признак аскомицетов — образование в результате полового процесса сумок (или асков) — одноклеточных структур, содержащих фиксированное число аскоспор, обычно 8 (рис. 50). Сумки образуются или непосредственно из зиготы (у низших аскомицетов), или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, а затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. У высших аскомицетов сумка представляет не только место образования аскоспор, но и активно участвует в их распространении.
Вегетативное тело аскомицетов — разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки (септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. В центре септы остается пора (рис. 51), через которую происходит движение цитоплазмы с со скоростью от 1—2 до 25—40 см/ч. Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста.

У некоторых аскомицетов мицелий может распадаться на отдельные клетки или почковаться. У дрожжей (порядок Endomycetales) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися, реже делящимися клетками, иногда образующими псевдомицелий. Дрожжеподобный рост наблюдается и у некоторых мицелиальных аскомицетов, например у грибов-дерматофитов (порядок Eurotiales), тафриновых (порядок Taphrinales), некоторых видов из рода цератостис (порядок Microascales).
Высокоспециализированные экзопаразиты насекомых из порядка лабульбениевых (Laboulbeniales) имеют редуцированный мицелий, а их тело (рецептакл) состоит из настоящей ткани.
В состав клеточных стенок аскомицетов, как и у хитридиомицетов и зигомицетов, входит хитин, но его содержание у грибов этого класса ниже и составляет не более 20—25% полисахаридов клеточной стенки (для сравнения: у некоторых хитридиомицетов — до 60%, у зигомицетов — до 37—40%). У некоторых дрожжей (род Schizosaccharomyces) хитин не обнаружен. Большую часть полисахаридов клеточной стенки аскомицетов (80—90%) составляют глюканы — полимеры D-глюкозы, отличающиеся от целлюлозы характером связи между мономерами. У дрожжей, кроме глюканов, обнаружены маннаны — полимеры маннозы. Целлюлоза у аскомицетов не обнаружена, за исключением двух видов из рода цератоцистис.
В цикле развития аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения аскомицетов очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды). Типы конидиальных спороношений и различные способы образования конидий подробно онисапы в главе о дейтеромицетах, или несовершенных грибах.
Конидиалыгые спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов — паразитов растений они обычно образуются на живом растении, а сумчатые спороношения, за немногими исключениями,— после отмирания растения или его частей, в конце периода вегетации или после перезимовки.
У некоторых аскомицетов конидиальное спороношение неизвестно, у других оно преобладает в цикле развития. В отдельных группах этого класса наблюдается редукция полового процесса и сумчатая стадия образуется редко. Иногда ее трудно обнаружить в природе и получить в искусственной культуре грибов. Поэтому многие аскомицеты как в природе, так и в коллекциях культур мы чаще встречаем в их конидиальной стадии. Это отражается и на их названиях. Конидиальные стадии многих аскомицетов имеют самостоятельные видовые наименования, и в литературе мы чаще встречаем их именно под этими названиями, а не под названиями сумчатых стадий. Например, широко используемый в генетических исследованиях аскомицет Emericella nidulans более известен как Aspergillus nidulans. Распространенный в почве и на различных целлюлозосодержащих субстратах аскомицет Hypocrea rufa обычно называют Trichoderma viride по его конидиальиой стадии. Особенпо распространено это для фитопатогенных аскомицетов: название Fusarium solani вместо Nectria haematococca, Helminthosporium sativum вместо Cochliobolus sativus и т. д. В Международном кодексе ботанической номенклатуры, который регулирует употребление названий растений, содержится даже специальный пункт, разрешающий использовать наряду с основным видовым названием аскомицета (по его сумчатой стадии) название его конидиальной стадии в тех случаях, когда речь идет именно об этой стадии гриба. Это единственный случай, когда для разных стадий в цикле развития одного организма допускаются самостоятельные видовые названия.
Для большого числа грибов, встречающихся в природе в гаплоидной конидиальной стадии, половые стадии неизвестны. Такие грибы относят к классу дейтеромицетов, или несовершенных грибов.
Половой процесс, типичный для аскомицетов,— гаметапгиогамия, т. е. слияние двух гаметангиев — специализированных клеток, не дифференцированных на гаметы.
У низших аскомицетов (подкласс Hemiascomycetidae) половой процесс сходен с зигогамией у зигомицетов. Гаметангии разного пола морфологически сходны или малоразличимы и представляют выросты или веточки мицелия. После их слияния сразу происходит кариогамия и сумка развивается непосредственно из зиготы. Однако, в отличие от зигомицетов, в многоядерных гаметангиях сливаются только два ядра (нет множественной кариогамии), зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития низших аскомицетов, следовательно, есть только гаплоидная и диплоидная стадии (рис. 52).

Для высших аскомицетов (подклассы Euascomycetidae и Loculoascomycetidae) характерны дифференциация и усложнение строения гаметангиев. Образуются одноклеточный антеридий и аскогон, обычно с трихогиной. При оплодотворении содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются сразу, а объединяются попарно, образуя дикарионы. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Аскогенные гифы ветвятся и разделяются на двухъядерные клетки. На концах аскогенных гиф развиваются сумки (см. рис. 50).
Конечная клетка аскогенной гифы загибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остается в месте перегиба крючка, одно ядро переходит в его кончик, а другое — в основание. Затем образуются две перегородки, отделяющие одноядерные конечную и базальную клетки крючка. В результате слияния этих клеток восстанавливается дикарион и может происходить повторное образование крючка. Средняя двухъядерная клетка крючка развивается в сумку. Она увеличивается в размерах, ядрадикариона сливаются. Образовавшееся диплоидное ядро делится редукционно, за мейозом следует еще одно, митотическое деление, и вокруг восьми гаплоидных ядер формируются аскоспоры.
Образование аскогенпых гиф увеличивает число сумок, а следовательно, и аскоспор, развивающихся из одного аскогона. Образование сумок по способу крючка с восстановлением дикариона в его базальной клетке и повторным образованием крючков обеспечивает расположение сумок пучком или слоем, что очень важно при активном освобождении аскоспор.
У некоторых аскомицетов (например, из порядка эуроциевых) сумки образуются на аскогенных гифах иными способами — цепочками по ходу аскогенных гиф, из их конечных клеток или боковых выростов.
В цикле развития высших аскомицетов чередуются, таким образом, три стадии: длительная — гаплоидная, в течение которой происходит бесполое размножение, непродолжительная — дикарионтическая (аскогенные гифы) и очень короткая — диплоидная (молодая сумка с диплоидным ядром).
Для многих аскомицетов характерна морфологическая редукция полового процесса. У некоторых представителей этого класса антеридии отсутствуют или не функционируют. В этом случае их функции могут выполнять конидии, вегетативные гифы, а часто мелкие специализированные клетки, называемые спермациями. Спермации нередко образуются на другой особи, на значительном расстоянии от аскогона и переносятся на трихогину токами воздуха, дождем, насекомыми. Трихогина некоторых аскомицетов хемотропична и подрастает к спермациям или конидиям совместимого типа (Ascobolus stercorarius, Podospopa anserina, виды Neurospora). Сперматизация обнаружена в разных группах аскомицетов, например у некоторых гелоциевых, видов из родов Ascobolus и Gelasinospora, в порядке лабульбениевых и у других.
В том случае, когда отсутствуют оба гаметангия, образование дикариона происходит в результате слияния клеток обычных вегетативных гиф одного или двух совместимых мицелиев — соматогамии (например, у Sclerotinia sclerotiorum).
Среди аскомицетов есть гомоталличные и гетероталличные виды, причем гетероталлизм здесь всегда биполярный (двухаллельного типа).
В сумке аскомицетов обычно образуется 8 аскоспор. Однако наблюдаются многочисленные отклонения от тина: например, происходит образование только четырех гаплоидных ядер и соответственно четырех аскоспор (Endomyces magnusii) или часть образовавшихся ядер дегенерирует и число аскоспор за счет этого сокращается (Verpa bohemica). Образование большего числа аскоспор происходит или при большем числе делений ядра (например, у Podospora — семь делений ядра и соответственно 128 аскоспор в сумке), или в результате почкования аскоспор в сумке (например, у тафриновых), или их распада на клетки (у некоторых гипокрейных).
Форма аскоспор очень разнообразна — от шаровидных или эллипсоидальных до нитевидных (Rhytisma, Claviceps). Аскоспоры могут быть одноклеточными или иметь поперечные перегородки, реже — поперечные и продольные перегородки (муральные споры).-
У некоторых аскомицетов аскоспоры имеют разнообразные придатки, играющие роль в их распространении, например слизистые придатки аскоспор копрофильных грибов из рода Podospora или аскоспор сапрофитных морских аскомицетов Ceriosporiopsis halima, Remispora maritima и др.
По строению оболочки и функциям сумки аскомицетов делят на две большие группы — прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую недифференцированную оболочку, которая разрушается или растворяется, освобождая аскоспоры пассивно. Такая сумка служит только местом формирования спор, но не участвует активно в их распространении. Эутуникатные сумки характеризуются более плотными оболочками, часто со специальными приспособлениями для вскрывания сумки. Они активно участвуют в распространении аскоспор (активное выбрасывание спор из сумки). По строению оболочки эутуникатные сумки могут быть двух типов: унитуникатные и битуникатные. У унитуникатных сумок оболочка относительно тонкая и выглядит однослойной, на вершине сумок имеется обычно апикальный аппарат различного строения, служащий для их вскрывания. Битуникатные сумки имеют ясновыраженную двухслойную оболочку, состоящую из жесткого наружного и эластичного внутреннего слоев. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины; внутренний слой под действием повышенного тургорного давления растягивается, и происходит активное выбрасывание аскоспор.
Форма сумок округлая или овальная (у гемиаскомицетов и аскомицетов с замкнутыми плодовыми телами) или цилиндрическая (у аскомицетов с активным выбрасыванием аскоспор).

У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших — в специальных вместилищах — плодовых телах (рис. 69). Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидпое плодовое тело с отверстием на вершине ) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеций, перитеции и апотеции — настоящие плодовые тела, развитие которых происходит по аскогимениальному типу — образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.
Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах — плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела.
Кроме настоящих плодовых тел, аскомицеты образуют аскостромы, развитие которых происходит по асколокулярному типу. Сначала закладывается строма из переплетающихся гиф. В ней образуются аскогоны и происходит половой процесс. Аскогенпые гифы и образующиеся на них сумки раздвигают или разрушают плектенхиму стромы, освобождая в ней полость — локулу. Каждая локула содержит одну или несколько сумок. Ткань стромы над локулой разрушается и образуется отверстие, через которое освобождаются аскоспоры. По внешнему виду аскостромы часто похожи на настоящие перитеции, но отличаются от них отсутствием собственного перидия — их оболочкой служит плектенхима стромы.
На основании отсутствия или наличия плодовых тел и способов их образования класс аскомицеты делят на три подкласса:
Подкласс Голосумчатые, или Гемиаскомицеты (Hemiascomycetidae). Плодовые тела отсутствуют. Сумки образуются непосредственно на мицелии, прототуникатные.
Подкласс Эуаскомицеты (Euascomycetidae). Сумки образуются в клейстотециях, перитециях или апотециях, прототуникатные или унитуникатные.
Аскомицеты широко распространены в природе во всех географических областях, на всех доступных для грибов субстратах. Они обитают как сапрофиты в почве, в лесной подстилке, на разнообразных растительных субстратах (древесина, отмершие растения и т. п.). Некоторые группы аскомицетов заняли своеобразные экологические ниши, недоступные для других грибов. Например, кератинофильные грибы, развивающиеся на различных субстратах животного происхождения, содержащих кератин. Некоторые аскомицеты обитают в морях или пресных водоемах на погруженной в воду древесине. Сапрофитные аскомицеты активно участвуют в минерализации органических веществ в природе, особенно в разложении растительных остатков, содержащих целлюлозу. Многие сапрофитные аскомицеты вызывают плесневение и порчу разнообразных материалов и изделий, а также пищевых продуктов.
Многочисленные аскомицеты паразитируют на различных организмах — на растениях (грибах, водорослях, лишайниках и высших), а также на животных и человеке, нередко вызывают серьезные заболевания.
Многие представители этого класса имеют большое экономическое значение как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), витаминов (рибофлавина и др.), ферментов, кормового белка, а также как возбудители спиртового брожения. Наконец, многие аскомицеты широко используются сейчас в качестве объектов генетических и биохимических исследований.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .

Аскомицеты (от греч. ἀσκός — сумка), или сумчатые грибы (лат. Ascomycota ) — отдел в царстве грибов, объединяющий организмы с септированным (разделённым на части) мицелием и специфическими органами полового спороношения — сумками (асками), содержащими чаще всего по 8 аскоспор. Имеют и бесполое спороношение, причём во многих случаях половой процесс утрачивается и такие виды грибов относят к несовершенным грибам (Deuteromycota).
К аскомицетам относят до 2000 родов и 30 000 видов. Среди них — дрожжи (класс Saccharomycetes) — вторично одноклеточные организмы. Из других известных представителей аскомицетов можно назвать сморчки, пармелию, строчки и трюфели.
Содержание
Морфология
Мицелий разделён поперечными перегородками (септирован), что позволяет гифе в случае повреждения терять меньше клеточного содержимого и обуславливает большую выживаемость аскомицетов по сравнению с ценоцитными зигомицетами. Так, наименьший жизнеспособный фрагмент мицелия, способный дать начало новому организму, для зигомицета составляет около 100 мкм, а для аскомицета — 15—20 мкм (1—3 целых клетки). В середине септы имеется простая пора, через которую возможен обмен цитоплазмой, органеллами и даже ядрами. Чаще всего у аскомицетов одно гаплоидное ядро в клетке.
Размножение и цикл развития
У аскомицетов выражены две стадии развития: анаморфа, в которой организм размножается бесполым путём, и телиоморфа, в которой он формирует половые структуры. Вместе обе стадии носят название голоморфа. Анаморфа и телиоморфа не схожи, в зависимости от условий организм может переходить в телиоморфу, а может и нет, в результате чего часто один и тот же аскомицет описывался как два разных вида. Например, название Aspergillus fisheri дано анаморфе, телиоморфа которой описана как Neosartoria fisheri, аналогичная ситуация с Amorphotheca resinae и Hormoconis resinae и многими другими. В наши дни молекулярные методы позволяют обнаружить описанные дважды виды.
Бесполое размножение
Бесполое размножение аскомицетов осуществляется практически всегда конидиями. Морфология конидий очень разнообразна. Они могут быть одноклеточными, с разным числом поперечных перегородок, с продольными и поперечными перегородками (муральными). Чаще всего конидии эллипсоидные, могут быть округлыми. Особенно сложное строение имеют конидии некоторых водных грибов, обитающих, в частности, на погружённых в воду разлагающихся листьях листопадных пород деревьев и кустарников.
Конидии могут быть бесцветными или окрашенными обычно в коричневые тона из-за наличия меланинов. Они могут быть сухими или погружёнными в слизь. Сухие конидии распространяются токами воздуха, слизистые — водой (часто с каплями дождя) или мелкими животными (например, перенос жуками-заболонниками конидий Ophiostoma ulmi — возбудителя голландской болезни вязов). При низкой влажности и высыхании слизи такие конидии переносятся также токами воздуха. Освобождение конидии обычно пассивное.
Конидии иногда развиваются непосредственно от мицелия, но гораздо чаще от специальных конидиеносцев, которые могут быть простыми или ветвящимися, иногда очень обильно. Образуются конидии поодиночке или группируются в цепочки или головки. Конидиеносцы также могут располагаться поодиночке или тем или иным образом группироваться. Простейший вариант такой группировки — коремия: конидиеносцы развиваются тесно сближенным пучком, чаще склеиваясь своими боками, а иногда анастомозируя. Коремии характерны, например, для анаморф некоторых Eurotiales (в частности, представителей формального рода Penicillium, причём у некоторых видов могут образовываться только при определённых условиях).
У некоторых видов конидиеносцы образуют слой на поверхности выпуклого сплетения гиф или стромы в виде подушечек. Такой тип спороношения — спородохий. Наиболее частый и характерный пример — анаморфа Nectria cinnabarina.
Ложа по характеру группировки конидиеносцев напоминают спородохии, но конидиеносцы в них образуют тесный слой не на выпуклой строме, а на более или менее плоском сплетении гиф. Такие структуры часто встречаются у грибов, паразитирующих на растениях. Сначала они развиваются в ткани растения под кутикулой или эпидермисом, а затем прорывают их и выступают наружу.
Если расположенные слоем на рыхлом сплетении гиф конидиеносцы погружены в общую слизь, то такие спороношения называют пионноты.
Наиболее сложные конидиальные структуры — пикниды. Они имеют шаровидную или кувшиновидную форму, одеты плотной светлой или тёмной оболочкой с узким отверстием на вершине. Внутри пикниды плотным слоем образуются короткие конидиеносцы, на которых развиваются конидии, выходящие затем из пикниды через выводное отверстие или трещины в оболочке, часто в массе слизи. Иногда даже конидиеносцы могут не дифференцироваться, а конидии образуются непосредственно от клеток оболочки пикнид.
Формирование конидий из конидиогенных клеток может происходить разными способами. Выделяют два типа:
- Таллический. Конидии образуются в результате видоизменения ранее существовавшей части мицелия, иногда имеющей специфический облик (так что получается конидиеносец). Артроконидии (артроспоры) образуются в результате фрагментации конидиеносца или гифы. Алевриоконидии (аревлиоспоры) формируются из отделившейся перегородкой части конидиогенной клетки, разрастающейся и дифференцирующейся в зрелую конидию. Они образуются одиночно на вершине конидиеносца или гифы и часто имеют крупные размеры и утолщённую клеточную стенку. Иногда они развиваются в коротких цепочках.
- Бластический. Зачаток конидии заметно увеличивается в размерах до своего отделения перегородкой от конидиогенной клетки. На основании участия стенки конидиогенной клетки в формировании стенки конидии различают три типа бластогенных конидий: холобластические, пороконидии и фиалоконидии.
У холобластических бластоконидий все слои клетки конидиогенной клетки участвуют в образовании стенки конидии. Бластоконидия развивается как вздутие конца конидиогенной клетки, затем отшнуровывается и отделяется от неё перегородкой (процесс напоминает почкование у аскомицетных дрожжей).
В формировании клеточной стенки пороконидий принимают участие только внутренние слои клеточной стенки конидиогенной клетки. Такие конидии образуются путём почкования через поры в стенках конидиеносцев. Пороконидии обычно толстостенные, располагающиеся по одной на вершине и по бокам конидиеносца.
При формировании фиалоконидий клеточная стенка формируется заново, а стенка конидиогенной клетки не участвует в её возникновении. Фиалоконидии развиваются на фиалидах — конидиогенных клетках, обычно несколько утолщённых у основания и слегка оттянутых в верхней части.
У аскомицетов, паразитирующих на растениях, конидиальные спороношения обычно образуются на живом растении, а сумчатые — после отмирания растения или его частей в конце периода вегетации или после зимовки.
Половое размножение
Половое размножение аскомицетов проходит в форме гаметангиогамии, без образования дифферинцированных гамет. Для этого на разных гифах образуются половые органы: мужские — антеридии и женские — архикарпы, представляющие собой видоизменённые оогонии.
У низших форм половой процесс напоминает зигогамию у зигомицетов (Zygomycota), поскольку образующиеся гаметангии внешне сходны и после их объединения сразу происходит слияние ядер. Однако у низших асковых грибов сливаются только два ядра многоядерных гаметангиев, а не все, то есть отсутствует множественная кариогамия. Образовавшееся диплоидное ядро без периода покоя делится мейотически, образуя гаплоидные ядра, а зигота трансформируется в сумку — аск (др.-греч. ἀσκός — мешок).
У высших форм половой процесс проходит более сложно. Архикарп у них дифференцирован на расширенную нижнюю часть — аскогон и верхнюю в виде изогнутой трубочки — трихогину. Антеридий представляет собой одноклеточную структуру цилиндрической формы. При соприкосновении гаметангиев трихогина врастает своим концом в антеридий, после чего содержимое антеридия перетекает по ней в аскогон. Слияние цитоплазмы гаметангиев (плазмогамия) не сопровождается слиянием их гаплоидных ядер, хотя они сближаются и располагаются попарно, образуя дикарионы. Затем из аскогона вырастают аскогенные гифы, одновременно ядра дикарионов делятся, что ведёт к увеличению числа дикарионов. В каждую из аскогенных гиф проникают дикарионы. Завершается процесс образованием сумок (асков), которыми становятся концевые клетки, расположенные на концах аскогенных гиф, содержащие дикарион. Вначале гаплоидные ядра дикариона сливаются (происходит кариогамия), образуя диплоидное ядро. Без периода покоя это ядро делится мейотически, образуя четыре гаплоидных ядра, а те, в свою очередь, делятся митотически. В итоге появляются восемь гаплоидных клеток, которые становятся аскоспорами, а клетка, в которой они находятся, становится аском. У некоторых видов количество спор может быть меньшим (четыре) за счёт отсутствия митотического деления после мейотического или за счёт дегенерации части гаплоидных ядер или большим.
Таким образом, в жизненном цикле асковых грибов имеется три стадии: гаплоидная стадия, когда мицелий размножается бесполым путём; стадия дикариона и самая короткая — диплоидная стадия, когда молодая сумка непродолжительное время содержит диплоидное ядро.
У многих сумчатых грибов половой процесс упрощается. У них не образуются антеридии, а вместо них функционируют вегетативные гифы, конидии или мелкие клетки спермации, которые могут образовываться даже не на соседних гифах, а на достаточном расстоянии от архикарпа. Потоками воздуха, воды или через насекомых спермации переносятся на трихогину, после чего происходит слияние цитоплазмы клеток. Случается, что могут отсутствовать оба гаметангия, тогда половой процесс протекает в форме соматогамии, то есть сливаются вегетативные клетки. При этом следует отметить, что у сумчатых грибов есть формы с гомоталличным и гетероталличным мицелиями, причём последние всегда отличаются набором аллелей.
Аскогенные гифы с асками могут образовываться не только беспорядочно (в любом месте мицелия), как это происходит у низших форм, но и на плодовых телах, состоящих из плотно переплетённых гиф. У сумчатых грибов имеются четыре типа плодовых тел:
Псевдотеции характерны для класса Loculoascomycetes. При их формировании вначале образуются мицелиальные стромы, в полостях которых формируются гаметангии, происходит половой процесс и образование сумок. Клейстотеции и перитеции часто развиваются на особых образованиях — стромах — плотных сплетениях гиф, которые часто ошибочно принимают за плодовые тела. Они могут находиться на поверхности стромы или быть погружёнными в её тело. Стромы могут быть довольно простого строения, как у видов рода Hypoxylon, а могут представлять собой сложные гифенные образования, например дифференцированные на стерильную ножку и фертильную верхнюю часть стромы видов родов Claviceps и Cordyceps.
Настоящие плодовые тела могут развиваться также непосредственно на мицелии, на рыхлом его сплетении — субикулуме.
Развитие аскостром идёт по-другому. Сначала строма гомогенная. В ней образуются половые структуры, происходит половой процесс, формируются аскогенные гифы и сумки. При этом ложная ткань (плектенхима) стромы в местах образования сумок раздвигается или разрушается, и там образуется полость — локула. Каждая локула может содержать всего одну сумку, но чаще сумок в ней достаточно много. На вершине локулы из-за разрушения ткани обычно формируется выводное отверстие. Внешне аскостромы могут становиться очень похожими на настоящее плодовое тело или их группу.
Сумки
Сумки могут быть разной формы — от округлых или овальных до сильно вытянутых, булавовидных или цилиндрических. По строению оболочки и функциям их можно разделить на прототуникатные и эутуникатные (эвтуникатные). У первых оболочка тонкая, недифференцированная, она разрушается или лизируется, а аскоспоры высвобождаются пассивно. У вторых оболочка более плотная, часто со специальными приспособлениями для вскрытия, и освобождение спор активное.
В свою очередь, эутуникатные сумки можно подразделить на унитуникатные и битуникатные. И те и другие имеют оболочку из двух слоёв, однако у унитуникатных сумок оболочка тонкая, выглядит однослойной, и слои её неразделимы между собой. На вершине их обычно имеется апикальный аппарат — специальное приспособление для вскрытия и освобождения аскоспор. У битуникатных сумок оболочка более толстая, явно двухслойная, обычно на вершине сильно утолщённая и без апикального аппарата. Наружный слой жёсткий, а внутренний эластичный. При созревании сумки под действием увеличивающегося тургорного давления наружный слой лопается, внутренний растягивается, и аскоспоры с силой выбрасываются.
Имеется также специальный лекапоровый тип сумок, встречающийся только у многих грибов, входящих в состав лишайников. Здесь, как и у битуникатных сумок, хорошо выражены два слоя, но они не отделимы друг от друга. Внутренний слой, более толстый, особенно утолщён на вершине. Здесь в нём может иметься особая внутренняя структура — осевое тельце. Оно отличается менее интенсивной амилоидной реакцией с йодом. При вскрытии сумки внешний слой лопается, а внутренний слой и осевое тельце растягиваются и выдаются наружу в виде клюва.
Аскоспор чаще всего бывает восемь, однако число их может быть и меньше (например, у многих мучнисторосяных) или, наоборот, много больше (например, у многих сордариевых). При этом большое число аскоспор может быть связано как с увеличением числа митозов после мейоза в сумке, так и с тем, что уже сформировавшиеся споры распадаются на фрагменты. Размер и вид аскоспор могут быть самыми различными. Обычно величина их составляет несколько микрометров, однако самые крупные аскоспоры (у некоторых микобионтов лишайников) могут быть до 525 мкм в длину. Форма аскоспор бывает от шаровидной или овальной до удлинённой и нитевидной. Как и конидии, они могут быть бесцветными (гиалиновыми) или окрашенными, состоять из одной, двух или большего числа клеток. Аскоспоры могут иметь различного рода придатки. У многих видов споры одеты слизистым чехлом.

Тип Аскомицеты (Ascomycota) – один из самых крупных объединений грибов. Включает около 30000 видов. Для аскомицетов или сумчатых грибов характерен многоклеточный (септированный) мицелий. В состав клеточной стенки входят полисахариды, хитин, бета-глюкокан. Основной признак грибов типа – формирование в результате полового процесса асков (сумок) с аскоспорами (сумкоспорами) [2] .
Аскомицеты (Ascomycota) и Базидиомикота (Basidiomycota) на основании формирования при половом размножении дикариона (ассоциации двух ядер (n+n)) объединяют в общую группу Дикариомицеты. В некоторых системах его трактуют как отдел Dikaryomycota с двумя подотделами Ascomycota и Basidiomycota, в других – как надотдел (надтип) Dikaryomycoteraс двумя отделами (типами) – Ascomycota и Basidiomycota [1] .

Аски на листьях

Систематика Аскомицетов
Тип Аскомикота традиционно подразделяют на четыре класса:
1. Класс Тафриномицеты (Taphrinomycetes) (= Археаскомицеты (Archaeascomycomycetes)) – наиболее древняя и примитивная группа, исходная для остальных аскомицетов. Группа разнородна по морфологии, включает мицелиальные и дрожжевидные формы, встречаются виды с плодовыми телами [1] .
2. Класс Гемиаскомицеты, Голосумчатые (Hemiascomycetes)(= Схизосахаромицеты, делящиеся дрожжи (Schizosacchromycetes) – плодовые тела отсутствуют, аски протоуникатьные, образуются непосредственно на мицелии либо при слиянии одиночных клеток [2] [1] .
3. Класс Эуаскомицеты, настоящие сумчатые или плодосумчатые (Euascomycetes)(= Ascomycetes) – аски эутуникатьные – унитуникатьные, реже протоуникатные, образуются внутри или на поверхности плодовых тел [2] .
4. Класс Локулоаскомицеты (Loculoascomycetes) – аски эутуникатные – битуникатные, образуются в локулах (особых полостях), формирующихся в сплетении мицелия, так называемой аскостроме [2] .
Тип Аскомицеты (Ascomycota) включает около 75% всех описанных видов грибов. В настоящее время существует тенденция перенесения в тип Аскомицеты видов из типа Anamorphic fungi, характеризующихся сходным строением вегетативных структур и клеточной стенки при отсутствии в цикле развития асков и наличии только анаморфной стадии [2] .
Кроме того к типу Аскомицеты относят лишайники, грибные компоненты которых почти исключительно аскомицеты [1] .

Кклейстотеции

Особенности номенклатуры
Четко отмечено, в отдельных таксонах различных рангов наблюдается тенденция либо к усовершенствованию в ходе эволюции половых спороношений при подавлении или отсутствии бесполого размножения, либо к усовершенствованию бесполого размножения при подавлении полового размножения. Во втором варианте гриб представлен только вегетативными структурами (чаще всего мицелием) и бесполыми спороношениями, которые затрудняют точное определение его таксономического положения. Это создает затруднения для систематики и диагностики [1] .
Точное систематическое положение видов, известных только в анаморфной стадии, выяснить часто невозможно. Одновременно не представляется возможным во многих случаях установить родственные связи между бесполым и половым спороношением. Эта ситуация привела к возникновению особых правил номенклатуры, то есть присвоения и использования видовых названий [1] .
Виды, имеющие два и более типов спороношения, в частности, представители рода Claviceps, называют плеоморфными. Бесполые спороношения и другие бесполые структуры, например, склероции – анаморфы, половые спороношения – телеоморфы, а всю совокупность стадий развития – голоморфа [1] .
Каждая стадия может иметь свое собственное название. Название как телеоморфы, так анаморфы в настоящее может распространяться на все стадии (голоморфу). Для некоторых видов название анаморфы не распространяется на телеоморфу и голоморфу. В частности, аскомицет Eurotium herbariorum имеет конидиальную стадию – Aspergillus glaucus. В данном случае можно говорить о конидиеносцах и конидиях Eurotium herbariorum, но нельзя называть плодовые тела и аски – Aspergillus glaucus [2] [1] .

Плодово тело

гриб Morchella vulgaris (Сморчок обыкновенный) [5]
Общая характеристика
Виды грибов, отнесенные к типу Аскомицеты (Ascomycota) характеризуются следующими общими признаками:
- Вегетативное тело – чаще всего разветвленный септированный, гаплоидный мицелий, Перегородки обычно не замкнутые, через пору в них мигрируют разнообразные клеточные структуры, в том числе ядра. Клеточные стенки аскомицетов, как правило, двухслойные. Клетки обычно многоядерные, реже, как у видов порядка Эризифовые (Erysiphales) – одноядерные. Нередко встречаются дрожжевые формы. Для целого ряда представителей известен мицелиально-дрожжевой диморфизм [1] .
- В состав клеточной стенки Аскомикот входят полисахариды – хитин и глюкан. Содержание хитина в клеточной стенке составляет не более 20–25 % полисахаридов [1] .
- Вегетативное размножение – фрагментами мицелия, путем деления дрожжевых клеток. Широко распространено бесполое размножение. Часто оно преобладает или служит единственным видом воспроизведения. Вегетативное размножение осуществляется путем образования конидий на конидиеносцах[1] .
- Половое размножение обычно – гаметангиогамия, партеногамия, дейтерогамия, редко – сперматизация, соматогамия. У подавляющего большинства видов, при половом размножении, ядра образуют дикарион[1][3] .
- Половой процесс и образование асков с аскоспорами часто сопряжен с развитием плодовых тел (клейстотеции, перитеции, апотеции, псевдотеции) [1] .
- В жизненном цикле аскомицетов преобладает гаплоидная стадия. Дикариотическая стадия, представленная аскогенными гифами, занимает незначительную часть цикла развития[1] .
- Тип Аскомицеты (Ascomycota) включает гомоталличные виды и гетероталличные виды. Гетероталлизм биполярный [1] .
Распространение в природе
Тип Аскомицеты (Ascomycota) – широко распространен в природе. Среди них можно найти представителей практически всех эколого-трофических групп грибов, как сапротрофов, так и паразитов растений, животных, человека [4] .
Практическое значение
Тип Аскомицеты (Ascomycota) – включает множество паразитических грибов, в том числе возбудителей экономически значимых заболеваний культурных растений. К таким относят виды, входящие в семейство Эризифовые (Erysiphaceae), вызывающие такие заболевания как роса мучнистая гороха, роса мучнистая моркови, роса мучнистая томатов [4] .
Многие аскомицеты являются активными агентами биодеструкции – разрушения природных материалов и изделий из них. От некоторых аскомицетов страдают пищевые продукты, в частности Eurotium herbariorum – возбудитель плесневения семян [4] .
Аскомицеты широко используются человеком в кулинарии (дрожжи), как продуценты антибиотиков, в пищу (трюфели, сморчки). Виды, паразитирующие на насекомых и других грибах, служат основой для производства биологических пестицидов [1] .
Читайте также: