Фораминиферы паразиты или нет
Обновлено: 24.04.2024
Фораминиферы часто считаются самой важной группой морских микрофоссилий, поскольку они очень многочисленны в отложениях. Кроме того, они представляют большое разнообразие видов, которые очень полезны для биостратиграфических , палеоэкологических , палеоокеанографических исследований и т. Д. [ 2 ]
Описано около 10 000 нынешних видов фораминифер [ 3 ] [ 4 ] в дополнение к примерно 40 000 вымерших видов, известных по их окаменелостям. [ 5 ] Обычно они имеют размер менее 1 мм, но некоторые виды достигают до 20 см. [ 6 ]
Характеристики
Экология
Фораминиферы в основном морские, и большинство из них обитает на или в донных отложениях (т. Е. Они бентические ), тогда как некоторые плавают в толще воды на разной глубине (т. Е. Они планктонные ). Некоторые из них были обнаружены в пресноводных или солоноватых условиях, а некоторые виды были идентифицированы в почве с помощью молекулярного анализа рибосомной ДНК . [ 7 ] [ 8 ] Они встречаются на всех широтах и глубинах, в том числе в полярных регионах и глубинных желобах.
Типичный размер фораминифер составляет от 0,1 до 0,5 см, хотя есть виды с размерами от 100 мкм до почти 20 см. Фораминиферы - это гетеротрофные организмы , которые питаются небольшими организмами, такими как диатомовые водоросли или бактерии, которых они захватывают с помощью сети псевдопод. Самые крупные виды фораминифер способны ловить нематод и ракообразных .
У некоторых фораминифер есть одноклеточные водоросли различных линий, такие как эндосимбионты , такие как зеленые водоросли , красные водоросли , золотые водоросли , диатомеи и динофлагелляты . Более крупные виды могут содержать большое количество плотно упакованных водорослей, составляющих настоящие фермы по выращиванию водорослей. Некоторые фораминиферы являются клептопластами , то есть они сохраняют хлоропласты водорослей, которых они поглощают, так что эти органеллы продолжают фотосинтезировать внутри фораминифер. [ 9 ]
Схема клетки фораминифера: 1 - эндоплазма , 2 - эктоплазма , 3 камеры, 4 поры, 5 отверстий, 6 - пищеварительная вакуоль , 7 - ядро , 8 - митохондрии , 9 - псевдоподии ( granurreticulopodia ), 10 - гранулы, 11-апертура, 12-пищевая частица, 13- аппарат Гольджи , 14 - рибосомы .


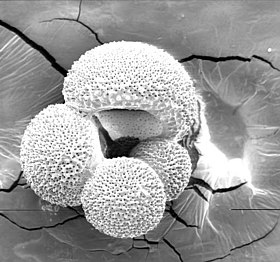
Гранурретикулоподия
Двумя отличительными чертами фораминифер являются раковина и сеть ложноножек. Клетка фораминифер делится на гранулярную эндоплазму, расположенную внутри оболочки, и на прозрачную эктоплазму, которая выходит из оболочки через основное отверстие или множество перфораций. Эктоплазма обычно полностью окружает раковину и отвечает за ее построение, поддерживаемую псевдоподиями. Между камерами оболочки через внутренние отверстия существует непрерывный поток цитоплазмы даже у видов со сложной оболочкой.
Псевдоноды образуются из эктоплазмы, ограничены клеточной мембраной и поддерживаются микротрубочками. Фораминиферы представляют механизмы для быстрой сборки и разборки микротрубочек, которые позволяют быстрое расширение и втягивание ложноножек. Они используются для передвижения, фиксации на субстрате, построения раковины и для захвата пищи. Длина ложноножек зависит от вида: у одних ложноножки в два или три раза превышают длину тела, у других - до 20 раз этой длины. [ 10 ] Движение фораминифер происходит путем прикрепления некоторых псевдопод к субстрату, выталкивая остальную часть клетки. Типичная скорость движения составляет от 1 до 2,5 см / час.
Псевдоподии называются гранурретикулоподиями ( гранулярные ретикулоподии ), поскольку они состоят из гранулярной цитоплазмы, длинные, тонкие и разветвленные, образующие сеть ретикулоподий путем укладки ( анастомоза ). Повсюду в псевдоподиях проходит двунаправленный цитоплазматический поток, несущий гранулы, которые могут состоять из частиц различных материалов, митохондрий, пищеварительных вакуолей или содержащих продукты жизнедеятельности, симбиотические динофлагелляты и т. Д. По этой причине одним из синонимов группы является гранулоретикулоза , хотя процесс транспортировки гранул еще полностью не изучен.
Скелет

Большинство видов образуют внутриэктоплазматический скелет, называемый панцирем, панцирем или панцирем. Это наиболее выдающаяся характеристика фораминифер и причина их относительной легкости к окаменению. Только примитивные фораминиферы являются моноталамусами (однокамерный скелет). Большинство фораминифер - политаламосы, то есть их раковина состоит из нескольких камер. Они разделены перегородками, которые имеют одно или несколько отверстий для соединения, называемых отверстиями, которые также дали название группе. Внутреннее пространство камер облицовано органической пленкой, по своей природе близкой к хитину . [ 11 ] [ 12 ] [ 13 ] Оболочка имеет одно или несколько основных отверстий, а также может иметь небольшие поры снаружи (перфорированные оболочки) или не иметь их (неперфорированные оболочки). Псевдоножки выходят через основные отверстия, а в случае перфорированных раковин также и через поры, если они достаточно широки.
Когда имеется более одной камеры, первое, что формируется, называется пролокулюсом и обычно является самым маленьким. Камеры разделены перегородками, пересечение которых с оболочкой приводит к образованию линии контакта или шва. Отверстия или отверстия, присутствующие в перегородках, позволяют прохождению пищи, сократительных вакуолей и ядер. Отверстия в стенке последней камеры соединяют внутреннюю эндоплазму с ложноножками и позволяют выпускать дочерние клетки. [ 14 ]
Оболочка может иметь стенки из органического материала, агглютинированные (образованные частицами, собираемыми организмом и зацементированными) или минерализованные (образованные минеральными веществами, выделяемыми организмом). У голых видов (амебоидные формы без оболочки, Athalamea ) тело клетки состоит из единственной камеры, ограниченной толстыми жилками, заключенными в слизистую и аморфную ткань. Самые ранние формы раковины (например, Allogromia ) имеют тонкую стенку, состоящую из белкового материала с небольшим количеством агглютинированного материала или без него. Переход между органическими и агглютинированными раковинами происходит у нескольких линий моноталамических фораминифер ( Monothalamea ). У видов с агглютинированными стенками частицы осадка, собираемые и цементируемые организмом, ограничивают аморфный или волокнистый органический матрикс. Наконец, в многокамерных или политаламических формах ( Tubothalamea и Globothalamea ) стенки содержат агглютинированный или минерализованный материал, включая кальцит , арагонит и кремнезем .
Ядро клетки и органеллы
Эндоплазма моноталамических фораминифер однородная, с равномерно распределенными клеточными органеллами. У polythalamus foraminifera первые камеры составляют хроматический сегмент, содержащий ядро и другие органеллы, в то время как последние камеры представляют собой вегетативные сегменты и отвечают за переваривание пищи. Клетка некоторых видов имеет одно ядро, которое может быть довольно большим, до 100 мкм в диаметре, в то время как другие виды содержат сотни небольших ядер, разбросанных по цитоплазме. У фораминифер со сменой поколений количество ядер зависит от поколения: гамонты обычно одноядерные, а шизонты они многоядерные. У некоторых фораминифер имеется два типа ядер: единственное соматическое ядро (которое участвует только в метаболических функциях) большое и занимает периферические камеры, а генеративные ядра (которые участвуют в образовании гамет) многочисленные, маленькие и занимают центральная часть оболочки.
Помимо ядра, доминирующими элементами, присутствующими в цитоплазме, являются большие пищеварительные вакуоли . Они образуются на внешней стороне скорлупы, охватывая пищу, хотя пищеварение происходит в основном внутри. Аппарат Гольджи похож на аппарат других протистов. И эктоплазма, и эндоплазма содержат множество мелких митохондрий , которые демонстрируют типичные трубчатые митохондриальные гребни многих протистов. [ 15 ] Другими клеточными органеллами, присутствующими в клетках фораминифер, являются микросомы и пероксисомы . , используется в обмене веществ и при разложении токсичных веществ.
Жизненный цикл
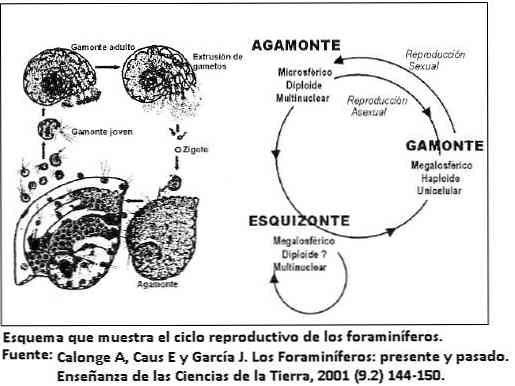
Жизненный цикл фораминифер часто включает смену поколений с бесполым и половым размножением. Чередование поколений приводит к двум различным морфологиям ( диморфизм ), которые очень похожи на невооруженный глаз. [ 16 ] [ 17 ] Таким образом, в репродуктивном цикле существует два разных поколения:
-
. Гаплоидный тип , одноядерный, микросферический (небольшого размера), с половым размножением по гамогонии . . Диплоидный тип , многоядерный, мегалосферный (или макросферный, большого размера) и с бесполым размножением при шизогонии .
Первоначально гамонте имеет одно ядро и делится на множество гамет, которые обычно имеют два жгутика. Шизонт многоядерный и после мейоза фрагментируется с образованием новых гамонтов. Нередко бывает несколько раундов бесполого размножения для каждого полового поколения среди бентосных форм. Фактически, среди нынешних популяций доля шизонтов намного выше, чем доля гамонтес, что указывает на более исключительное половое размножение. Микросферная форма консервативна в онтогенетическом смысле, так как воспроизводит филогенетическое развитие группы. Напротив, мегалосферная форма является прогрессивной, непосредственно начиная свое развитие в эволюционно продвинутых формах.
Самым обширным отрядом среди корненожек являются обитатели моря — фораминиферы (Foraminifera). В составе современной морской фауны известно свыше 1000 видов фораминифер. Небольшое число видов, представляющих, вероятно, остаток морской фауны, обитает в подпочвенных соленых водах и солоноватых колодцах Средней Азии.
Подобно раковинным корненожкам, все фораминиферы имеют раковину. Строение скелета достигает здесь большой сложности и огромного разнообразия.
В океанах и морях фораминиферы распространены повсеместно. Их находят во всех широтах и на всех глубинах, начиная от прибрежной литоральной зоны и кончая самыми глубокими абиссальными впадинами. Все же наибольшее разнообразие видов фораминифер встречается на глубинах до 200—300 м. Подавляющее большинство видов фораминифер является обитателями придонных слоев, входят в состав бентоса. Лишь очень немногие виды живут в толще морской воды, являются планктонными организмами.

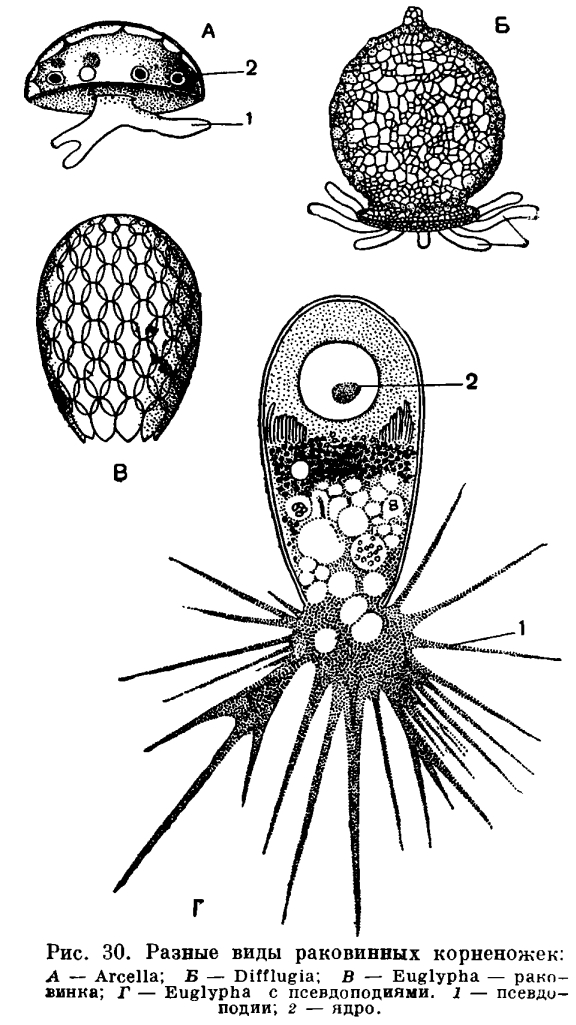
Среди огромного разнообразия строения раковин фораминифер можно различить по составу их два типа. Одни из них состоят из посторонних телу корненожки частиц — песчинок. Подобно тому как мы это видели у диффлюгии (рис. 30), фораминиферы, обладающие такими агглютинированными раковинами, заглатывают эти посторонние частицы, а затем выделяют их на поверхности тела, где они закрепляются в тонком наружном кожистом слое цитоплазмы. Такой тип строения раковины имеют часто встречающиеся представители родов Hyperammina, Astrorhiza (рис. 32, 3—7) и др. Например, в некоторых районах наших северных морей (море Лаптевых, Восточно-Сибирское море) эти крупные фораминиферы, достигающие 2—3 см длины, почти сплошным слоем покрывают дно.
Число видов фораминифер с агглютинированной раковиной относительно невелико (хотя число особей этих видов может быть огромным). Большая часть обладает известковыми раковинами, состоящими из углекислого кальция (СаС03).
Эти раковины выделяются цитоплазмой корненожек, которые обладают замечательной особенностью концентрировать в своем теле кальций, содержащийся в морской воде в небольших количествах (соли кальция в морской воде составляют немногим более 0, 1%). Размеры известковых раковин разных видов фораминифер могут быть очень различны. Они варьируют в пределах от 20 мк до 5—6 см. Это примерно такое же соотношение размеров, как между слоном и тараканом. Наиболее крупных из фораминифер, раковина которых имеет 5—6 см в диаметре, уже нельзя назвать микроскопическими организмами. Наиболее крупные (роды Cornuspira и др.) живут на больших глубинах.
Однокамерные фораминиферы имеют одну-единственную полость внутри раковины, которая сообщается с наружной средой устьем. Форма однокамерных раковин разнообразна. У одних (например, Lagena) раковинка напоминает бутылочку с длинным горлышком, иногда снабженную ребрами (рис. 32, 2).
Очень часто происходит спиральное закручивание раковины, и тогда внутренняя полость ее становится длинным и тонким каналом (например, Ammodiscus, рис. 32, 8, 9).
Большинство известковых раковин корненожек является не однокамерными, а многокамерными. Внутренняя полость раковины поделена перегородками на ряд камер, число которых может достигать нескольких десятков и сотен. Перегородки между камерами не сплошные, в них имеются отверстия, благодаря чему протоплазматическое тело корненожки не расчленено на части, а представляет собой единое целое. Стенки раковинок не у всех, но у многих фораминифер пронизаны мельчайшими порами, которые служат для выхода наружу псевдоподий. Об этом подробнее будет сказано ниже.
Число, форма и взаимное расположение камер в раковинке может быть очень различным, что и создает огромное разнообразие фораминифер (рис. 32). У некоторых видов камеры расположены в один прямой ряд (например, Nodosaria, рис. 32, 12), иногда же их расположение двурядное (Textularia, рис. 32, 22). Широко распространена спиральная форма раковины, когда отдельные камеры расположены по спирали, причем по мере приближения к камере, несущей устье, размеры их возрастают. Причины этого постепенного нарастания размеров камер станут понятны, когда мы рассмотрим ход их развития.
В спиральных раковинках фораминифер имеется несколько оборотов спирали. Наружные (более крупные) обороты могут быть расположены рядом с внутренними оборотами (рис. 32, 17, 18) так, что все камеры видны снаружи. Это эволютный тип раковины. У других форм наружные (более крупные) камеры целиком или частично охватывают внутренние камеры (рис. 33, 1). Это инволютный тип раковины. Особую форму строения раковины находим у фораминифер милиолид (семейство Miliolidae, рис. 32, 19). Здесь камеры сильно вытянуты параллельно продольной оси раковины и расположены в нескольких пересекающихся плоскостях. Вся раковина в целом оказывается продолговатой и по форме несколько напоминает тыквенное зерно. Устье расположено на одном из полюсов и обычно снабжено зубцом.
Большой сложностью строения отличаются раковинки, относящиеся к циклическому типу (роды Archiacina, Orbitolites и др., рис. 33, 2, 34). Число камер здесь очень велико, причем внутренние камеры располагаются по спирали, наружные же — концентрическими кольцами.
В чем заключается биологическое значение столь сложного строения многокамерных раковинок корненожек? Специальное исследование этого вопроса показало, что многокамерные раковинки по сравнению с однокамерными обладают гораздо большей прочностью. Основное биологическое значение раковинки — это защита мягкого протоплазматического тела корненожки. При многокамерном строении раковинки эта функция осуществляется весьма совершенно.
Внутренняя полость раковины заполнена цитоплазмой. Внутри раковинки помещается и ядерный аппарат. В зависимости от стадии размножения (о чем будет сказано несколько ниже) ядро может быть одно или их несколько. Из раковинки через устье наружу выдаются многочисленные очень длинные и тонкие псевдоподии, ветвящиеся и анастомозирующие между собой. Эти особые свойственные фораминиферам ложные ножки называют ризоподиями. Последние образуют вокруг раковинки тончайшую сеточку, общий диаметр которой обычно значительно превосходит диаметр раковинки (рис. 34). У тех видов фораминифер, у которых имеются поры, ризоподии выдаются наружу через поры.

Выполненные за последние годы с применением цейтраферной киносъемки исследования показали, что цитоплазма, входящая в состав ризоподии, находится в непрерывном движении. Вдоль ризоподии в центростремительном (к раковине) и центробежном (от раковины) направлениях довольно быстро текут токи цитоплазмы. По двум сторонам тонкой ризоподии цитоплазма как бы струится в противоположных направлениях. Механизм этого движения до сих пор остается не выясненным.
Размножение фораминифер происходит довольно сложно и у большинства видов связано с чередованием двух разных форм размножения и двух поколений. Одно из них бесполое, второе — половое. В настоящее время процессы эти изучены у многих видов фораминифер. Не вдаваясь в детали, рассмотрим их на каком-нибудь конкретном примере.

На рисунке 35 изображен жизненный цикл фораминиферы Elphidium crispa. Этот вид представляет собой типичную многокамерную фораминиферу со спирально закрученной раковинкой. Начнем рассмотрение цикла с многокамерной корненожки, обладающей маленькой зародышевой камерой в центре спирали (микросферическое поколение).
В цитоплазме корненожки первоначально имеется одно ядро. Бесполое размножение начинается с того, что ядро последовательно несколько раз делится, в результате чего образуется множество небольшого размера ядер (обычно несколько десятков, иногда свыше сотни). Затем вокруг каждого ядра обособляется участок цитоплазмы и все протоплазматическое тело корненожки распадается на множество (по числу ядер) одноядерных амебообразных зародышей, которые выходят через устье наружу. Сразу же вокруг амебовидного зародыша выделяется тонкая известковая раковинка, которая и явится первой (эмбриональной) камерой будущей многокамерной раковины. Таким образом, при бесполом размножении на первых стадиях своего развития корненожка является однокамерной. Однако очень скоро к этой первой камере начинают добавляться следующие. Происходит это так: из устья сразу выступает наружу некоторое количество цитоплазмы, которая тотчас же выделяет раковинку. Затем наступает пауза, в течение которой простейшее усиленно питается и масса протоплазмы его увеличивается внутри раковины. Затем вновь часть цитоплазмы выступает из устья и вокруг нее образуется очередная известковая камера. Этот процесс повторяется несколько раз: возникают все новые и новые камеры, пока раковина не достигнет характерных для данного вида размеров. Таким образом, развитие и рост раковины носит ступенчатый характер. Размеры и взаимное положение камер определяется тем, какое количество протоплазмы выступает из устья и как эта протоплазма располагается по отношению к предшествующим камерам.
Мы начали рассмотрение жизненного цикла Elphidium с раковинки, обладавшей очень маленькой эмбриональной камерой. В результате бесполого размножения получается раковинка, эмбриональная камера которой значительно крупнее, чем у той особи, которая приступала к бесполому размножению. В результате бесполого размножения получаются особи макросферического поколения, которые существенно отличаются от дающего им начало микросферического поколения. В данном случае потомство оказывается не вполне похожим на родителей.
Они развиваются в результате полового размножения макросферического поколения. Происходит это следующим образом. Как и при бесполом размножении, половой процесс начинается с деления ядра. Количество образующихся при этом ядер значительно больше, чем при бесполом размножении. Вокруг каждого ядра обособляется небольшой участок цитоплазмы, и таким путем формируется огромное количество (тысячи) одноядерных клеток. Каждая из них снабжена двумя жгутиками, благодаря движению которых клетки активно и быстро плавают. Эти клетки являются половыми клетками (гаметами). Они сливаются друг с другом попарно, причем слияние затрагивает не только цитоплазму, но и ядра. Этот процесс слияния гамет и есть половой процесс. Образующаяся в результате слияния гамет (оплодотворения) клетка носит название зиготы. Она дает начало новому микросферическому поколению фораминиферы. Вокруг зиготы сразу же по ее образовании выделяется известковая раковина — первая (эмбриональная) камера. Затем процесс развития и роста раковины, сопровождающийся увеличением числа камер, осуществляется по тому же типу, как и при бесполом размножении. Раковина получается микросферической потому, что размер зиготы, выделяющей эмбриональную камеру, во много раз меньше одноядерных амебовидных зародышей, образующихся при бесполом размножении. В дальнейшем микросферическое поколение приступит к бесполому размножению и вновь даст начало макросферическим формам.
На примере жизненного цикла фораминифер мы встречаемся с интересным биологическим явлением закономерного чередования двух форм размножения — бесполого и полового, сопровождающегося чередованием двух поколений — микросферического (развивается из зиготы в результате оплодотворения) и макросферического (развивается из одноядерных амебоидных зародышей в результате бесполого размножения).
Отметим еще одну интересную особенность полового процесса фораминифер. Известно, что у большинства животных организмов половые клетки (гаметы) бывают двух категорий. С одной стороны, это крупные, богатые протоплазмой и запасными питательными веществами неподвижные яйцевые (женские) клетки, а с другой — мелкие подвижные сперматозоиды (мужские половые клетки). Подвижность сперматозоидов обычно связана с наличием у них активно двигающегося нитевидного хвостового отдела. У фораминифер, как мы видели, морфологических (структурных) различий между половыми клетками нет. По строению своему все они одинаковы и благодаря наличию жгутиков обладают подвижностью. Здесь нет еще структурных различий, которые позволили бы различать мужские и женские гаметы. Такая форма полового процесса является исходной, примитивной.
Как уже было сказано, огромное большинство современных видов фораминифер являются придонными (бентическими) организмами, встречающимися в морях всех широт от прибрежной зоны вплоть до самых больших глубин мирового океана. Изучение распределения корненожек в океане показало, что оно зависит от ряда факторов внешней среды — от температуры, глубины, солености. Для каждой из зон типичны свои виды фораминифер. Видовой состав фораминифер может служить хорошим показателем условий среды обитания.

Широко распространенные в современных морях и океанах фораминиферы были богато представлены и в прежние геологические периоды начиная с самых древних кембрийских отложений. Известковые раковинки после размножения или смерти корненожки опускаются на дно водоема, где входят в состав отлагающегося на дне ила. Процесс этот совершается десятки и сотни миллионов лет; в результате на дне океана образуются мощные отложения, в состав которых входит несметное количество раковинок корненожек. При горообразовательных процессах, которые совершались и совершаются в земной коре, как известно, некоторые области дна океана поднимаются и становятся сушей, суша опускается и становится дном океана. Большая часть современной суши в различные геологические периоды была дном океана. Это относится в полной мере и к территории Советского Союза (за исключением немногих северных районов нашей страны: Кольский полуостров, большая часть Карелии и некоторые другие). Морские донные отложения на суше превращаются в горные осадочные породы. Во всех морских осадочных породах присутствуют раковинки корненожек. Некоторые же отложения, как например меловые, в основной своей массе состоят из раковин корненожек. Столь широкое распространение фораминифер в морских осадочных породах имеет большое значение для геологических работ, и в частности для геологической разведки. Фораминиферы, как и все организмы, не оставались неизменными. В течение геологической истории пашей планеты происходила эволюция органического мира. Изменялись и фораминиферы. Для разных геологических периодов истории Земли характерны свои виды, роды и семейства фораминифер. Известно, что по остаткам организмов в горных породах (окаменелостям, отпечаткам и т. п.) можно определить геологический возраст этих пород. Для этой цели могут быть использованы и фораминиферы. Как ископаемые они благодаря своим микроскопическим размерам представляют очень большие преимущества, так как могут быть обнаружены в очень небольших количествах горной породы. При геологической разведке полезных ископаемых (в особенности при разведке нефти) широко используется метод бурения. При этом получается колонка породы небольшого диаметра, охватывающая все слои, через которые прошел бур. Если эти слои представляют собой морские осадочные породы, то в них при микроскопическом анализе всегда обнаруживаются фораминиферы. Ввиду большой практической важности вопрос о приуроченности определенных видов фораминифер к тем или иным осадочным породам известкового возраста разработан с большой степенью точности.
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева . 1970 .
Фораминиферы отряд простейших подкласса корненожек(рассматривается также как самостоятоятельный класс саркодовых, входящий в надкласс корненожек). Известны с кембрия. Размеры обычно 0,1—1 мм, редко до 20 см. наружный скелет в виде раковинок, у большинства известковых, изредка хитиноидных или состоящих из агглютинированных посторонних частиц (песчинок и т. п.). Раковинки однокамерные и многокамерные, расположены в один или два ряда, по спирали, иногда ветвящиеся. Через устье и поры раковинок выдаются тончайшие ветвящиеся и анастомозирующие псевдоподии (ризоподии). Для них характерно чередование полового и бесполого размножения. свыше 4000 современных видов (с ископаемыми около 30 тыс. видов). Все они морские, преимущественно бентосные организмы (за исключением 2 планктонных семейств Globigerinidae и Globorotaliidae). Раковины образуют значительную, часть океанических илов, морскоих осадков и осадочных пород.
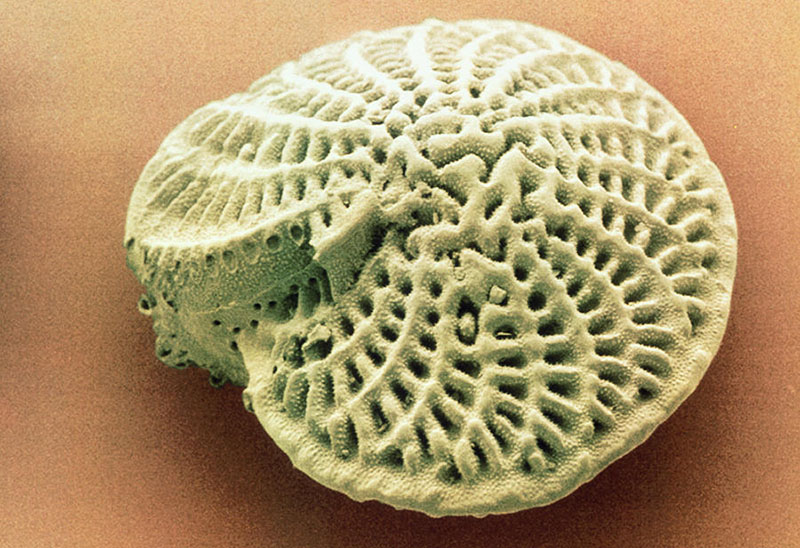
Фораминиферы строение раковина
Латинское название Foraminiferida
Фораминиферы Общая характеристика
Фораминиферы (лат. foramen – отверстие, дыра и fero – носить) – одноклеточные животные организмы, относимые к Простейшим, с сетью тонких разветвленных и соединяющихся между собой (анастомозирующих) псевдоподий (ложноножек). Это большая и разнообразная группа современных и ископаемых форм, насчитывающая в настоящее время более 34 000 видов, из которых около 4000 являются современными. Средние размеры их колеблются от 0,1 до 1 мм, однако некоторые формы могут достигать размера 10 мм и более. Цитоплазматическое тело фораминифер заключено в раковину различного состава и строения. Внутренняя полость раковины сообщается с внешней средой при помощи устья. Псевдоподии либо выходят только из устья раковины, либо образуют вокруг раковины сложную сеть, отходящую от наружной части цитоплазмы. Псевдоподии способны сокращаться и вытягиваться. В них осуществляется непрерывный ток цитоплазмы. По одной и той же псевдоподии одни струи ее текут в центростремительном (к раковине), другие – в центробежном направлении. Псевдоподии служат для улавливания и частично переваривания пищи, а также для передвижения животного.
Фораминиферы имеют сложный жизненный цикл, характеризующийся чередованием бесполого и полового поколений. Это в основном морские организмы. Небольшая часть фораминифер приспособилась к жизни в солоноватоводных бассейнах, а некоторые из них известны даже в пресноводных бассейнах. Большинство фораминифер являются бентосными формами (свободноживущими или прикрепленными), обитающими на разных глубинах. Некоторые ведут паразитический образ жизни на прикрепленных животных (мшанках, губках, кораллах). Планктонные формы значительно менее многочисленны; они живут на различных глубинах водного столба, будучи наиболее многочисленными в верхних пятидесяти метрах и особенно между 6 и 30 м от поверхности. Вертикальная миграция может быть суточной или в течение индивидуального развития.
Эти организмы распространены во всех морских бассейнах. Остатки их известны начиная с докембрия. Филогенетическое развитие сопровождалось усложнением строения раковины, которое было связано с прогрессивным усложнением строения и физиологических функций цитоплазматического тела.
Строение и физиология клетки

Форам.
Морфология раковин
Тип строения и терминология элементов раковины. Под типом строения подразумеваются характер последовательности нарастания камер и отчасти число камер, составляющих раковину. При этом один и тот же тип строения может иметь раковины как с агглютинированной, так и с секреционной стенкой. По числу камер все фораминиферы подразделяются на однокамерные, двухкамерные и многокамерные.
Однокамерные раковины имеют самую различную форму – трубчатую, трубчатую разветвленную или неправильно завернутую, звездчатую, древовидную, сферическую, полусферическую или субсферическую, колбовидную, лепешковидную. Они сообщаются с внешней средой с помощью одного или нескольких отверстий – устьев. У сферических и близких к ним форм имеется одно устье, расположенное в устьевой части (дистальном конце) раковины; часть раковины, противоположная устьевой, называется начальной (проксимальный конец). У трубчатых, звездчатых и других форм устьями являются свободные концы трубки.
Двухкамерные раковины состоят из начальной шарообразной камеры (пролокулюм) и второй трубчатой или ложнотрубчатой камеры. Трубчатая камера представляет собой настоящую трубку с собственной стенкой на всем протяжении, у ложнотрубчатой камеры в месте ее сочленения со смежным оборотом или с субстратом собственная минеральная стенка отсутствует, а имеется лишь тонкая органическая мембрана. Вторая – трубчатая или ложнотрубчатая – камера двухкамерных раковин может быть свернута в плоскую или коническую спираль или в неправильный клубок, у некоторых форм вторая камера представляет собой прямую несвернутую часть раковины или может быть свернутой зигзагообразно.
Стенка раковины у фораминифер, способ образования, состав и структура
Палеонтологи придают большое значение изучению состава и структуры стенки раковин фораминифер, рассматривая их в качестве ведущего признака в систематике этой группы организмов.
По составу и способу образования различают три типа раковин: органические, агглютинированные и секреционные известковые. Много новых данных по структуре раковин было получено за последние десятилетия благодаря использованию электронной микроскопии и рентгеноструктурного анализа.
Размножение, жизненный цикл, полиморфизм раковин
Система и характеристика высших таксонов
Система фораминифер находится в последнее время в состоянии активной разработки, но до сих пор отсутствует единое представление о таксономическом ранге этой группы и классификации ее на уровне высших таксонов. В настоящей работе принимается один из новых вариантов классификации этой гоуппы согласно изданию (Маслакова и др., 1995), учитывающий взгляды авторов предшествующих классификационных схем и новые представления. Здесь фораминиферы рассматриваются в ранге класса типа саркодовых. В составе класса выделяются 8 подклассов и 29 отрядов. При построении системы использовалась совокупность всех возможных критериев, т. е. морфологического, онтофилогенетического, геохронологического и экологического, которые определяют сочетание признаков, свойственных тому или иному таксону.
Основой классификации по-прежнему остается морфология раковины, поскольку, судя по имеющимся, хотя и немногочисленным, данным по цитологии и жизненным циклам, прогрессивное усложнение мягкого тела сопровождалось в эволюции этой группы многообразием и усложнением строения раковины. Наиболее важными признаками, положенными в основу выделения подклассов, являются определяющиеся природой цитоплазмы химический состав, способ образования и микроструктура стенки раковины. В качестве дополнительных признаков при выделении подклассов использованы такие особенности строения секреционных известковых раковин, как отсутствие или наличие и характер пористости в стенке раковины, наличие каналовой системы у специализированных форм, особая форма камер, специфический характер устья, образ жизни (экологический тип: бентосный или планктонный). Приняты во внимание данные по цитологии (особенности ядерного аппарата, присутствие в цитоплазме планктонных форм везикулярного ретикулума и криптосом), а также особенности жизненных циклов и характер гамет.
Для характеристики отрядов использовано сочетание таких признаков, как отсутствие или. наличие септации (однокамерные, двухкамерные и многокамерные раковины с учетом вторичного появления однокамерных форм у многих секреционных известковых форм), тип строения раковины или их определенное сочетание, преобладающий характер устья, форма камер, наличие каналовой системы, характер ранних стадий онтогенеза. При этом учитывалось, что таксономическое значение некоторых морфологических признаков не оставалось постоянным, а менялось в ходе исторического развития фораминифер (обычно повышалось).
Класс Foraminifera Orbigny, 1826
Саркодовые, тело которых заключено в различную по способу образования и составу раковину: органическую, агглютинированную или секреционную известковую (преимущественно кальцитовую или арагонитовую). Раковины имеют одно или несколько устьев.
Жизненные циклы с чередованием полового и бесполого поколений, с промежуточной редукцией; гаметы обычно двухжгутиковые, часто разножгутиковые (один из жгутиков с двумя рядами ресничек), реже трехжгутиковые, могут быть вторично-амебоидными.
Ядерный аппарат со сложными изменениями в течение жизненного цикла; лишь у низших форм часть цикла одноядерна, а у остальных многоядерность сохраняется в течение всего жизненного цикла (кроме зиготы). При этом у наиболее продвинутых форм возникает ядерный дуализм агамонтов (шизонтов), реже гамонтов.
Преимущественно морские бентосные свободноживущие, реже прикрепленные организмы; небольшая часть перешла к планктонному образу жизни, изредка наблюдается переход к паразитизму. Докембрий – современные.
Класс включает 8 подклассов: Allogromiata, Textulariata, Fusulinata, Miliolata, Lagenata, Rotaliata, Spirillinata, Globigerinata.
Экология и тафономия
Являются преимущественно морскими организмами, но кроме типично морских бассейнов некоторые их представители заселяют солоноватоводные, даже иногда пресные водоемы. Так, известны находки фораминифер в солоноватоводных и пресных бассейнах Сахары, ряд видов заходит на различное расстояние в эстуарии рек. Некоторые формы сохранились в виде реликтов в водоемах, отделившихся от моря и сильно опресненных (оз. Эльтон). Известны находки фораминифер в грунтовых водах и соленых колодцах Средней Азии. Предполагается, что это реликты миоценового моря, когда-то заходившего на территорию теперешних Каракумов. В условиях пониженной солености раковины фораминифер тонкие, при повышенной солености – толстые. Для фораминифер, живущих в условиях значительного опреснения, характерна утрата извести вплоть до образования чисто органической раковины.
Существуют две биономические группы фораминифер: бентосная и планктонная.
Этапы развития
Развитие фораминифер происходило в направлении усложнения морфологии раковины и появления различных типов стенки, а соответственно и новых таксонов. На основании смены систематического состава фораминифер в истории развития этой группы организмов установлены семь основных этапов:
- 1) раннепалеозойский, включающий ордовик и силур, к ним может быть; добавлен кембрий и частично докембрий;
- 2) среднепалеозойский (девон–ранний карбон);
- 3) позднепалеозойский (средний карбон – пермь);
- 4) – триасово-раннемеловой;
- 5) позднемеловой;
- 6) палеогеновый;
- 7) неоген-антропогеновый
Стратиграфическое значение
Фораминиферы имеют большое значение для разработки детальной стратиграфии, местной, межрегиональной и субглобальной корреляции. Это определяется тем, что их раковины в ряде случаев присутствуют в породе в больших количествах, благодаря чему возможны подробная послойная характеристика и датировка разреза. Кроме того, представители ряда групп фораминифер, такие, как фузулиниды, эндотириды, нуммулитиды, милиолиды, глобигериниды и другие, быстро эволюционировали, в связи с чем можно расчленять разрез на стратоны различного ранга по отдельным видам или их комплексам. Небольшие размеры раковин фораминифер позволяют извлекать их в достаточном количестве и из кернового материала, что придает им особую ценность при проведении буровых работ – расчленении и сопоставлении разрезов скважин.
Разные группы фораминифер дают возможность детально расчленять разрезы и выделять различные по широте распространения стратоны. Если по бентосным фораминиферам разрабатываются главным образом местные и региональные схемы, то по планктонным – субглобальные, простирающиеся в пределах одного климатического пояса. На основании изучения видов устанавливаются характерные комплексы для групп ярусов, отдельных ярусов, подъярусов; при более детальных исследованиях выделяются слои с видами или зоны, отвечающие обычно части подъяруса или подъярусу. При частом отборе образцов, когда улавливаются даже незначительные изменения фауны, и при детальном изучении филогении той или иной группы могут быть выделены филозоны, отражающие определенные этапы в эволюции этих организмов.
Хотя находки фораминифер известны начиная с докембрия и некоторое стратиграфическое значение имеют их более примитивные представители при расчленении силура и девона, но достаточно дробная зональная стратиграфия по этой группе ископаемых разработана начиная с карбона.
Схемы расчленения мезозойских и кайнозойских отложений первоначально были разработаны на материале континентов, в последние десятилетия более дробные схемы получены при исследовании из океанических скважин.
Фузулиниды имеют большое практическое значение для детального расчленения каменноугольных и пермских отложений таких регионов, как Поволжье, Западное Приуралье, Западная Европа, Северная Америка и др. Для разработки стратиграфии триасовых и юрских отложений главную роль играют лагениды (в триасе они составляют 70 % всех известных видов), а также милиолиды, эпистоминиды и в меньшей степени роталииды. Наиболее дробно стратиграфия триасовых отложений разработана для юга Западной Европы (особенно для Альп), юга бывшего СССР, Аляски. Для юры дробные стратиграфические схемы с выделением зон созданы для платформенных областей, в частности, для Восточно-Европейской и Западно-Сибирской платформ, и менее дробные для геосинклинальных областей.
В начале раннего мела в значительной степени наследуется систематический состав фораминифер позднеюрского времени, и руководящими формами на протяжении берриаса – баррема являются также лагениды, роталииды, эпистоминиды, некоторые атаксофрагмииды и литуолиды. Начиная с апта широкое распространение получают планктонные фораминиферы, которые делают возможным разработку зональной стратиграфии для отложений геосинклинальных областей. В областях развития платформенных отложений главная роль принадлежит роталиидам (Gavelinellidae и др.), лагенидам, эпистоминидам и агглютинированным формам.
Чрезвычайно велико практическое значение планктонных форм для разработки зональной стратиграфии верхнемеловых и палеогеновых отложений. Швейцарским микропалеонтологом Г. Болли в 1966 г. была предложена субглобальная зональная схема для меловых и кайнозойских отложений тепловодного пояса. Со временем эта схема совершенствовалась другими исследователями.
Наряду с планктоном большую роль в расчленении верхнемеловых и кайнозойских отложений играет и бентос. Так, зональная схема расчленения верхнего мела п-ова Мангышлак по аномалинидам широко применяется и для расчленения разрезов Восточно Европейской платформы; многочисленны работы по расчленению палеогеновых отложений Западной Европы и юга бывшего СССР по нуммулитидам.
Стратиграфия неогеновых и четвертичных отложений также разрабатывается как по планктонным, так и по бентосным фораминиферам. Среди последних наиболее важны милиолиды, различные роталииды (Rotaliidae, Elphidiidae, Nonionidae, Cassidulinidae).
Результаты изучения используются не только в стратиграфии, но и при решении вопросов фациального анализа, палеогеографии и палеозоогеографии. Отдельные экологические типы приурочены к определенным условиям разных зон морей и океанов. Это особенно хорошо устанавливается при исследовании современных фораминифер, полученные результаты с определенными поправками могут быть перенесены на ископаемый материал для восстановления палеообстановок.
Методика изучения
Раковины встречаются практически во всех породах морского происхождения, но преобладают в карбонатных глинах, мергелях, органогенно-обломочных известняках. Процесс исследования фораминифер состоит из сбора образцов в поле, технической обработки пород и научной обработки материала.

Фораминиферы Этапы развития
Развитие фораминифер происходило в направлении усложнения морфологии раковины и появления различных типов стенки, а соответственно и новых таксонов. На основании смены систематического состава фораминифер в истории развития этой группы организмов установлены семь основных этапов: 1) раннепалеозойский, включающий ордовик и силур, к ним может быть; добавлен кембрий и частично докембрий; 2) среднепалеозойский (девон–ранний карбон); 3) позднепалеозойский (средний карбон – пермь); 4) – триасово-раннемеловой; 5) позднемеловой; 6) палеогеновый; 7) неоген-антропогеновый (рис. 2.51).
Все эти этапы связаны с развитием земной коры: с горообразовательными процессами, колебательными движениями, трансгрессиями и регрессиями моря. В периоды трансгрессий происходили широкое распространение однородных условий, расселение фауны, обмен видами между бассейнами разных биогеографических провинций. С регрессиями связаны возникновение разнообразных условий, иногда неблагоприятных для существования некоторых групп морских организмов, изоляция фауны и резкие провинциальные отличия. В периоды регрессий вымирают высокоспециализированные элементы предшествующей фауны и открывается путь для развития прогрессивных элементов новых фаун.
Первый этап – ранний палеозой – приходится на каледонский цикл развития земной коры. В это время развитие и расселение фораминифер связаны с каледонскими геосинклиналями и областями морских трансгрессий на Восточно-Европейской и СевероАмериканской платформах.
Этап характеризуется существованием примитивных, преимущественно однокамерных форм с органической протеиновой раковиной, от которых в конце кембрия или начале ордовика независимо произошли однокамерные агглютинированные формы отряда Astrorhizida (подкласс Textulariata) и также однокамерные секреционные известковые формы с микрогранулярной стенкой отряда Parathuramminida (подкласс Fusulinata). Несколько позже среди агглютинированных форм появляются двухкамерные прямые или спирально свернутые раковины, в том числе спирально-плоскостные – отряд Ammodiscida, а среди секреционных – первые многокамерные (отряд Nodosinellida). Таким образом, уже на раннепалеозойском этапе развития у фораминифер возникли способности к агглютинации, секретированию кальцита, септация и связанная с ней многокамерность, спиральное нарастание трубчатой камеры.
Новым элементом в мезозойской фауне фораминифер являются также представители семейств Epistominidae и Ceratobuliminidae (надсемейство Robertinoidea) с арагонитовой раковиной, широко распространенные в юре и раннем мелу, а также такие роталоидные формы, как Discorbidae, Gavelinellidae.
Фораминиферы этого этапа присутствуют как в терригенных, так и в карбонатных фациях. К концу четвертого этапа планктонные фораминиферы приобретают глобальное распространение в пределах тепловодного климатического пояса.
Пятый этап –- позднемеловой – связан с широко распространившейся трансгрессией, дальнейшим формированием впадины Атлантического океана и других океанических впадин, что является благоприятным для развития и расселения планктонных фораминифер. Позднемеловая эпоха – это эпоха расцвета планктонных фораминифер, особенно таких семейств, как Globotruncanidae, Rotaliporidae, Rugoglobigerinidae и Planomalinoidae, убыстрения темпов их эволюции, появления новых морфологических признаков (кили, дополнительные устья). В позднем мелу планктонные фораминиферы распространились из эпиконтинентальных морей в открытый океан, где разные их группы адаптировались к различным глубинным уровням. При этом большого разнообразия и расцвета достигли не только Globigerinida, но и спирально-винтовые Heterohelicida.
На этом этапе существовали представители всех отрядов, известных в раннем мелу – конце предыдущего этапа, и появились представители трех новых отрядов. Из агглютинирующих – это Rzehakinida, возникшие еще в конце альба, но достигшие расцвета в позднем мелу, а из секреционных форм с известковой гиалиновой стенкой первые представители Orbitoidida и Nummulitida – высокоспециализированные формы, обладающие системой каналов и дополнительными внутренними скелетными образованиями, появившиеся в конце позднего мела.
Значительного разнообразия, по сравнению с предыдущим этапом, и расцвета достигают здесь атаксофрагмииды, некоторые текстулярииды и Gavelinellidae (последние принадлежат отряду Rotaliida). Отряды Buliminida, Rotaliida, Heterohelicida переживают в позднем мелу свой расцвет. В то же время значительно снижается роль представителей подкласса Lagenata.
Шестой этап развития фораминифер – палеогеновый – начинается с широкой регрессии и сокращения площадей морских бассейнов в датском и палеоценовом веках, сменившихся в эоцене трансгрессий. На границе мела и палеогена произошли значительные перестройки в органическом мире Земли – очередные кризисные явления в биоте и биосфере, которые ряд исследователей связывают с космическими причинами. На этом рубеже вымирают многие представители планктонных фораминифер, характерные для позднего мела, и появляются новые: Globorotaliidae, Globigerinidae, Hastigerinidae, Hantkeninidae и др. К. концу палеоцена почти исчезает другая группа планктонных фораминифер – Heterohelicida, игравшая большую роль в позднем мелу.
Однако самым ярким показателем палеогенового этапа являются Nummulitida, которые по своему геологическому значению могут быть приравнены к фузулинидам в позднем палеозое.
В палеогене продолжается расцвет Rotaliida (многочисленные семейства) и Bu-liminida; продолжают существовать разнообразные Ataxophragmiida, Textulariida и Lituolida и представители других отрядов, известных из позднего мела, но на уровне родов и семейств на рубеже мела и палеогена происходит значительная смена во всех этих группах.
Седьмой этап развития фораминифер – неоген-антропогеновый – проходил в исключительно разнообразных физико-географических условиях: мелководные и глубоководные, тепловодные и холодноводные, пониженной солености в полузамкнутых и даже замкнутых бассейнах.
В целом для этого этапа характерны большое разнообразие Miliolida, переживающих в неогене второй период расцвета, обилие представителей Rotaliida, среди которых наиболее характерны Elphidiidae, Nonionidae, Rotaliidae, Cassidulinidae, Anomalinidae, Discorbidae. Из крупных Orbitoidida в неогене существуют Miogipsinidae, пришедшие на смену палеогеновым Discocyclinidae и Lepidocyclinidae и вымершие в конце неогена. Значительно сокращается и число нуммулитид. Продолжают существовать известные из предыдущего этапа агглютинированные формы, а также Lagenida, Polymorphinida и Buliminida.
Планктонные фораминиферы по числу семейств менее разнообразны, чем в палеогене, здесь преобладают Globigerinidae, Catapsydracidae, Globorotaliidae и Hastigerinidae.
Перечисленные этапы развития фораминифер отражают общую картину их эволюции на уровне высших таксонов. Однако в зависимости от географического распространения и места обитания в морском бассейне одновозрастные фауны значительно отличаются друг от друга. В мезозое и кайнозое, так же как и в современных бассейнах, значительная разница в систематическом составе фораминифер наблюдается между климатическими поясами – тепловодным (Тетическим) и холодноводными (Бореальным и Нотальным), холодноводными и полярными (Арктическим и Антарктическим). Иногда эти различия так велики, например в позднеюрское время, что для отложений одного и того же времени устанавливаются разные ярусы – титонский в Тетическом поясе и волжский в Бореальном.
фораминифера это группа амебоидных простейших, некоторые морские и другие пресноводные. Они появились в начале Первой эры (кембрия), и их преемники заселили современные океаны. Они могут быть найдены от прибрежных районов (гипо или гиперсолевой) до дна океана, а также от тропиков до холодных арктических и антарктических океанов..
Его распределение зависит от нескольких факторов. Например, есть виды, которые поддерживают большие и частые изменения температуры, в то время как другие не могли выжить, поэтому термическая структура океанов отмечает важные различия между ассоциациями фораминифер..

Кроме того, глубина также является определяющим фактором в распределении фораминифер из-за их прямого попадания в свет. С другой стороны, давление связано с предыдущими факторами (температура и глубина), непосредственно влияющими на растворимость СО2, что влияет на секрецию карбоната кальция для образования оболочек.
С другой стороны, энергия воды на мелководных участках является важным элементом, поскольку она влияет на тип субстрата (жесткий или мягкий) и распределение питательных веществ..
Аналогичным образом, другие факторы, такие как соленость, мутность воды, pH, присутствие микроэлементов и / или органических компонентов, токи, скорость оседания, могут локально определять распределение фораминифер..
- 1 Известные виды
- 2 Характеристики
- 2.1 Факторы, влияющие на размер и морфологию фораминифер
- 4.1 Аталамеа
- 4.2 Моноталамея
- 4.3 Ксенофиофорея
- 4.4 Tubothalamea
- 4.5 Globothalamea
- 5.1-Размер
- 5.2 -Протоплазма
- 5.3-Скелет или оболочка
- 5.4 -Типы фораминифер
- 5.5 -Seudopods
- 9.1 Дело Маврикия
Известные виды
В настоящее время известно более 10 000 видов, а около 40 000 вымерли. Некоторые виды обитают на морском дне, то есть являются донными организмами, часто живущими в камуфляже, образующем часть эпифауны (epibetónicos), или они могут жить под песком (endobetónicos). По этой причине они также известны как живой песок.
Они также могут жить на растениях, в которых они покоятся как эпифиты, и даже многие из них выбирают сидячую жизнь, то есть они живут прикрепленными к субстрату на протяжении всего своего существования..
В то время как другие фораминиферы живут плавающими на разных глубинах в океане (от 0 до 300 м), то есть они являются планктоническими формами жизни, являющимися частью морского микропланктона. Эти формы более редки и менее разнообразны.
Крупнейшие и наиболее сложные планктонные фораминиферы чаще встречаются в тропических и субтропических условиях. В то время как в высоких широтах эти организмы обычно редки, меньше и очень простыми способами.
черты
Особенностью, которая выделяется в фораминиферах, является скелет или оболочка, структура, которая позволила, чтобы вымершие формы могли быть изучены в форме морских микрофоссилий, которые откладываются на дне моря.
Таким образом, оболочка является основным элементом для дифференцировки фораминифер и является единственной структурой организма, которая окаменела. Эти окаменелости очень много в морских отложениях, также участвуя в формировании осадочных пород.
Основными химическими соединениями оболочек являются кальцит, арагонит и кремнезем. Форма и размеры эмбриональной камеры зависят от ее происхождения, будь то продукт полового или бесполого размножения.
Во время их онтогенеза фораминиферы контролируют рост и размер камер. Этот контроль осуществляется через длину и расположение псевдоподиальных токов, поскольку псевдопод отвечает за формирование органической мембраны, которая предшествует минеральной оболочке..
Этот процесс очень важен для поддержания клеточных процессов, так как камера действует как биореактор.
Факторы, которые влияют на размер и морфологию фораминифер
Следует отметить, что размер и окончательная морфология, которую может принять фораминифера, зависят от нескольких факторов, в том числе:
- Форма и размеры зародышевой камеры.
- Количество стадий роста до взрослой стадии (то есть количество камер фораминифер).
- Форма камеры и ее модификации при онтогенезе.
- Расположение камер.
Более крупные фораминиферы обладают стратегическими моделями роста для поддержания объема постоянных камер без превышения оптимального размера. Эти стратегии заключаются в разделении камер на несколько отсеков, называемых кликами..
Эти кластеры расположены таким образом, чтобы обеспечить транспортные и регуляторные функции между протоплазмой внутри камер и снаружи. То есть все камеры и клики прекрасно взаимосвязаны.
Расположение камер может следовать прямолинейной или спиральной оси. Это будет зависеть от положения псевдоподиальных токов и расположения отверстия или отверстий в камере..
таксономия
Без дальности: SAR Супергруппа
Классы и заказы
- Athalamea (Reticulomyxida)
- Monothalamea (Allogromiida, Astrorhizida, Komokiida)
- Ксенофиофорея (Псамимид, Станномид)
- Tubothalamea(fusulinida, involutinida, miliolida, silicoloculinida, spirillinida)
- Globothalamea(Lituolida, Loftusiida, Schlumbergerinida, textulariida, Trochamminida, rotaliida, buliminida, globigerinida, robertinida, Carterinida, lagenida).
классификация
Хотя многое еще предстоит уточнить, мы можем выделить 5 классов:
Athalamea
Здесь вы можете найти фораминиферы, которые не имеют раковины или которые голые.
Monothalamea
Он включает бетонические фораминиферы, которые представляют собой органическую или агглютинированную оболочку с одной камерой.
ксенофиофоры
В этом случае фораминиферы представляют собой специализированный бетонный тип большого размера, многоядерный и с агглютинированной оболочкой. Как правило, они являются детритиворами или сапрофагами, то есть получают пищу из детрита или органических веществ в разложении..
Tubothalamea
Это включает в себя бетонные фораминиферы, которые имеют несколько трубчатых камер, по меньшей мере, на ювенильной стадии, которые могут быть намотаны по спирали, со связанной или известковой оболочкой.
Globothalamea
Эта классификация включает как бетонные, так и планктонные фораминиферы с многокамерными оболочками глобулярной, агглютинированной или известковой формы. Снаряды могут быть одноплодными, бисериадными, трисериадными или трокоспираладами.
Тем не менее, эта классификация находится в постоянной эволюции.
морфология
-размер
Размер фораминифер обычно составляет от 0,1 до 0,5 см, при этом некоторые виды могут измерять от 100 мкм до 20 см..
-протоплазма
Фораминиферы образованы протоплазматической массой, которая составляет клетку фораминиферы..
Протоплазма обычно бесцветна, но иногда может содержать небольшое количество органических пигментов, липидного материала, симбиотических водорослей или соединений железа, которые придают ей цвет.
Протоплазма состоит из внутренней части, которая называется эндоплазмой, и внешней части эктоплазмы..
В эндоплазме он защищен оболочкой, и в нем органеллы распределяются в виде пищеварительных вакуолей, ядра, митохондрий, гранул, аппарата Гольджи или рибосом. По этой причине его иногда называют гранулярной эндоплазмой. Эктоплазма прозрачна и оттуда отходят выдвигающиеся псевдопод.
Протоплазма ограничена снаружи органической мембраной, состоящей из наложенных друг на друга слоев мукополисахаридов.
Протоплазматическая масса расширяется из оболочки через одно или несколько отверстий (пор) и покрывает ее наружно (экстракамерная протоплазма), и таким образом образуются псевдоподии..
-Скелет или оболочка
Foraminifera навсегда фиксируют свою клеточную поверхность посредством строительства минерального скелета (оболочки).
Оболочка состоит из камер, разделенных перегородками, но в то же время они сообщаются друг с другом через межсоединительные отверстия, называемые фораминами, отсюда и название фораминифер. Химический состав скелета или оболочки делает их структуры, которые очень легко окаменели.
Внутренняя часть камер покрыта органическим материалом, очень похожим на хитин. Кроме того, оболочка может иметь основные отверстия; Вы также можете иметь поры наружу или не иметь их.
Минеральная оболочка может быть сформирована из одного компартмента (примитивного foraminifera или monotalamos) или камеры, которая растет непрерывно, или из нескольких камер, которые формируются в последовательных стадиях, в сложной системе прерывистого роста (foraminifera polylamas).
Этот последний процесс состоит в добавлении к ранее сформированной оболочке и в стратегических местах нового материала скелета..
Многие фораминиферы могут выбирать материал для формирования своей оболочки в соответствии с их химическим составом, размером или формой, поскольку краевые псевдоподиальные токи, которые находятся в контакте с субстратом, способны его распознать..
-Типы фораминифер
По форме конструкции раковины их можно разделить на три основных типа фораминифер:
Агглютинированный (или гнойный)
В этом типе раковин фораминиферы собирают своими псевдоподами большое количество органических материалов, доступных в окружающей среде, в которой они живут, которые впоследствии агглютинируют, такие как минеральные зерна, спикулы губок, диатомовые водоросли и т. Д..
Большинство агглютинированных фораминифер цементируют свою оболочку карбонатом кальция, но если это соединение не присутствует в окружающей среде, как, например, те, которые живут в глубоких районах океана, где кальций не существует, они могут сделать это с кремнистыми, железистыми, органическими цементами, и т.д.
porcelaneous
В этом случае оболочка образуется через иглы магнезиального кальцита, которые синтезируются в аппарате Гольджи фораминифера..
Эти иглы транспортируются и накапливаются снаружи и могут служить в качестве соединительных элементов для посторонних структур (цемента) или непосредственно формировать внешний каркас. Они обнаружены в гиперсоленых средах (> 35% солености).
Они обычно неперфорированные, то есть обычно имеют псевдопоры, которые не проходят через оболочку полностью.
стекловидный
Они образуются в результате роста кристаллов кальцита благодаря органическому шаблону, образованному процессом, называемым биоминерализация (минерализация in situ), проводимым снаружи к протоплазматическому телу..
Они характеризуются прозрачностью благодаря тонкости стены. Они также перфорированы там, где расположение, плотность и диаметр пор изменяются в зависимости от вида..
-seudópodos
Эта структура используется для мобилизации, прикрепления к подложкам, захвата добычи и создания скелета. Для ретракции и удлинения псевдоподов фораминиферы имеют сложную сеть микротрубочек, расположенных в более или менее параллельных рядах..
Удлинение псевдоподов может достигать двух-трехкратной длины тела и даже может быть до 20-кратной длины. Это будет зависеть от каждого конкретного вида.
Тип движения во время смещения напрямую связан с формой раковины и положением отверстий (откуда берутся псевдоподы)..
Но большинство фораминифер движутся следующим образом: псевдопод прикрепляется к субстрату и затем толкает остальную часть клетки. Движение таким образом может продвигаться со скоростью примерно от 1 до 2,5 см / час..
С другой стороны, псевдопод фораминифер называется Granurreticulopodia, потому что внутри псевдопод существует двунаправленный цитоплазматический поток, который несет гранулы.
Гранулы могут состоять из частиц различных материалов, митохондрий, пищеварительных или отходящих вакуолей, синбиотических динофлагеллят и т. Д. По этой причине одним из синонимов группы является Granuloreticulosa..
Другая важная характеристика псевдопод - это то, что они имеют тенденцию быть длинными, тонкими, разветвленными и очень обильными, образуя таким образом сеть ретикулоподий путем укладки (анастомоз).
Жизненный цикл
Жизненный цикл фораминифер обычно короткий, обычно несколько дней или недель, но в больших формах жизненный цикл может достигать двух лет..
Продолжительность будет зависеть от жизненной стратегии, которую принимает фораминифера. Например, маленькие формы с простой морфологией развивают короткую оппортунистическую стратегию.
Принимая во внимание, что крупные формы и с чрезвычайно сложной морфологией раковины развивают консервативную жизненную стратегию.
Это последнее поведение очень редко встречается у одноклеточных организмов; позволяет им поддерживать равномерную плотность населения и медленный рост.
воспроизведение
Большинство фораминифер имеют две морфологии с чередованием поколений в соответствии с типом размножения, половым или бесполым, за исключением планктонных фораминифер, которые размножаются только половым путем..
Это изменение в морфологии называется диморфизмом. Получающаяся форма полового размножения (гамогония) называется гамоном, а бесполое размножение (шизогония) дает форму шизонта. Оба морфологически различны.
Некоторые фораминиферы координируют цикл размножения с сезонным циклом, чтобы оптимизировать использование ресурсов. Нередко наблюдается несколько непрерывных бесполых репродукций до того, как половое поколение происходит в бетонных формах..
Это объясняет, почему формы шизона более распространены, чем у гамонтов. Изначально гамон представляет собой единое ядро, а затем делится, образуя многочисленные гаметы..
Пока шизонт многоядерный и после мейоза фрагменты образуют новые гаметы.
![]()
питание
Фораминиферы характеризуются гетеротрофностью, то есть питаются органическим веществом..
В этом случае фораминиферы питаются главным образом диатомовыми или бактериальными, но другие более крупные виды питаются нематодами и ракообразными. Заключенные попали в ловушку своих псевдоподов.
Кроме того, эти организмы могут использовать водоросли симбионтов различных типов, таких как зеленые, красные и золотые водоросли, а также диатомовые и динофлагелляты, и даже у одного и того же особи их может быть очень сложное разнообразие..
С другой стороны, некоторые виды фораминифер являются клептопластическими, что означает, что хлоропласты из проглоченных водорослей становятся частью фораминифер, чтобы продолжать выполнять функцию фотосинтеза..
Это представляет альтернативный способ производства энергии для жизни.
приложений
Обилие окаменелостей фораминифер по геологическому времени, эволюции, сложности и размеру делает их любимым инструментом для изучения настоящего и прошлого Земли (геологические часы).
Поэтому его большое разнообразие видов очень полезно при изучении биостратиграфического, палеоэкологического, палеоокеанографического типа..
Но это также может помочь предотвратить экологические катастрофы, которые могут повлиять на экономику, поскольку изменения в популяциях фораминифер указывают на изменения в окружающей среде..
Например, фораминиферы без оболочек чувствительны к изменениям окружающей среды и быстро реагируют на изменения окружающей их среды. Поэтому они являются идеальным индикаторным видом для изучения качества и здоровья рифовой воды..
Дело Маврикия
Кроме того, некоторые события заставили нас задуматься об этом. Так обстоит дело с явлением, наблюдаемым на Маврикии, где часть белого песка пляжа исчезла, и теперь они должны импортировать его с Мадагаскара, чтобы поддержать поток туристов..
И что там произошло? Откуда берется песок? Почему это исчезло??
Песок - это не что иное, как скопление оболочек из карбоната кальция многих организмов, в том числе фораминифер, которые тянутся к краю пляжа. Исчезновение песка было связано с постепенным и устойчивым сокращением производителей карбонатов.
Это произошло в результате загрязнения морей азотом и фосфором, которые достигают побережья из-за чрезмерного использования удобрений при посеве определенных продуктов, таких как сахарный тростник..
Вот почему важно изучать фораминиферы в социальных науках, чтобы предотвратить такие экологические катастрофы, как описанная выше, которые непосредственно влияют на экономику и общество..
Читайте также: