
Беклемишев в н экология малярийного комара
Обновлено: 24.04.2024
Экология личинки малярийного комара [Текст] : [Сборник статей] : 2 рис. в тексте / Под ред. Е. И. Марциновского, П. Г. Сергиева, В. Н. Беклемишева. - Москва ; Ленинград : Биомедгиз, 1934 ([М.] : 16 тип. треста "Полиграфкнига"). - Обл., 167 с. : граф.; 23х15 см. - (Труды Тропического института/ Лаборатория гидробиологии).
(Труды Тропического института/ Лаборатория гидробиологии)
Некоторые основные моменты эпидемиологические значения в экологии личиночной стадии Anopheles maculipennis
В. И. Олифан
О количественном методе, применяемом нами в полевых исследованиях экологии личинки Anopheles maculipennis
В. И. Олифан
Экспериментальные наблюдения над влиянием температуры на развитие личинок Anopheles maculipennis
В. И. Олифан
Развитие и выживаемость личинок Anopheles maculipennis при разном pH
В. И. Олифан
Экология личинок Anopheles maculipennis в водоемах торфоразработок Московсковской области
В. И. Олифан
В. Д. Успенская
П. И. Рахманова
Н. Я. Маркович
Планктон водоемов торфоразработок и личинки Anoph. maculipennis
П. И. Рахманова
Наблюдения над экологией личинок Anopheles maculipennis в водоемах г. Рязани
М. Л. Мельникова
В. И. Олифан
К вопросу о влиянии биологических и химических факторов на развитие личинок анофелеса
Лонгинов
Охотникова
Лебедева
Платонова
Экспериментальные исследования влияния плотности популяции Anopheles maculipennis на численность их потомства
В. И. Олифан
К вопросу о причинах большой заселенности личинками Anopheles maculipennis водоемов, заросших водными растениями
В. И. Олифан
Влияние света на развитие личинок Anopheles maculipennis
В. Олифан
П. Рахманова
О влиянии рода пищи на личинки Anopheles maculipennis и Anopheles bifurcatus
В. И. Олифан
М. С. Владимирова
и П. И. Рахманова
Испытание на водоемах применимости искусственного понижения pH как метода борьбы с личинками малярийного комара
В. И. Олифан
Г. Е. Раевский
О. Н. Виноградская
Результаты гидробиол. обследования прудов-холодильников
Г. Т. Винберг
О. Н. Виноградская
М. С. Владимирова
Г. И. Павлова
Шифр хранения:
FB Я 199/14
FB T 41/89
Электронный заказ
ГОНОТРОФИЧЕСКИЙ ЦИКЛ (греч, gonos рождение, семя + trophe питание) — совокупность взаимосвязанных процессов питания и размножения у кровососущих насекомых отряда двукрылых. Самки кровососущих двукрылых могут откладывать несколько кладок; каждой яйцекладке предшествует акт кровососания. Т. о., жизнь самки состоит из ряда следующих друг за другом Г. ц., которые и обусловливают роль кровососущих насекомых как переносчиков инфекций. В природе кровососание может проходить на разных, в т. ч. и на зараженных, животных; нападая затем на человека и осуществляя паразитарный контакт его с дикой природой, кровососущие насекомые могут стать переносчиками возбудителей болезней (см. Переносчики, Природная очаговость, природноочаговые болезни, Трансмиссивные болезни). Периодичность питания кровью и параллелизм процессов пищеварения и развития яичников, обозначаемый как гонотрофическая гармония, свойственны всем семействам кровососущих низших двукрылых: комарам (см. Комары кровососущие), мошкам (см.), мокрецам (см.), слепням (см.), москитам (см.).
Наличие связи между этими процессами впервые отметил Г. А. Кожевников в 1903 г. В случае гонотрофической гармонии прием одной полной порции крови необходим и достаточен для созревания одной порции яиц. Каждый Г. ц., по В. Н. Беклемишеву (1940, 1944), состоит из трех фаз: поиска добычи, нападения и акта кровососания; переваривания крови и созревания яиц; поиска водоема и откладки яиц. Селла (М. Sella, 1920) разделил 1-ю и 2-ю фазы цикла у комаров рода Анофелес на 7 стадий, отличающихся по степени переваривания крови и развитию яичников. Стадии различимы по внешнему виду брюшка. Число Г. ц., проделанных самкой, является показателем ее физиол. возраста. Зная продолжительность каждого Г. ц. и физиол, возраст самки, можно рассчитать календарный возраст переносчика, т. е. число прожитых им дней. Методы определения продолжительности всех трех фаз цикла и методы определения физиол, возраста самок переносчиков описаны Т. С. Детиновой (1962). Изучение гонотрофических взаимоотношений и физиол, возраста самок позволяет определить возрастной состав популяций вида, эпидемиол, значимость переносчиков, оценить эффективность проводимых противоимагинальных мероприятий и дифференцировать методы борьбы для отдельных групп кровососов.
Библиография: Беклемишев В. Н. Экология малярийного комара, М., 1944; Д е-тиноваТ. С. Методы установления возрастного состава двукрылых насекомых, имеющих Медицинское значение, М., 1962, библиогр.; П о л о в о д о в В. П. Определение физиологического возраста самки Anopheles, то есть числа проделанных ею тонотрофических циклов, Мед. паразитол., в. 4, с. 352, 1949; Руководство по медицинской энтомологии, под ред. В. П. Дербе-невой-Уховой, М., 1974, библиогр.; Мап-son’s tropical diseases, ed. by Ch. Wilcocks -a. Ph. E. G. Manson-Bahr, Baltimore, 1972.
(22.09(04.10) 1890, Гродно – 04.09.1962, Москва)
зоолог, действительный член АМН СССР, лауреат Сталинской премии, профессор Московского университета, почетный член МОИП.
В.Н.Беклемишев родился в г.Гродно в семье врача. В 1913 г. окончил Императорский Санкт-Петербургский университет, в котором специализировался по зоологии беспозвоночных, под руководством профессоров В.Т.Шевякова, В.А.Догеля, В.М.Шимкевича. После окончания университета был оставлен для подготовки к профессорскому званию. В 1918 г. переезжает в Пермь, где занимает должность доцента в недавно основанном Пермском университете (в 1920 г. становится профессором того же университета). С 1932 г. заведующий отделом энтомологии Института малярии и медицинской паразитологии в Москве (ныне Институт медицинской паразитологии и тропической медицины им. Е.И.Марциновского). С 1934 г. профессор кафедры зоологии и сравнительной анатомии Московского государственного университета, где читал курс сравнительной анатомии беспозвоночных.
Академик АМН СССР В.Н. Беклемишев выполнил классические исследования биологии малярийного комара и других кровососущих членистоногих. Под его руководством была разработана система мер, в результате которых в СССР была практически полностью ликвидирована малярия. Он создал ряд оригинальных концепций в биоценологии и общей паразитологии.
В.Н.Беклемишев – крупный специалист по ресничным червям, основоположник школы паразитологов и медицинских энтомологов, создал учение о малярийных ландшафтах, явившееся основой прогнозирования заболеваемости малярией и разработки мероприятий по ее ликвидации. Основные труды по теоретическим основам сравнительной анатомии беспозвоночных, экологии, биоценологии, медицинской энтомологии, сравнительной и эволюционной паразитологии. Государственная премия СССР (1944, 1952).
В.Н.Беклемишев – действительный член АМН СССР (1945) и Польской академии наук (1949), заслуженный деятель науки РСФСР (1947), лауреат Сталинских (1946, 1951) и Государственных премий СССР (1944, 1952), член Московского общества испытателей природы. За научные заслуги избран почетным членом МОИП.
Библиография:
Мазурмович Б.Н. Выдающиеся отечественные зоологи. – М.: Учпедгиз, 1960. – 428 с.
Вопросы общей зоологии и медицинской паразитологии : к 70-летию со дня рождения В.Н. Беклемишева : Сб. ст. / под ред. П.Г.Сергиева. – М.: Медгиз, 1962. – 611 с.
Светлов П.Г. Памяти Владимира Николаевича Беклемишева (1890–1962) // Беклемишев В.Н. Методология систематики. М. : KMK Scientific Press, Ltd., 1994. C. 6-16.

3. Перевозкин В.П., Бондарчук С.С., Гордеев М.И. Популяционно-видовая структура малярийных комаров (Diptera, Culicidae) Прикаспийской низменности и Кумо-Манычской впадины // Медицинская паразитология и паразитарные болезни. – 2012. – № 1. – С. 12-17.
6. Стегний В. Н. Популяционная генетика и эволюция малярийных комаров. – Томск: Изд-во Томск. ун-та, 1991. – 136 с.
7. Стегний В.Н., Кабанова В.М. Хромосомный анализ малярийных комаров Anophelesatroparvus и A. maculipennis (Diptera, Culicidae) // Зоол. журн. – 1978. – Т. 57, № 4. –С. 613 619.
8. CatorLauren J., Arthur Ben J., Harrington Laura C. and HoyRonald R. Harmonic Convergence in the Love Songs of the Dengue Vector Mosquito. Science, 2009. V. 323. No. 5917. P. 1077-1079.
9. Stegniy V.N., Kabanova V.M., Novikov Y.M., Pleshkova G.N. Inversion polymorphism of malaria mosquito Anopheles messeae. I. Distribution of inversions in area of species // Rus. J. Genetics. 1976. V.12. № 4. P. 47-55.
10. Warren B., Gibson G. and Russell I. J.Sex Recognition through Midflight Mating Duets in CulexMosquitoes Is Mediated by Acoustic Distortion. Current Biology, 2009. Vol. 19. No. 6. Р. 485-491.
Акустические особенности насекомых имеют важнейшее значение при размножении, благодаря чему обеспечивается встреча особей противоположного пола и репродуктивная изоляция близких симпатрических видов [1]. Неослабевающий интерес к звуковой коммуникации комаров объясняется как желанием понять наиболее сложные формы их жизнедеятельности, так и необходимостью управлять поведением и контролировать численность переносчиков опасных трансмиссивных заболеваний.
Малярийные комары рода Anopheles (Diptera, Culicidae) являются объектами постоянного повышенного внимания со стороны биологов и медицинских служб ввиду эпидемиологического значения этих насекомых. В то же время представители данной группы являются прекрасными модельными объектами для генетического и экологического анализа [2, 6].
Материал и методы
Материалом для данной работы послужили комары четырех видов-двойников: An. messeaeFalleroni, An. beklemisheviStegniyandKabanova, An. maculipennisMeigen и An. atroparvusvanThiel. Имаго An. atroparvus использованы из лабораторной линии; имаго остальных видов выведены из потомств от гоноактивных самок, выборки которых были взяты в природных популяциях – An. messeae иAn. beklemishevi в Томской области, An. maculipennis в Белоруссии. Среди использованных потомств An. beklemishevi, An. atroparvus и An. maculipennis оказались цитогенетически мономорфными, а An. messeae – инверсионно полиморфным. Поэтому в последнем случае цитогенетически определяли кариотипы особей из каждого потомства. У выращенных имаго всех четырех видов снимались акустические характеристики. Для этого комаров приклеивали дорсальной стороной груди к препаровальной игле (рис. 1) и помещали в звуковую камеру с микрофоном (рис. 2). Для регистрации акустических сигналов использовался компьютер и соответствующее программное обеспечение.

Рисунок 1.Фиксация комара перед микрофоном

Рисунок 2.Звуковая камера с микрофоном
Результаты и обсуждение
В различных регионах Палеарктики установлена симпатрия представителей малярийных комаров в разнообразных сочетаниях [3, 6]. В связи с этим в процессе эволюции у комаров должны были сформироваться четкие биологические докопуляционные механизмы, обеспечивающие встречу и распознавание особей противоположного пола своего вида для скрещивания. Учитывая то, что скрещивание комаров происходит в полете и не редко в темное время суток [2], первостепенное значение в видовой идентификации, очевидно, имеют акустические сигналы, издаваемые машущими крыльями.
Межвидовой сравнительный анализ акустики трех представителей Anopheles с учетом полов показал неоднозначные различия в значениях их частот.
У самок An. atroparvus в звуковом спектре выявлены три пика (рис. 3, Female): первый, самой высокой амплитуды – основная частота (среднее значение для выборки f = 293 Гц, число особей n = 72); второй и третий пики – гармоники, или обертоны, кратные значению главного пика (584 и 880 Гц, соответственно).
У самцов An. atroparvus два хорошо выраженных пика (третий слабо отличается от фоновых колебаний) (рис. 3, Male): первый (основная частота), среднее значение для выборки f = 444 Гц (n = 29); второй пик – гармоника – средняя частота 887 Гц. Необходимо подчеркнуть, что анализируемые частоты особей одного пола An. atroparvus статистически совпадают на уровне значимости α = 0,05. Таким образом, средняя основная частота самца выше, чем у самки, на 151 Гц, соответственно различаются и гармоники.
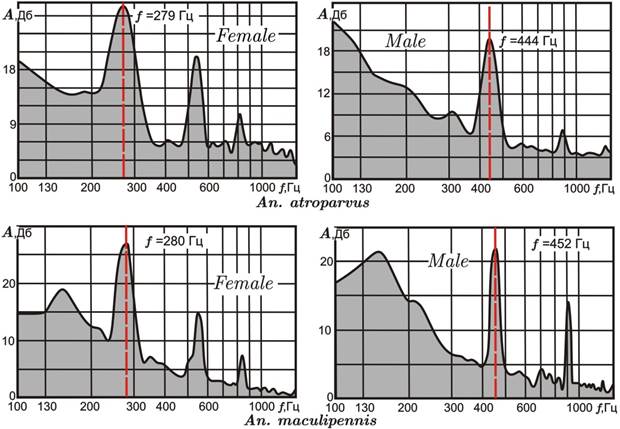
Рисунок 3. Типовые спектры звуковых частот самок (Female) и самцов (Male) An. atroparvusи An. maculipennis
Красными пунктирными линиями на диаграммах отмечены пики основных частот
У самок An. maculipennis в звуковом спектре выявлены также три пика (рис. 3, Female): первый (основная частота) – среднее значение для выборки f = 272 Гц; второй и третий (гармоники) – 548 и 820 Гц, соответственно (n = 11). У самцов An. maculipennis так же, как и у An. atroparvus, в звуковом спектре хорошо выражены два пика (рис. 3, Male): первый (основная частота) – среднее значение для выборки f = 448 Гц (n = 15); второй пик (гармоника) – средняя частота 890 Гц. Средняя основная частота самца выше, чем у самки, на 176 Гц, соответственно различаются и гармоники.
Таким образом, An. atroparvus и An. maculipennis имеют близкие акустические характеристики (у самцов они практически совпадают), чем в частности можно объяснять феномен межвидовой гибридизации этих двух видов. Единичные межвидовые гибриды на стадии личинок, отловленные ранее в природных биотопах Молдавии (г. Бельцы) и Калмыкии (г. Элиста), были выявлены посредством цитогенетического анализа [3, 7].
Лабораторная гибридизация An. atroparvus × An. maculipennis показала полную стерильность самцов F1 и некоторую фертильность самок F1. Последние, будучи способными к скрещиванию с самцами An. atroparvus, совершенно не развивали яиц. Очевидно, репродуктивная изоляция между An. atroparvus и An. maculipennis, четко проявляющаяся на уровне стерильности гибридов, не подкрепляется поведенческой изоляцией. Можно сделать вывод, что абсолютных этологических барьеров при скрещивании между этими видами не существует, что хорошо соотносится с полученными характеристиками их звуковых частот.
В-третьих, диапазон главных частот, прежде всего у самцов An. messeae, оказался гораздо разнообразнее, чем у других исследованных видов, что, очевидно, связанно с внутривидовым инверсионным полиморфизмом этого представителя. Перестройки затрагивают как половую хромосому (XL), так и плечи двух аутосом (2R, 3R, 3L) [9].
В юго-западных регионах Палеарктики преимущественно встречаются эволюционно исходные варианты – XL0 (XL1), 2R0, 3R0, 3L0; в северо-восточных – XL1, XL2, 2R1, 3R1, 3L1. В центре ареала An. messeae популяции характеризуются смешенным составом инверсий, хотя их определенные сочетания в кариотипах регистрируются значимо чаще, чем это ожидалось исходя из частот отдельных перестроек [4, 6]. В соответствие с этим было сделано предположение, что в полиморфных гемипопуляциях имаго An. messeae имеет место ассортативное скрещивание (предпочтительное спаривание генетически сходных особей).
Статистический анализ по критерию Стьюдента не выявил значимых различий по всем частотным пикам у самок с различными кариотипами (основная частота от 211 до 225 Гц).
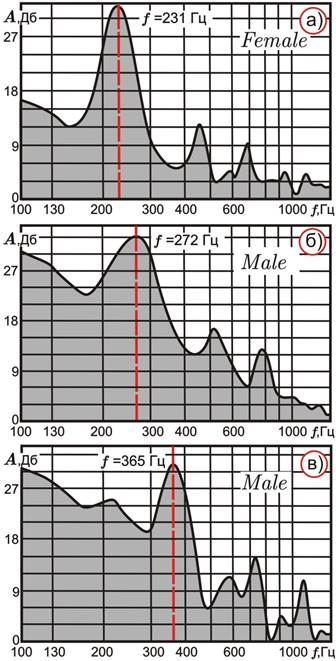
Рисунок 4. Типовые акустические спектры самки и самцов An. messeae
Известно, что самцы Anopheles в вечернее время собираются в рои, чтобы усилить звучание для привлечения самок [2]. Остается открытым вопрос, каким образом происходит окончательная идентификация самкой полового партнера, так как кариотипическая структура роев до сих пор не изучалась.
Возможны два варианта: либо рой преимущественно состоит из самцов с определенными кариотипами, и самка удаленно его идентифицирует; либо рой представляет собой полиморфную гемипопуляцию самцов, и тогда выбор самкой полового партнера осуществляется уже внутри роя.
Таким образом, полученные результаты акустических характеристик близкородственных видов малярийных комаров позволяют рассматривать их звуковую коммуникацию как основной механизм для видовой и внутривидовой идентификации полового партнера при спаривании.
Работа выполнена при поддержке Минобрнауки России в рамках базовой части государственного задания № 2014/387/1029.
Рецензенты:
Сибатаев А.К., д.б.н., старший научный сотрудник лаборатории эволюционной цитогенетики НИИ биологии и биофизики при ТГУ (НИИББ), г. Томск.
Читайте также: