
Центри кори головного мозку людини
Обновлено: 19.04.2024
Значение, роль коры больших полушарий головного мозга человека
В статье мы рассмотрим локализацию функций, участки, анализаторы, поля, участки, области зоны коры больших полушарий головного мозга человека (мужчины, женщины). Неврологи, невропатологи, рефлексотерапевты, рефлексологи выделяют 4 основных положения, применительно к практической деятельности невропатолога, современного учения о локализации функций в коре головного мозга.
1. Очень сложная морфологическая и функциональная дифференциация коры больших полушарий головного мозга. Лобная доля больше отвечает за двигательные функции. Теменная, затылочная и височная зоны больше отвечают за чувствительные функции.
2. Динамичность и относительность локализаций функций коры головного мозга. Определенный участок коры головного мозга, обеспечивая какую-то одну функцию, в то же время в разнообразных сочетаниях с другими ее полями может участвовать в осуществлении различных корковых функций и образовывать новые кортикальные связи. Это имеет значение в процессах компенсации при таких состояниях, как поражение коры головного мозга, нарушение коры головного мозга, смерть или повреждение коры головного мозга, отмирание, незрелость коры головного мозга.
3. Формирование специальных корковых областей в процессе практической деятельности.
Функция творит центр
Локализация функций и симптомов
Проводя топическую диагностику рефлексотерапевт, невролог, невропатолог, микроневропатолог, детский невролог, взрослый невролог определяет не только локализацию поражения корковых центров, но и локализацию симптомов. Простые корковые функции связаны с проекционными пластинками коры (пятой и четвертой), имеющими непосредственную связь с периферией и являющимися корковыми отделами анализаторов. Сложные корковые функции связаны с ассоциативными слоями коры (вторым и третьим). Последние слои соединены горизонтальными волокнами с другими участками коры головного мозга в пределах одного полушария и не имеют прямого выхода на периферию. Большое значение в обеспечении сложных корковых функций имеют также комиссуральные связи между полушариями, проходящими через мозолистое тело.
Простые корковые функции обычно представлены в обоих полушариях головного мозга. Сложные корковые функции чаще имеют асимметричное представительство в правом или левом полушарии головного мозга. Итак, какие бывают поля, участки, области, типы коры головного мозга, отделы, анализаторы, части коры головного мозга?
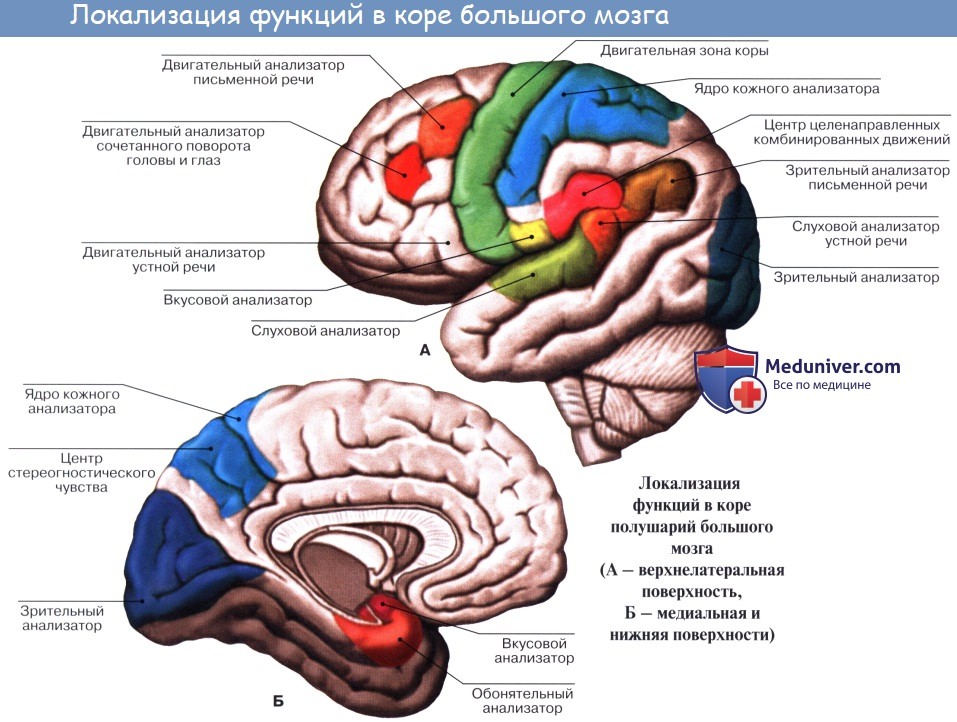
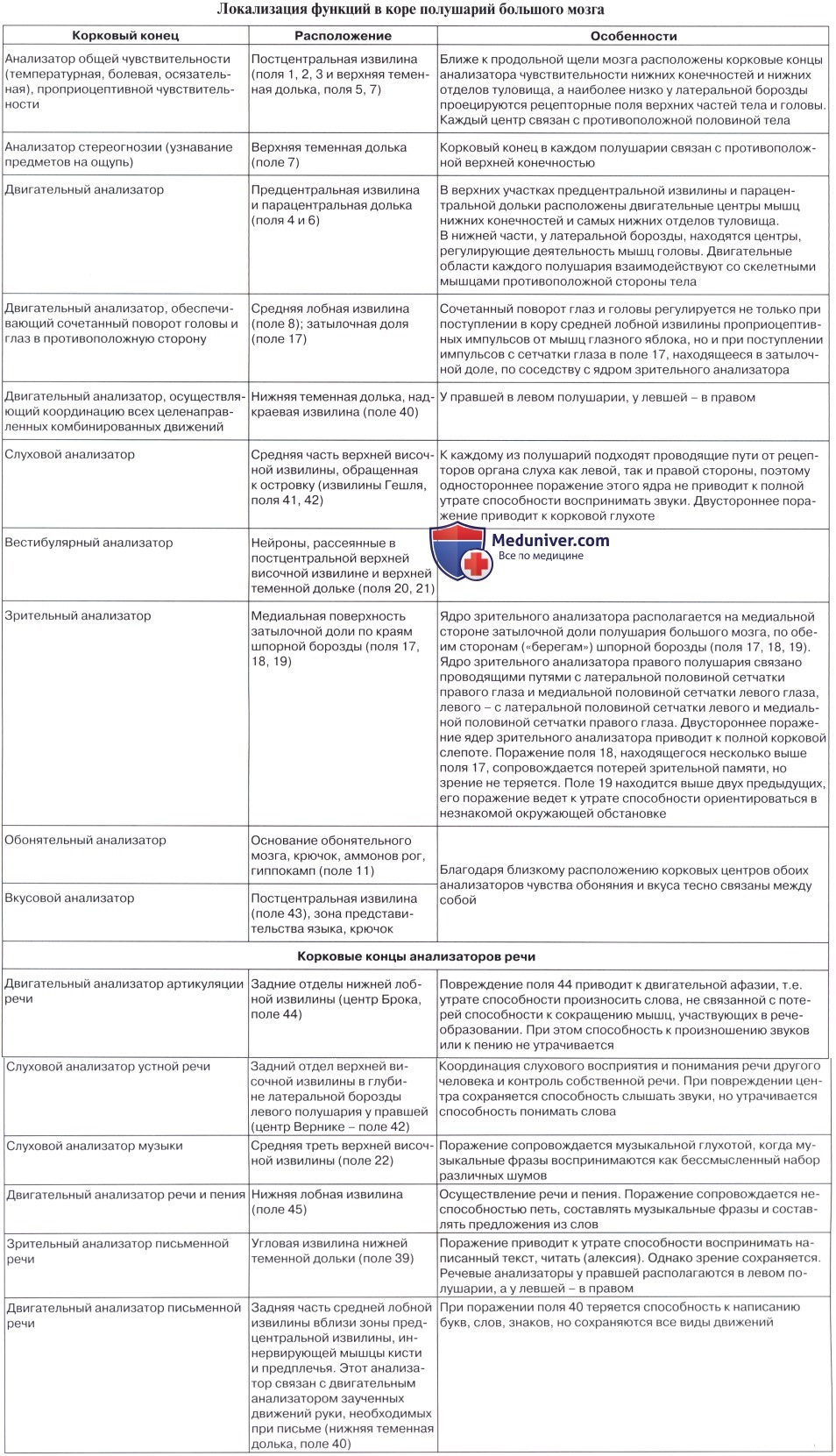
Двигательная кора головного мозга, двигательные центры головного мозга, двигательные анализатор, моторный
Главным корковым отделом двигательного анализатора, его первичным полем, является предцентральная извилина, в верхних отделах которой находится проекционная область мышц стопы, голени, бедра, в средней части – туловища и руки, в нижней трети – лица. Двигательная иннервация построена по соматотопическому принципу. На этом уровне осуществляются тонкие дифференцированные движения. Кроме того, имеются дополнительные двигательные зоны – это вторичные поля двигательного анализатора и третичные поля двигательного анализатора. Дополнительные двигательные зоны обеспечивают сложные автоматизированные двигательные акты. Например, в парацентральной дольке находятся корковые центры тазовых органов. В задних отделах верхней лобной извилины находится переднее адверсивное поле. Заднее адверсивное поле располагается на границе верхней теменной дольки и затылочной области. Задние отделы средней лобной извилины отвечают за сочетанный поворот головы и глаз в противоположную сторону. Задние отделы нижней лобной извилины осуществляет движения типа орального автоматизма – глотание, жевание, лизание.
Чувствительная кора головного мозга, чувствительные центры головного мозга, чувствительный анализатор
Главным корковым отделом поверхностных и глубоких видов чувствительности является постцентральная извилина, где также имеется соматотопическое представительство участков периферии, аналогичное вышеуказанному. К поверхностной чувствительности относятся температурная чувствительность, болевая чувствительность, тактильная чувствительность.
Стереогноз, стереогнозис
Сложные виды чувствительности локализованы в коре полушарий головного мозга на уровне верхней теменной дольки, где отсутствует соматотопика. К сложным видам чувствительности относятся стереогностическая чувствительность (стереогноз, стереогнозис), двумерно-пространственная чувствительность, чувство локализации и дискриминации. Зрительная проекционная зона (зрительная зона коры) занимает область шпорной борозды – внутренняя поверхность затылочной доли. Слуховая проекционная зона (слуховая зона коры) занимает центр верхней височной извилины и извилину Гешля. Вестибулярная проекционная зона находится рядом со слуховой. Обонятельная проекционная зона локализуется на внутренней поверхности височной доли, в извилине гиппокампа. Вкусовая проекционная зона находится рядом с последней, а также в области покрышки и островка Reili.
Теперь остановимся на локализации сложных корковых функций.
Обычно сложные корковые функции локализуются в левом полушарии головного мозга у правшей и в правом полушарии головного мозга у левшей.
Речевой анализатор, центр Вернике, центр Брока, функция речи - сенсорный центр
Функцию речи обеспечивает сенсорный центр (центр Вернике), который располагается в заднем отделе верхней височной извилины. При поражении центра Вернике наблюдается сенсорная афазия. Также функцию речи обеспечивает двигательный центр (центр Брока), который располагается в области задних отделов нижней лобной извилины. При поражении центра Брока наблюдается моторная афазия. При патологии на стыке височной и затылочной долей формируется амнестическая афазия и семантическая афазия. Речевые зоны коры головного мозга.
Лексический анализатор, центр лексии, функция чтения
Функции чтения обеспечивает лексический центр (центр лексии). Центр лексии располагается в угловой извилине.
Графический анализатор, центр графии, функция письма
Функции письма обеспечивает графический центр (центр графии). Центр графии располагается в заднем отделе средней лобной извилины.
Счетный анализатор, центр калькуляции, функция счета
Функции счета обеспечивает счетный центр (центр калькуляции). Центр калькуляции располагается на стыке теменно-затылочной области.
Праксис, праксический анализатор, центр праксиса
Гностический центр, центр гнозиса
В правом полушарии у правшей, в левом полушарии головного мозга у левшей представлены многие гностические функции. При поражении преимущественно правой теменной доли может возникать анозогнозия, аутопагнозия, конструктивная апраксия. С центром гнозиса также связаны музыкальный слух, ориентация в пространстве, центр смеха.
Память, мышление
Наиболее сложные корковые функции – это память и мышление. Эти функции не имеют четкой локализации.
Память, функция памяти
В реализации функции памяти участвуют различные участки. Лобные доли обеспечивают активную целенаправленную мнестическую деятельность. Задние гностические отделы коры связаны с частными формами памяти - зрительной, слуховой, тактильно-кинестической. Речевые зоны коры осуществляют процесс кодирования поступающей информации в словесные логико-грамматические системы и словесные системы. Медиобазальные отделы височной доли, в частности гиппокамп, переводят текущие впечатления в долговременную память. Ретикулярная формация обеспечивает оптимальный тонус коры, заряжая ее энергией.
Мышление, функция мышления
Функция мышления – это результат интегративной деятельности всего головного мозга, особенно лобных долей, которые участвуют в организации целенаправленной сознательной деятельности человека, мужчины, женщины. Происходят программирование, регуляция и контроль. При этом у правшей левое полушарие является основой преимущественно абстрактного словесного мышления, а правое полушарие связано главным образом с конкретным образным мышлением.
Развитие корковых функций начинается с первых месяцев жизни ребенка, достигает своего совершенства к 20 годам.
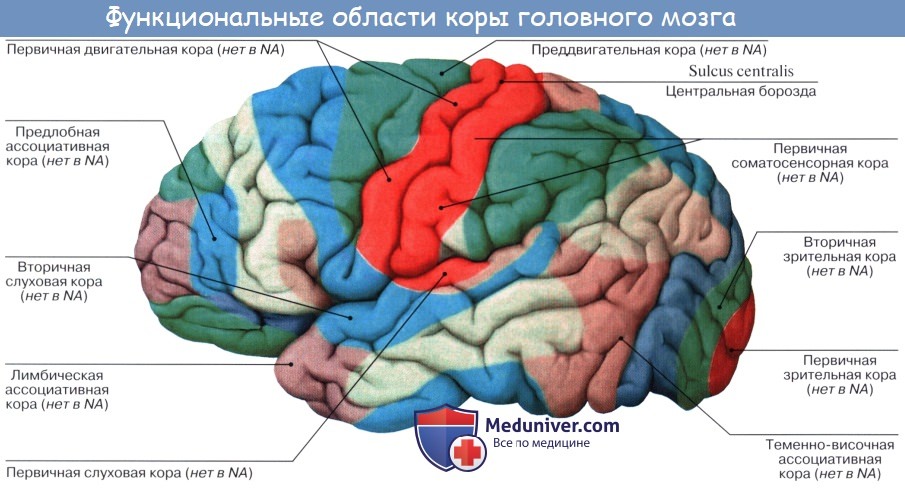
Зоны коры головного мозга
В последующих статьях мы остановимся на актуальных вопросах неврологии: зоны коры головного мозга, зоны больших полушарий, зрительная, зона коры, слуховая зона коры, моторные двигательные и чувствительные сенсорные зоны, ассоциативные, проекционные зоны, моторные и функциональные зоны, речевые зоны, первичные зоны коры головного мозга, ассоциативные, функциональные зоны, фронтальная кора, соматосенсорная зона, опухоль коры, отсутствие коры, локализация высших психических функций, проблема локализации, мозговая локализация, концепция динамической локализации функций, методы исследования, диагностики.
Кора головного мозга лечение
Строение коры головного мозга: слои, клетки
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
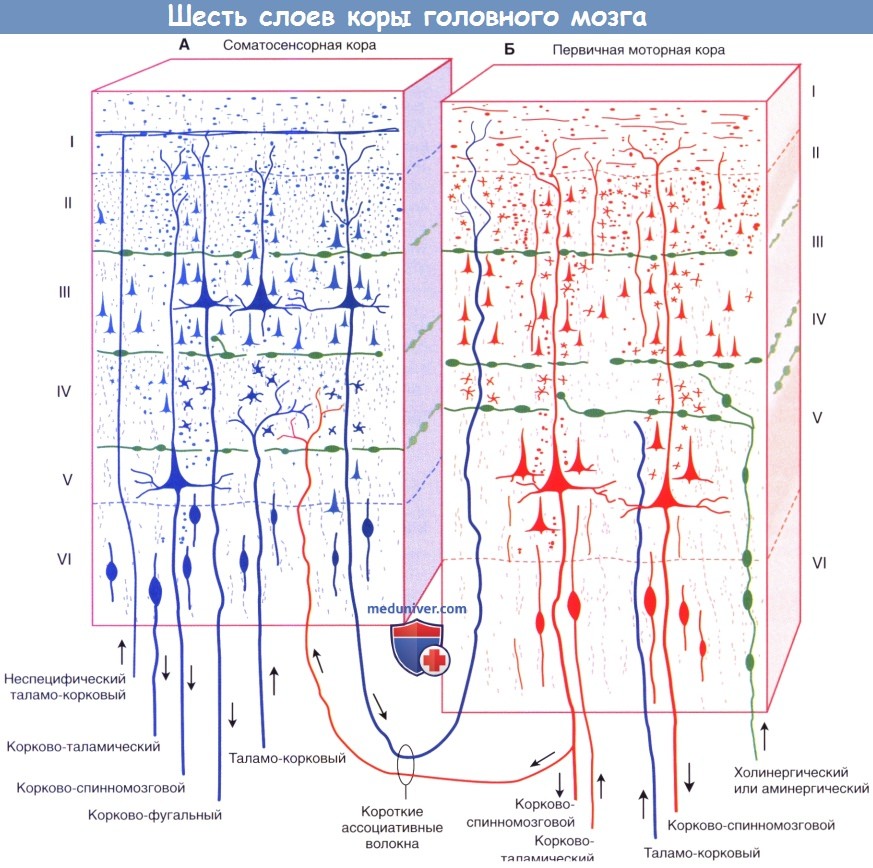
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
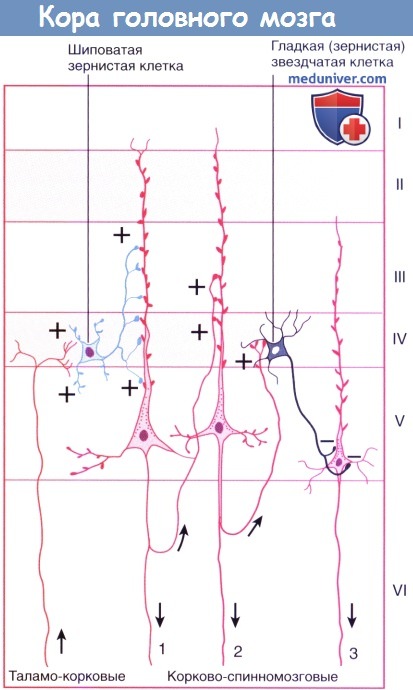
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
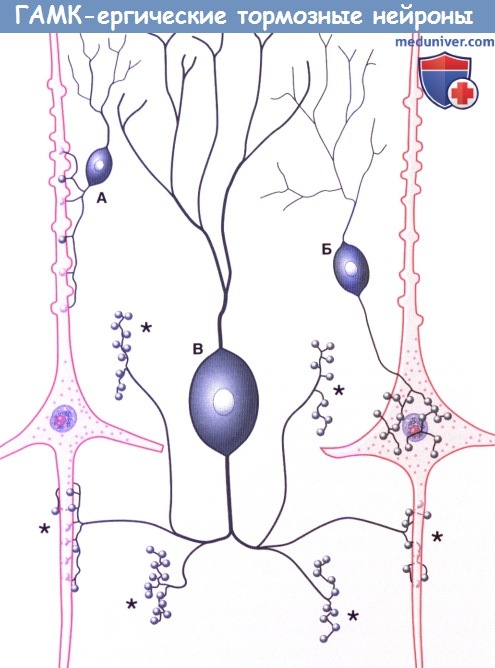
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Кора головного мозга. Физиологическая анатомия коры головного мозга
По иронии судьбы, из всех отделов головного мозга меньше всего мы знаем о функциях коры большого мозга, хотя совершенно очевидно, что это самая большая часть нервной системы. В тоже время известны эффекты поражения или специфической стимуляции различных участков коры. В первых статьях по физиологии на сайте обсуждаются фактические данные о функциях коры; затем кратко представлены основные теории о нервных механизмах, участвующих в мыслительных процессах, памяти, анализе сенсорной информации и др.
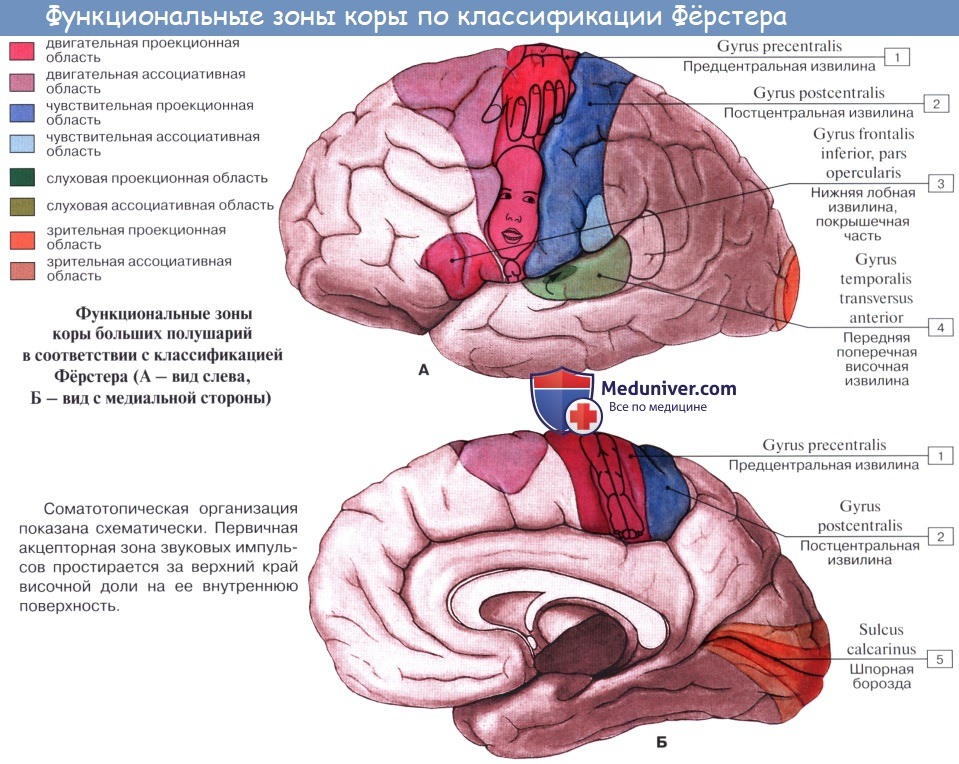
а) Физиологическая анатомия коры большого мозга. Функциональном частью коры большого мозга является гонкий слои нейронов, покрывающий поверхность всех извилин. Толщина этого слоя составляет лишь 2-5 мм, а общая площадь равна примерно 0,25 м. В целом кора большого мозга содержит примерно 100 млрд нейронов.
На рисунке ниже показана типичная гистологическая структура коры большого мозга с последовательными слоями разных типов нейронов.
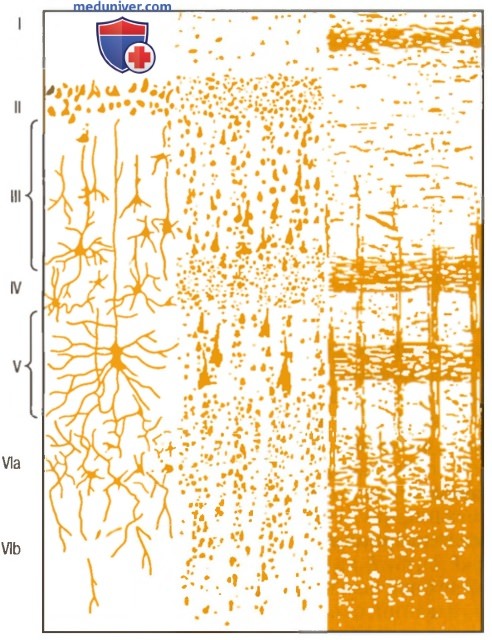
Структура коры большого мозга.
I - молекулярный слой; II — наружный зернистый слой;
III - слой пирамидных клеток; IV - внутренний зернистый слой;
V - слой больших пирамидных клеток. VI - слой веретенообразных клеток
Большинство нейронов относят к трем типам:
(1) зернистые (также называемые звездчатыми);
(3) пирамидные, получившие свое название за характерную пирамидальную форму.
Зернистые нейроны обычно имеют короткие аксоны и, следовательно, функционируют в основном как вставочные нейроны, которые проводят нервные сигналы только на небольшие расстояния в пределах самой коры. Некоторые нейроны — возбуждающие, они выделяют в основном нейромедиатор глутамат; другие — тормозящие, основным непромедиатором которых является гамма-аминомасляная кислота — главный тормозящий нейромедиатор.
Сенсорные области коры, а также ассоциативные, расположенные между сенсорными и моторными областями, содержат большое количество зернистых клеток, что дает основание предполагать высокую степень внутрикоргикалыюй обработки входящих сенсорных сигналов в пределах сенсорных и ассоциативных областеи.
Пирамидные и веретенообразные клетки дают начало почти всем волокнам, выходящим из коры. Пирамидные клетки — более крупные и мно гочисленпые, чем веретенообразные. Они являются источником длинных толстых нервных волокон, которые идут, не прерываясь, в спинной мозг. Они также дают начало крупным подкорковым пучкам ассоциативных волокон, проходящим от одной крупной части головного мозга к другой.
Справа на рисунке выше показана типичная организация нервных волокон в пределах разных слоев коры большого мозга. Обратите особое внимание на большое число горизонтальных волокон, которые распространяются между при лежащими областями коры. Также отметьте вертикальные волокна, идущие к коре и от нее к нижележащим областям мозга. Некоторые из них проходят весь путь до спинного мозга или направляются к отдаленным регионам коры большого мозга через длинные ассоциативные пучки.
Функции специфических слоев коры большого мозга обсуждаются в отдельных статьях на сайте (просим вас пользоваться формой поиска выше). Здесь следует вспомнить, что большинство входящих специфических сенсорных сигналов от тела поступают в IV слой коры. Большая часть сигналов покидают кору через нейроны, локализованные в V и в VI слоях; очень крупные волокна, идущие к мозговому стволу и спинному мозгу, берут начало главным образом в V слое, а громадное количество волокон, идущих к таламусу, возникают в VI слое.
Слои I, II и III осуществляют основную часть ассоциативных функций внутри коры, причем особенно большое число нейронов II и III слоев формируют короткие горизонтальные связи с прилежащими областями коры.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Морфологические основы динамической локализации функций в коре полушарий большого мозга (центры мозговой коры)
Знание локализации функций в коре головного мозга имеет огромное теоретическое значение, так как дает представление о нервной регуляции всех процессов организма и приспособлении его к окружающей среде. Оно имеет и большое практическое значение для диагностики мест поражения в полушариях головного мозга.
Представление о локализации функций в коре головного мозга связано прежде всего с понятием о корковом центре. Еще в 1874 г. киевский анатом В. А. Бец выступил с утверждением, что каждый участок коры отличается по строению от других участков мозга. Этим было положено начало учению о разнокачественности коры головного мозга — цитоархитектонике (цитос — клетка, архитектонес — строю). В настоящее время удалось выявить более 50 различных участков коры — корковых цитоархитектони-ческих полей, каждое из которых отличается от других по строению и расположению нервных элементов. Из этих полей, обозначаемых номерами, составлена специальная карта мозговой коры человека.
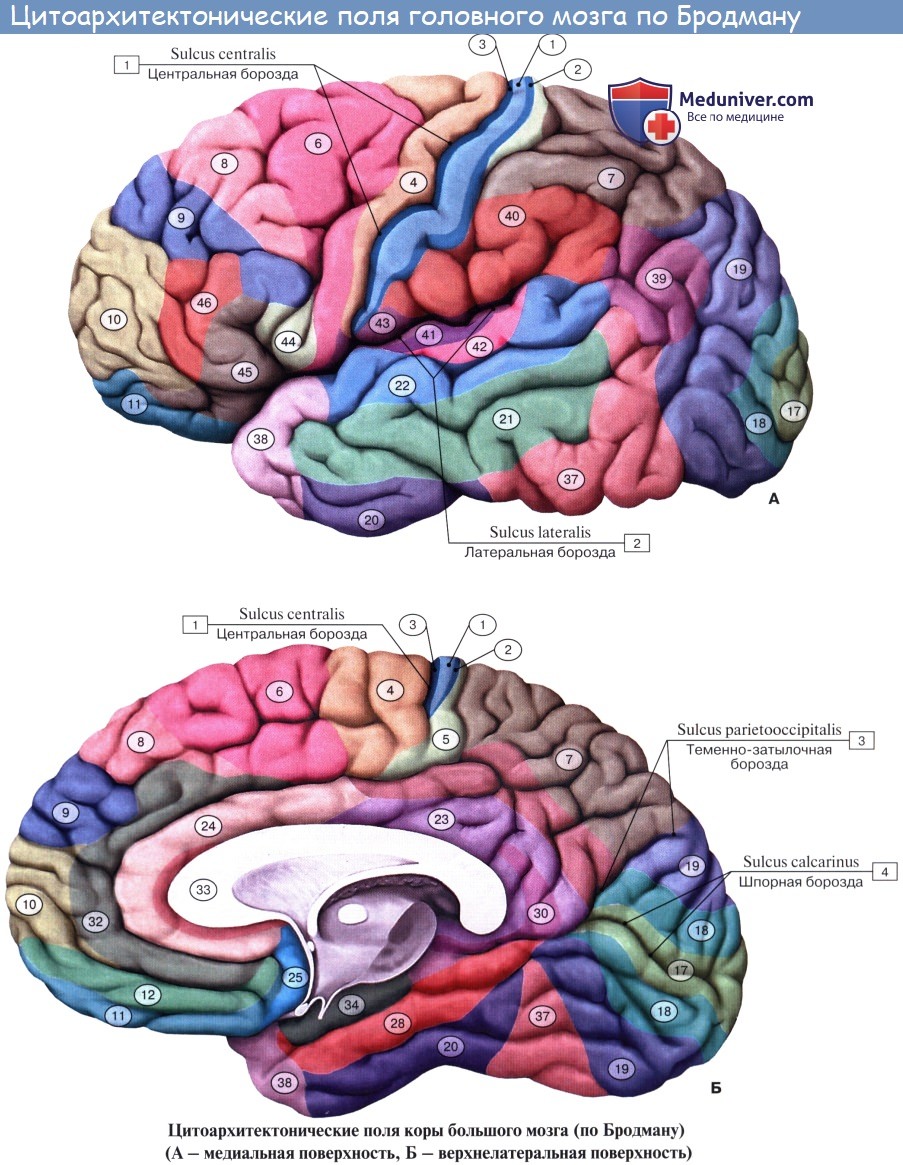
По И. П. Павлову, центр — это мозговой конец так называемого анализатора. Анализатор — это нервный механизм, функция которого состоит в том, чтобы разлагать известную сложность внешнего и внутреннего мира на отдельные элементы, т. е. производить анализ. Вместе с тем благодаря широким связям с другими анализаторами здесь происходит и синтез, сочетание анализаторов друг с другом и с разными деятельностями организма.
Редактор: Искандер Милевски. Дата последнего обновления публикации: 30.7.2020
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: