
Размножение бактерий спирохет грибов
Обновлено: 25.04.2024
Соотношение поверхность/объем у бактериальных клеток очень велико, что способствует быстрому поглощению питательных веществ из окружающей среды за счет диффузии и активного транспорта.
Поэтому в благоприятных условиях бактерии способны расти очень быстро. Рост бактериальных клеток в большой степени зависит от таких факторов среды, как температура, наличие питательных веществ, рН среды и концентрация ионов. Кроме того, облигатным аэробам необходим кислород, а облигатным анаэробам необходимо, чтобы его не было.
Достигнув определенных размеров, диктуемых соотношением объемов ядра и цитоплазмы, бактерии переходят к бесполому размножению путем простого деления, т. е. путем деления на две идентичные дочерние клетки.
Клеточному делению предшествует репликация ДНК, причем до тех пор, пока процесс репликации не завершится, мезосомы могут удерживать ДНК в определенном положении. Мезосомы могут прикрепляться и к новым перегородкам, образующимся между дочерними клетками, участвуя каким-то образом в синтезе материала клеточной стенки. У самых быстрорастущих бактерий деление происходит через каждые 20 мин.
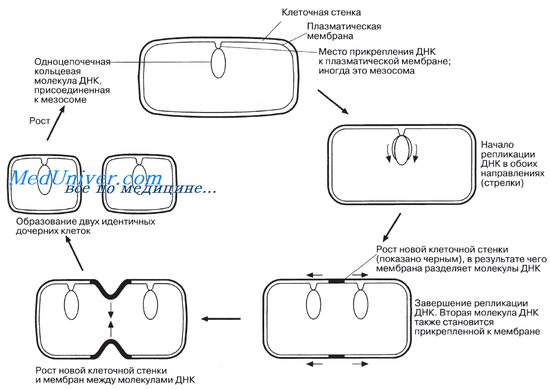
Бесполое размножение бактерий (например, Е. coli) простым делением.
Половое размножение бактерий
В 1946 г. у бактерий было обнаружено половое размножение, но в самой примитивной форме. Гамет в данном случае не образуется, однако наиважнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Генетическая рекомбинация впервые была обнаружена при изучении E.coli.
В норме при наличии в среде достаточного количества глюкозы и неорганических солей E.coli сама синтезирует все необходимые ей аминокислоты. В результате облучения этих бактерий у них иногда возникают случайные мутации. Были выделены два типа мутантов: один, не способный синтезировать биотин (витамин) и аминокислоту метионин, и другой — не способный синтезировать аминокислоты треонин и лейцин.
В среду, не содержавшую всех четырех факторов роста, помешали по 108 клеток каждого мутантного штамма. Теоретически клетки не должны были расти на этой среде. Однако все же было получено несколько сотен колоний (каждая колония возникает из одной исходной клетки), причем оказалось, что в таких клетках имеются все гены, необходимые для образования этих четырех факторов роста. Следовательно, в клетках каким-то образом произошел обмен генетической информацией, но выделить вещество, ответственное за этот процесс, в то время не удалось.
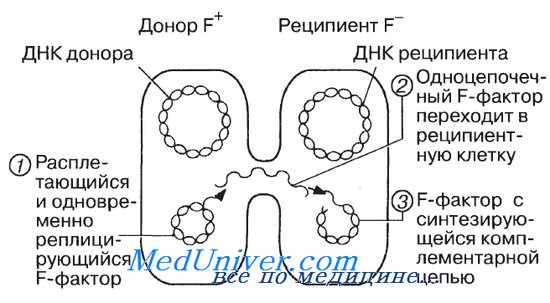
Конъюгация двух бактериальных клеток. Цифрами 1, 2 и 3 обозначены последовательные этапы переноса F-фактора.
В конце концов было установлено (при помощи электронного микроскопа), что клетки E.coli могут непосредственно контактировать друг с другом, т. е. у них может происходит конъюгация.
Пили — структуры полые и предполагается, что именно по этим пилям осуществляется перенос ДНК от донора (F+) к реципиенту (F-). Процесс этот показан на рисунке.
Обратите внимание на то, что донорная клетка сохраняет F-фактор, а реципиентная клетка его приобретает и становится F+. Процесс этот протекает медленно, и поэтому прежде чем перенос F-плазмиды завершится, клетка, бывшая изначально F-, успевает реплицироваться один или несколько раз, и в результате в популяции всегда сохраняются F-клетки.
F-фактор вызывает особенно большой интерес и потому, что время от времени, примерно в 1 случае из 100 000, он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации происходит перенос не только F-фактора, но и всей остальной ДНК. Этот процесс занимает примерно 90 мин, но клетки могут расходиться и раньше, чем произойдет полный обмен ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называют Hfr-штаммами (от англ. Н — High — высокая, f — frequency — частота, г — recombination — рекомбинация), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Бактерии: Жизнедеятельность бактерий характеризуется ростом — формированием структурно-функциональных компонентов клетки и увеличением самой бактериальной клетки, а также размножением — самовоспроизведением, приводящим к увеличению количества бактериальных клеток в популяции.
Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как и грибы, могут размножаться спорами. Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Актиномицеты, являясь ветвящимися бактериями, размножаются путем фрагментации нитевидных клеток Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, синтезируют поперечную перегородку от периферии к центру при участии мезосом. а грамотрицательные — путем перетяжки(на месте деления обнаруживается постепенно увеличивающееся искривление ЦПМ и клеточной стенки внутрь.), в результате образования гантелевидных фигур, из которых образуются две одинаковые клетки. При почковании на одном из полюсов материнской клетки формируется и растёт почка, материнская клетка проявляет признаки старения и обычно не может дать более 4 дочерних.
У других бактерий кроме размножения наблюдается половой процесс, но в самой примитивной форме. Половой процесс бактерий отличается от полового процесса эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового процесса, а именно обмен генетическим материалом, происходит и в этом случае. Это называется генетической рекомбинацией. Делению клеток предшествует репликация бактериальной хромосомы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается и каждая нить достраивается комплементарной нитью), приводящая к удвоению молекул ДНК бактериального ядра — нуклеоида. Репликация ДНК происходит в три этапа: инициация, элонгация, или рост цепи, и терминация.
Размножение спирохет: поперечное деление-деление клетки у бактерий, при котором материнская клетка дает начало двум дочерним клеткам. Осуществляется в три стадии:
1) репликация молекулы ДНК кольцевой хромосомы, присоединенной к мезосоме, которая так же делится на две части;
2) разведение с помощью мезосом двух дочерних кольцевых хромосом;
3) разделение цитоплазмы поперечной перегородкой, которая образуется от периферии к центру клетки.
Большинство грибов способно к вегетативному, собственно бесполому и половому размножению. Характерен плеоморфизм — наличие одновременно нескольких видов спороношений, например, бесполого и полового.
Вегетативное размножение
· Специализированными образованиями: артроспорами (оидиями) с тонкими стенками или хламидиоспорами с толстыми, образуются они, с некоторыми отличиями, при распаде мицелия на части, а затем дают начало новому.
· Почкование гиф или отдельных клеток (например, у дрожжей). Также почкуются аскоспоры у сумчатых и базидиоспоры у головнёвых. Образующиеся почки постепенно отделяются, растут и со временем сами начинают почковаться.
Бесполое размножение
Собственно бесполое размножение идёт посредством спор. В зависимости от способа образования различают эндогенные и экзогенные споры.
· Эндогенные споры (спорангиоспоры) характерны для низших грибов. Образуются внутри особых клеток, называемыхспорангиями.
· Экзогенные споры обычно называют конидиями ,они имеются у высших и у некоторых низших грибов. Образуются на вершинах или сбоку специальных гиф — конидиеносцев, ориентированных вертикально, которые могут быть простыми или разветвлёнными. Покрыты плотной оболочкой, поэтому довольно устойчивы, но неподвижны. Могут подхватываться воздушными потоками или животными и переноситься на значительные расстояния. При прорастании дают ростовую трубку, а затем гифы.
Половое размножение

Для низших грибов свойственно слияние гаплоидных гамет путём изогамии, анизогамии (гетерогамии) или оогамии. В случае оогамии развиваются половые органы — оогонии (женские) и антеридии (мужские). При оплодотворении происходит образование ооспоры — это зигота, которая покрывается толстой оболочкой, некоторое время проводит в состоянии покоя, после чего прорастает.
Скорость и фазы размножения бактерий в стационарных условиях.
При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или поверхностный (в виде пленки) рост культуры. Рост периодической культуры бактерий, выращиваемых на жидкой питательной среде, подразделяют на несколько фаз, или периодов:
1. лаг-фаза;
2. фаза логарифмического роста;
3. фаза стационарного роста, или максимальной концентрации бактерий;
4. фаза гибели бактерий.
Эти фазы можно изобразить графически в виде отрезков кривой размножения бактерий, отражающей зависимость логарифма числа живых клеток от времени их культивирования.
Лаг-фаза — период между посевом бактерий и началом размножения. Продолжительность лаг-фазы в среднем 4—5 ч. Бактерии при этом увеличиваются в размерах и готовятся к делению; нарастает количество нуклеиновых кислот, белка и других компонентов.
Фаза логарифмического (экспоненциального) роста является периодом интенсивного деления бактерий. Продолжительность ее около 5— 6 ч. При оптимальных условиях роста бактерии могут делиться каждые 20—40 мин. Во время этой фазы бактерии наиболее ранимы, что объясняется высокой чувствительностью компонентов метаболизма интенсивно растущей клетки к ингибиторам синтеза белка, нуклеиновых кислот и др.
Затем наступает фаза стационарного роста, при которой количество жизнеспособных клеток остается без изменений, составляя максимальный уровень (М-концентрация). Ее продолжительность выражается в часах и колеблется в зависимости от вида бактерий, их особенностей и культивирования. Завершает процесс роста бактерий фаза гибели, характеризующаяся отмиранием бактерий в условиях истощения источников питательной среды и накопления в ней продуктов метаболизма бактерий. Продолжительность ее колеблется от 10 ч до нескольких недель. Интенсивность роста и размножения бактерий зависит от многих факторов, в том числе оптимального состава питательной среды, окислительно-восстановительного потенциала, рН, температуры и др.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Для того чтобы изучать микроорганизмы, определять этиологические факторы инфекционных заболеваний, заниматься вопросами профилактики и лечения инфекционных заболеваний и решать многие другие вопросы, связанные с микроорганизмами, необходимо иметь их в достаточном количестве, а это значит - создавать все условия для нормального роста и размножения микроорганизмов.
Размножение микроорганизмов происходит путем поперечного деления, почкованием, образования спор, репродукции.
Рост микроорганизмов означает увеличение массы микробов в результате синтеза клеточного материала и воспроизведения всех клеточных компонентов и структур.
О бактериях, спирохетах, актиномицетах, грибах, риккетсиях, микоплазмах, простейших, хламидиях говорят, что они размножаются, а вирусы и фаги (вирусы микробов) – репродуцируются.
Размножение микроорганизмов соответствует определенным закономерностям. Скорость деления микроорганизмов различна, она зависит от вида микроба, возраста культуры, особенностей естественной и искусственной питательной среды, температуры, концентрации углекислого газа и многих других факторов.
В процессе размножения микроорганизмы на различных этапах претерпевают морфологические и физиологические изменения (по форме, размерам, окрашиваемости, биохимической активности, чувствительности к физическим и химическим факторам и пр.).
Микроорганизмы обладают возрастной изменчивостью, т.е. особи изменяются на разных стадиях роста, созревания и старения. Эти изменения наблюдаются в нормальном цикле индивидуального развития микроорганизма, который зависит от природы организма, от сложности его строения и последовательности в развитии.
Наиболее простым циклом развития среди микроорганизмов обладают бактерии. Размножаются они простым поперечным делением в различных плоскостях. В зависимости от этого клетки могут располагаться беспорядочно, гроздями, цепочками, пакетами, попарно, по четыре и т.д.
Характерной чертой бактерий, отличающей их от многочисленных животных и растений, является их необыкновенная скорость размножения.
Каждая бактериальная клетка в среднем в течение получаса претерпевает деление, что обусловлено усиленным обменом веществ, скоростью, с которой питательный материал поступает внутрь клетки.
Фактором, тормозящим размножение бактерий, является истощение питательного субстрата и отравление окружающей среды продуктами распада.
У бактерий различают восемь основных фаз размножения.
1. Исходная стационарная фаза, которая представляет собой период времени один – два часа от момента посева бактерий на питательную среду. В этой фазе размножение не происходит
2. Фаза задержки размножения (лаг – фаза), в течение которой размножение бактерий происходит очень медленно, а скорость их роста увеличивается. Продолжительность второй фазы около двух часов.
3. Фаза длится пять – шесть часов. Третья фаза характеризуется максимальной скоростью деления, уменьшением размеров клеток.
4. Фаза отрицательного ускорения (продолжается около двух часов). Скорость размножения бактерий снижается, число делящихся клеток уменьшается.
5. Стационарная фаза, длящаяся около двух часов. Число новых бактерий почти равно числу отмерших особей.
6. Фаза ускорения гибели клеток (длится около трех часов).
7. Фаза логарифмической гибели клеток (длится около пяти часов), при которой гибель клеток происходит с постоянной скоростью
8. Фаза уменьшения скорости отмирания. Оставшиеся в живых особи, переходят в состояние покоя.
Продолжительность фаз размножения не является постоянной величиной. Она может быть различной в зависимости от вида микроорганизмов и условий культивирования.
Цикл развития кокковидных бактерий сводится к росту клетки и последующему ее делению. Палочковидные аспорогенные бактерии в молодом возрасте растут, достигают максимума величины, затем делятся на две дочерние клетки, которые повторяют тот же цикл. У бацилл и клостридий в цикл развития включается при определенных условиях процесс спорообразования.
Спирохеты и риккетсии, как и бактерии, размножаются путем бинарного деления.
Среди микоплазм способностью размножаться обладают все элементарные тела сферической или овоидной формы. В процессе развития на элементарном теле появляется несколько нитевидных выростов, в которых формируются сферические тела. Постепенно нити становятся тоньше и образуются цепочки с четко выраженными сферическими тельцами. Затем происходит деление нитей на фрагменты и освобождение сферических телец.
Размножение некоторых микоплазм происходит путем отпочкования дочерних клеток от более крупных шаровидных тел. Поперечным делением микоплазмы размножаются, если процессы деления микоплазм идут синхронно с репликацией ДНК нуклеоида. При нарушении синхронности образуются нитевидные многонуклеоидные формы, в последующем делящиеся на кокковидные клетки.
Актиномицеты и грибы имеют две различные стадии развития: стадию вегетативного роста, при которой характерным является образования мицелия и стадию образования спор, формирующихся на спороносцах.
Важной особенностью актиномицетов и грибов является значительное разнообразие способов их размножения. Для них характерны вегетативное, бесполое и половое размножение.
Вегетативное размножение осуществляется путем деления на фрагменты гиф с последующим образованием отдельных палочковидныхи кокковидных клеток.
Бесполое размножение происходит вегетативным путем (рост фрагментов гиф или их отдельных клеток) и при помощи более или менее специализированных органов размножения (спор и конидий). Наиболее частый, бесполый, путь размножения проявляется в образовании экзогенных и эндогенных спор. Экзоспоры или конидии образуются на концах плодоносящих гиф, но заключены внутри общего мешочка – спорангия. Гифы, несущие спорангии, называются спорангионосцами. Спорангионосцы могут быть прямыми, волнистыми, спиральными.
Половое размножение происходит при помощи специальных органов – аскоспор, базидиоспор, образованию которых предшествует половой процесс. По биологическому назначению споры актиномицетов и грибов бывают покоящиеся, служащие для сохранения вида в течение определенного периода и служащие для быстрого размножения.
Споры актиномицетов и грибов образуются каждой особью в большом количестве, так как в отличие от спор бактерий служат, в основном, целям размножения. Они менее устойчивы к факторам окружающей среды, чем споры бактерий.
У простейших, так же как у актиномицетов и грибов, наряду с размножением путем деления существует и половой процесс.
Хламидии, вирусы и фаги имеют своеобразные циклы развития.
Размножение хламидий начинается с проникновения элементарных телец в чувствительную тканевую клетку путем эндоцитоза. Эти тельца в вакуоле клетки превращаются в вегетативные формы, называемые инициальными или ретикулярными тельцами, которые обладают способностью делиться. Ретикулярные тельца имеют пластинную клеточную стенку, а в цитоплазме – рыхло расположенные ядерные фибриллы и многочисленные рибосомы. После многократного деления ретикулярные тельца превращаются в промежуточные формы, из которых развивается новое поколение элементарных телец. Весь цикл развития хламидий длится 40 – 48 часов и заканчивается формированием микроколонии хламидий в цитоплазме клетки – хозяина.
После разрыва стенки вакуоли и полного разрушения клетки – хозяина, микроколонии хламидий, оказавшись за пределами целой клетки, распадается на самостоятельные элементарные тельца, и цикл проникновения хламидий в клетку с последующим их размножением повторяется.
Репродукция вирусов характеризуется последовательностью отдельных стадий.
1. Стадия адсорбции. Вирионы адсорбируются на поверхностных структурах клетки. При этом происходит взаимодействие комплементарных структур вириона и клетки, которые называются рецепторами.
3. Стадия разрушения внешней оболочки и капсида вириона при помощи протеолитических ферментов клетки – хозяина. У одних вирионов процесс разрушения их оболочки начинается еще на стадии адсорбции, у других – в пиноцитарной вакуоле, у третьих – непосредственно в цитоплазме клетки при участии тех же протеолитических ферментов.
4. Стадия синтеза вирусных белков и репликации нуклеиновых кислот. После полного или частичного освобождения вирусной нуклеиновой кислоты начинается процесс синтеза вирусных белков и репликация нуклеиновых кислот.
5. Стадия сборки или морфогенез вириона. Формирование вирионов возможно только при условии строго упорядоченного соединения вирусных структурных полипептидов и их нуклеиновой кислоты, что обеспечивается самосборкой белковых молекул вокруг нуклеиновой кислоты. У одних вирусов этот процесс происходит в цитоплазме, у других – в ядре клетки хозяина. У сложноорганизованных вирусов, имеющих внешнюю оболочку, дальнейшая сборка происходит в цитоплазме во время выхода их из клетки.
6. Стадия выхода вирионов из клетки – хозяина. Ряд сложных вирусов выходят из клетки – хозяина, при этом клетки в течение некоторого времени сохраняют жизнеспособность, а потом погибают. Простые вирионы выходят из клетки через образовавшиеся в ее оболочке отверстия, клетка – хозяин погибает, не сохраняя в течение какого – то времени жизнеспособность.
В некоторых случаях репродукция вирионов в клетках может происходить в течение многих месяцев и даже лет. Вирусы выделяются через клеточную оболочку. При делении таких клеток вирионы передаются дочерним клеткам, в свою очередь начинающим продуцировать вирусные частицы.
Существует три типа взаимодействия вируса с клеткой: продуктивный, абортивный и вирогенный.
Продуктивный тип взаимодействия заключается в образовании новых вирионов.
Абортивный тип взаимодействия может внезапно прерваться в стадии репликации вирусной нуклеиновой кислоты или синтеза вирусных белков, или морфогенеза вирионов.
Вирогенный тип характеризуется встраиванием (интеграцией) вирусной нуклеиновой кислоты в ДНК клетки, которая обеспечивает синхронность репликации вирусной и клеточной ДНК.
При репродукции фага также происходит адсорбция его на поверхности клетки (1 стадия) в результате взаимодействия аминогрупп белков, локализованных в периферической части хвостового отростка фага, и отрицательно заряженных карбоксильных групп на поверхности бактериальной клетки.
Различают обратимые и необратимые фазы адсорбции. Обратимая фаза характеризуется тем, что фиксированные фаги можно отделить от клетки путем энергичного помешивания или резко уменьшить концентрацию ионов. Освободившиеся фаги при сохраняют свою жизнеспособность.
В период второй необратимой фазы адсорбции фаг не отделяется от тела микробной клетки. Процесс адсорбции длится несколько минут. Под влиянием фермента, находящегося в хвостовом отростке фага, в теле микробной клетки на месте прикрепления фага образуется отверстие, через которое внутрь клетки проникает ДНК фага. Оболочка фага остается снаружи (2 стадия).
Некоторые фаги вводят свою нуклеиновую кислоту в клетку без предварительного механического повреждения клеточной стенки. В наступивший после проникновения в клетку нуклеиновой кислоты фага латентный период, осуществляется биосинтез фаговой нуклеиновой кислоты и белков капсида фага.
Происходит синтез ферментов, необходимых для репликации фаговой нуклеиновой кислоты и структурных белков фага (3 стадия).
В четвертой стадии происходит заполнение фаговой нуклеокислотой пустотелых фаговых частиц и формирование зрелых фагов. Осуществляется морфогенез фага.
В конце латентного периода происходит лизис зараженных микробных клеток и выход зрелых фаговых частиц (5 стадия).
Считают, что адсорбция фага длится 40 минут, латентный период – 75 минут. Весь цикл взаимодействия фага с микробной клеткой продолжается немногим больше трех часов.
Внедрение фага в микробную клетку не всегда сопровождается ее лизисом. Нередко взаимодействие фага с микробной клеткой ведет к образованию лизогенных культур.
По характеру взаимодействия с микробной клеткой различают умеренные и вирулентные фаги. Состояние лизогении вызывается умеренными фагами. Лизогенные микробные клетки являются устойчивыми к вирулентным фагам. Вирулентные фаги обуславливают формирование новых фагов и лизис микробной клетки.
Спирохеты — тонкие, длинные, извитые (спиралевидной формы) бактерии, отличающиеся от спирилл подвижностью, обусловленной сгибательными изменениями клеток. Спирохеты имеют наружную мембрану клеточной стенки, окружающую протоплазматический цилиндр с цитоплазматической мембраной. Под наружной мембраной клеточной стенки (в периплазме) расположены периплазматические фибриллы, которые, как бы закручиваясь вокруг протоплазматического цилиндра спирохеты, придают ей винтообразную форму (первичные завитки спирохет). Фибриллы прикреплены к концам клетки и направлены навстречу друг другу. Другой конец фибрилл свободен. Число и расположение фибрилл варьирует у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение (рис. 7). При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками.
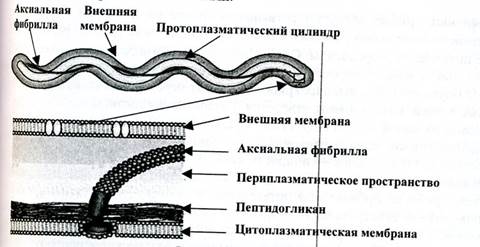
Рис. 7. Двигательный аппарат спирохет
Спирохеты плохо воспринимают красители из-за большого количества липидов в оболочке. Их окрашивают по методу Романовского-Гимзы или серебрением, а в живом виде исследуют с помощью фазово-контрастной или темнопольной микроскопии. Спирохеты представлены тремя родами, патогенными для человека: Тгероnеmа, Воrreliа, Leptospira.
Лептоспиры (род Leptospira, вид L.interrogans, возбудитель лептоспироза) имеют завитки неглубокие и частые - в виде закрученной веревки (завитки первого порядка). Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв "S" или "С"; имеют две осевые нити. Окрашиваются в красно-розовый цвет по методу Романовского-Гимзы.
Таблица 6 Дифференциальные признаки спирохет
5. Грибы
Грибы — это эукариоты, относящиеся к царству Fungi (Муcetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофилльные) микроорганизмы с клеточной стенкой. Широко распространены в природе, особенно в почве.
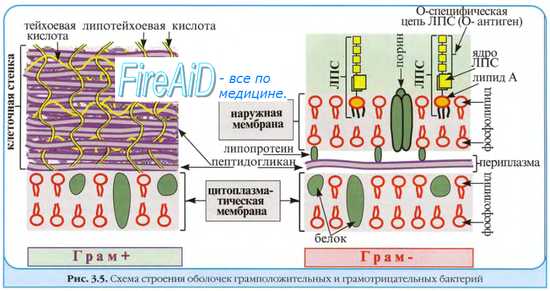
Грибы имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную ригидную клеточную стенку, состоящую из нескольких типов полисахаридов (маннанов, глюканов, целлюлозы, хитина), а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы (в отличие от холестерина — главного стерола тканей млекопитающих). Грибы являются грамположительными микробами, вегетативные клетки — некислотоустойчивые. Тело гриба называется талломом. Различают два основных типа грибов: гифальный и дрожжевой.
Гифальные (плесневые грибы)образуют ветвящиеся тонкие нити (гифы). Гифы (от греч. hypha, паутина) представляют собой разветвленные микроскопические трубки диаметром 2-10 мкм, содержащие цитоплазму и органеллы. Совокупность гиф обозначают термином мицелий (от греч. туkes — гриб и helos — нарост). Образование мицелия — отличительный признак истинных грибов (Еumycota). Гифы высших грибов содержат перегородки (септы), разделяющие их на отдельные клетки. Септы имеют отверстия, позволяющие цитоплазме и отдельным органеллам перетекать из одной клетки в другую. Гифы низших грибов не имеют перегородок и называются ценоцитными, или асептированными. Таким образом, плесневый гриб представляет собой ценоцит (от греч. koinos— общий и kytos — клетка) — обширную территорию цитоплазмы с множеством ядер, располагающуюся в скоплении трубок-гиф. Врастающая в субстрат часть тела гриба, абсорбирующая питательные вещества, — вегетативный мицелий, а растущая на поверхности субстрата часть — воздушный мицелий. Воздушный мицелий придает поверхности колоний плесневых грибов характерную шерстистую или пушистую фактуру. Нередко воздушный мицелий образуют специализированные гифы, несущие репродуктивные структуры.

Таблица 7 Грибы, имеющие медицинское значение
| Таксоны | Основные роды | Болезни людей |
| Зигомицеты (тип Zygomycota) Низшие грибы (имеют несептированный мицелий, размножение половое и бесполое) | Mucor, Rhizopus, Rhizomucor | Зигомикоз (мукоромикоз) |
| Аскомицеты (тип Ascomycota) Сумчатые грибы, большинство имеют септированный мицелий | Дрожжи: Saccharomyces | Многочисленные микозы |
| Arthroderma (Trichophyton, Microsporum) | Дерматомикозы | |
| Aspergillus, Penicillium | Аспергиллез, пенициллоз | |
| Базидиомицеты (тип Basidiomycota) | Дрожжи: Filobasidiella (Cryptococcus neoformans) | Криптококкоз |
| Дейтеромицеты (формальная группа Deiteromycota) | Несовершенные дрожжи: Candida, Cryptococcus, Trichosporon | Кандидозы, криптококкоз |
| Coccidioides (C.immitis) | Кокцидоидомикоз |
Дрожжевые и дрожжеподобные грибы.Дрожжи и дрожжеподобные грибы представлены отдельными овальными клетками размером 3-10 мкм, морфологически сходными между собой. При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий), состоящие из цепочек удлиненных клеток. Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называются дрожжевыми или дрожжеподобными. Они размножаются только бесполым способом — почкованием или делением. На питательных средах дрожжи и дрожжеподобные грибы образуют блестящие, выпуклые колонии, сходные с колониями бактерий.
Многие грибы обладают диморфизмом — способностью давать мицелиальный или дрожжеподобный рост в зависимости от различных факторов (например, условий культивирования).
У грибов выделяют половой и бесполый типы размножения. Бесполое размножение реализуется путем образования конидий и спорангиоспор, содержащих весь генетический материал, необходимый для возникновения и развития новой колонии. Половым путем образуются аскоспоры, базидиоспоры и зигоспоры.
Среди грибов, имеющих медицинское значение, выделяют три типа (Phylum) или отдела, имеющие половой способ размножения (совершенные грибы): зигомицеты (Zygomycota), аскомицеты (Askomycota) и базидиомицеты (Ваsidiomycota). Кроме того, выделяют условный тип/группу грибов — дейтеромицеты (Deiteromycota), у которых имеется только бесполый способ размножения (несовершенные грибы).
Нитевидные инволюционные формы бактерий. Размножение бактерий. Принципы размножения бактерий. Репликация бактерий.
Некоторые палочковидные бактерии могут образовывать нитевидные формы, особенно в неблагоприятных условиях. Формирование подобных структур осуществляется путём соединения отдельных клеток с помощью слизи, мостиков или специальных полисахаридных футляров. Многие патогенные бактерии способны также образовывать нитевидные и даже ветвящиеся структуры при нарушениях условий роста и регуляции деления. После нормализации условий существования бактерии восстанавливают нормальные форму и размеры (реверсируют к исходному фенотипу). Следует помнить, что подобные инволюционные формы особенно часто наблюдают при культивировании на искусственных питательных средах либо при проведении антибактериальной терапии.
Размножение бактерий. Принципы размножения бактерий. Репликация бактерий.
Бактерии размножаются поперечным (бинарным) делением с образованием двух идентичных особей. Делению предшествует удвоение бактериального генома, известное как репликация. Каждая цепь двойной спирали ДНК представляет собой линейную полярную последовательность дезоксирибонуклеотидов, расположенных в строго определённом порядке. Полярность цепи связана с тем, что каждый нуклеотид имеет 5'-фосфатный конец и З'-гидроксильный конец. При соединении нуклеотидов образуется цепь, также имеющая 5'-фосфатный и 3'-гидроксильный конец, причём цепи двойной спирали имеют противоположную полярность, то есть антипараллельны (рис. 2-6). Информационное содержание обеих цепей идентично (цепи гомологичны, или комплементарны), так как каждая из них содержит последовательность нуклеотидов, строго соответствующую последовательности другой цепи благодаря образованию водородных связей между парами аденин-тимин и гуанин-цитозин.
Репликация начинается в определённой точке ДНК и осуществляется ферментами ДНК-полимеразам и одновременно на каждой матричной цепи в двух противоположных направлениях, поскольку ДНК-полимеразы могут считывать матрицу только в направлении 3'-5'. При этом на одной из матричных цепей синтез ДНК происходит непрерывно (образуется ведущая дочерняя цепь), а на другой — короткими фрагментами длиной около 100 нуклеотидов, названными по имени открывшего их учёного фрагментами Оказаки (отстающая цепь). Синтез всегда начинается с РНК-затравки, которая затем удаляется, а фрагменты сшиваются лигазами (рис. 2-7). В результате репликации число молекул ДНК в клетке удваивается. Вновь синтезированные молекулы ДНК постепенно расходятся в образующиеся дочерние клетки. Всё это позволяет дочерней клетке иметь тождественную материнской клетке молекулу ДНК. Считают, что репликация ДНК занимает почти 80% времени, в течение которого осуществляется деление бактериальной клетки.
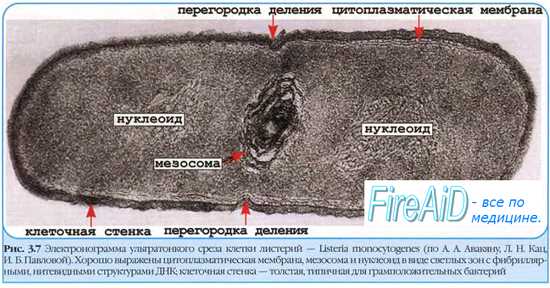
После завершения репликации ДНК начинается образование межклеточной перегородки; вначале с обеих сторон клетки происходит врастание двух слоев ЦПМ, а затем между ними синтезируется пептидогликан и образуется перегородка, состоящая из двух слоев ЦПМ и пептидогликана. Во время репликации ДНК и образования перегородки бактериальная клетка непрерывно растёт. В этот период происходят синтез пептидогликана клеточной стенки, ЦПМ, образование новых рибосом и других органелл и соединений, входящих в состав цитоплазмы.
На последней стадии деления дочерние клетки отделяются друг от друга. Процесс разделения клеток у некоторых бактерий идёт не до конца; в результате образуются цепочки клеток. При делении грамположительных бактерий образуется перегородка, отрастающая от клеточной стенки к центру клетки. У видов рода Mycobacterium перегородка образуется внутри клетки, затем расщепляется на два слоя и разделяет одну клетку на две. Грамотрицательные бактерии (включая риккетсии) истончаются в центре и разделяются перегородкой на две клетки. Бактерии рода Francisetla размножаются способом, напоминающим почкование у дрожжей (см. далее).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: