
Репликация ретровирусов осуществляется по схеме
Обновлено: 18.04.2024
Репликация ДНК. Механизмы
Деление клеток происходит посредством митоза, при этом, во избежание потери генетической информации, вначале удваивается весь ядерный геном в S-фазе клеточного цикла. Продолжительность S-фазы составляет 8 ч. ДНК центромер хромосом реплицируется во время средней стадии митоза, что предшествует процессу сегрегации хромосом.
Репликация митохондриальной и ядерной ДНК происходит в разные фазы клеточного цикла. Несмотря на то что общая последовательность стадий при репликации ядерной ДНК у высших существ (эукариот) и у бактерий (прокариот) одинакова, сам процесс имеет незначительные отличия. Так, у эукариот во время репликации ДНК (ядерная) остаётся в нуклеосомной конфигурации.
Гены с большим содержанием пар А—Т, кодирующие различные свойства и функционирующие лишь в определённых клетках, входят в состав факультативного гетерохроматина. Их репликация происходит на ранней стадии S-фазы только в тех клетках, в которых они экспрессируются, и на поздних стадиях — в клетках, где экспрессии не происходит.
Область спирали ДНК, которая в начале репликации раскручивается в первую очередь, называют участком начала репликации (репликоном). В этом месте двойную нить расплетает фермент хеликаза, раскрывающий последовательность оснований. Процесс репликации происходит вдоль одной цепи со скоростью примерно 40-50 нуклеотидов в секунду одновременно в обоих направлениях. У высших существ имеется множество репликонов, расположенных на расстоянии 50 000—300 000 п.н. В месте разделения нити ДНК возникают репликационные вздутия, на каждом конце которого формируется репликационная вилка.
Новая ДНК синтезируется при участии ферментов, называемых ДНК-полимеразами, из дезоксирибонуклеотидтрифосфатов (АТФ, ГТФ и др.), которые превращаются в монофосфатные нуклеотиды (АМФ, ГМФ и др.). Отщепление и гидролиз пирофосфатов из трифосфатов обеспечивают процесс энергией и обусловливают его полную необратимость, делая молекулу ДНК достаточно устойчивой.
Все ДНК-полимеразы могут выстраивать новую ДНК только в направлении от 5'- к 3'-концу. Это означает, что ферменты должны двигаться вдоль матричной цепи от 3'- к 5'-концу. В связи с этим репликация может непрерывно происходить от репликона только по одной цепи, называемой опережающей. Из-за расположения Сахаров репликация по второй, отстающей цепи происходит только на коротких отрезках, известных как фрагменты Оказаки.
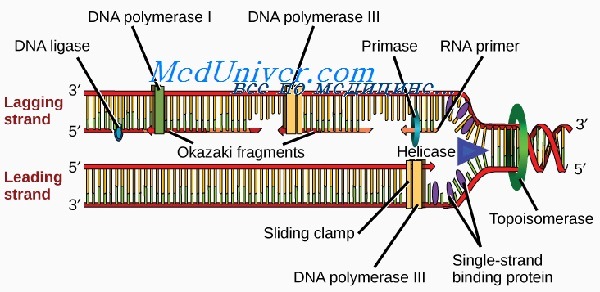
Длина новых фрагментов ДНК, образующихся вдоль отстающей цепи, в среднем составляет 100—200 пар нуклеотидов. Во время синтеза фрагменты Оказаки сшивает между собой фермент ДНК-лигаза. В ожидании репликации стабильность первичной одноцепочечной нуклеотидной последовательности отстающей цепи поддерживается белком, связывающим одноцепочечную ДНК (или спиральдестабилизирующим белком).
Для синтеза опережающей цепи необходим фермент ДНК-полимераза S, а для синтеза отстающей — ДНК-полимераза а. Последняя имеет субъединицу, называемую ДНКпраймазой, которая синтезирует короткую РНК-затравку, играющую роль праймера. Репликация мито-хондриальной ДНК происходит независимо от процессов в ядре. При этом используется ряд других ферментов, один из которых — ДНК-полимераза у.
В геноме присутствует большое количество копий пяти гистонных генов, благодаря чему происходит синтез множества гистонов (особенно во время S-фазы), которые сразу после репликации связываются с новой цепью ДНК.
Следует отметить, что процесс репликации носит название полуконсервативного, так как в состав дочерних молекул ДНК входит одна первичная цепь и одна синтезированная.
Репликация теломер ДНК
Основной проблемой синтеза ДНК на конце отстающей цепи служит то, что ДНК-полимеразе а необходимо прикрепиться выше конца последовательности, которая реплицирована, и работать проксимально в направлении от 5'- к 3'-концу. Для решения этой проблемы нужен ДНК-синтетический фермент теломераза, который продлевает отстающую цепь.
Теломераза — рибонуклеопротеин, содержащий матричную РНК с последовательностью 3'-ААУЦЦЦААУ-5', которая комплементарна полутора повторам шестиосновной теломерной ДНК (5'-ГГГТТА-3'). Фрагмент последовательности 3'-ААУ РНК-теломеразы связывается с терминальным концом ТТА-5' матричной отстающей цепи, при этом остальная часть РНК остаётся свободной. Затем к этой матричной РНК присоединяются дезоксирибонуклеотиды, тем самым продлевая повторяющуюся последовательность в ДНК на один сегмент.
После этого теломераза отщепляется и направляется к другому терминальному концу с последовательностью ТТА-5', и процесс повторяется. Как только возникает достаточно длинный терминальный повтор, ДНК-полимераза а прикрепляется к полученному одноцепочечному фрагменту и достраивает вторую цепь по методу комплементарности в проксимальном 5'—3'-направлении, двигаясь к уже существующему двухцепочечному участку, последующее слияние с которым происходит благодаря действию ДНК-лигазы.
Репаративные механизмы ДНК
Иногда в растущую цепь случайно вклинивается неправильное основание, однако, к счастью, у здоровых клеток присутствуют пострепликационные репаративные ферменты и система коррекции ошибочного спаривания оснований, которые исправляют подобные ошибки. В основе механизма действия данных систем лежат удаление и замена ошибочно вставленных оснований в соответствии с последовательностью матричной цепи. Для их функционирования необходимы ДНК-полимеразы b и е.
Значение ДНК для медицины. Патология пострепликационных механизмов репарации иногда обусловливает предрасположенность пациентов к некоторым онкологическим заболеваниям. К ним относят синдром множественной ломкости хромосом (синдром Блума), наследственную предрасположенность к раку молочной железы, вызванную мутациями генов BRCA1 и BRCA2, и аутосомно-доминантную форму рака кишечника (наследственный неполипозный рак толстой кишки).
Существует теория, утверждающая, что после каждого клеточного цикла теломеры укорачиваются на один повтор, а следовательно, количество делений клетки ограничено числом повторов в теломерной цепи. Согласно этому бесконечный рост и деление опухолевых клеток происходят из-за присутствия активных мутантных теломераз, которые препятствуют разрушению теломер.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
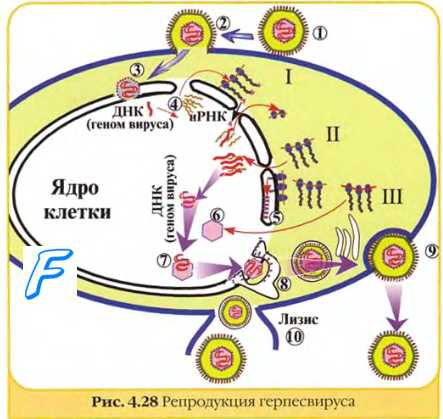
Основные отличия репродуктивного цикла герпесвирусов от остальных ДНК-вирусов связаны с более сложной структурой генома. Адсорбция вирусов на клетках осуществляется через специфические рецепторы. После взаимодействия с рецепторами вирусная оболочка сливается с клеточной мембраной, а нуклеокапсид высвобождается в цитоплазму. Раздевание (депротеинизация) вирусного генома происходит на ядерной мембране, и вирусная ДНК оказывается в ядре клетки-хозяина. Репродукция включает раннюю и позднюю стадии, однако они разграничиваются нечётко.
Поздняя стадия репродукции герпесвирусов. В позднюю стадию вирусная ДНК-полимераза индуцирует репликацию материнской ДНК. В результате образуются молекулы ДНК дочерней популяции. Часть дочерней ДНК считывают клеточные полимеразы, что вызывает транскрипцию концевых генов, кодирующих структурные протеины (белки оболочки и гликопротеины шипов).
Сборка дочерних популяций герпесвирусов осуществляется в ядре, где капсидные белки окружают молекулы ДНК, формируя нуклеокапсиды. Финальная стадия морфогенеза герпесвирусов — формирование суперкапсида на внутренней поверхности ядерной мембраны. Зрелые дочерние популяции отпочковываются от модифицированной ядерной мембраны, транспортируются через цитоплазму и выделяются наружу.
Поксвирусы. Репродукция поксвирусов.
Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется более 100 различных белков, входящих в состав вирионов (большинство образует наружную оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
• Транскрипция ДНК начинается до полной депротеинизации вируса, так как она полностью осуществляется вирусными полимеразами.
• Репликация происходит только в цитоплазме и полностью независима от клеточных полимераз, так как, в отличие от прочих вирусов, поксвирусы имеют собственную ДНК-зависимую РНК-полимеразу, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий.
В репродуктивном цикле выделяют три стадии— начальную, раннюю и позднюю.
Начальная стадия репродукции поксвирусов запускается сразу же после раздевания вируса и выхода вирусной ДНК в цитоплазму.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Репликация вирусных РНК молекул. Этапы репликации РНК генома вирусов.
Репликация вирусных РНК является уникальным феноменом. Существенное отличие механизма синтеза вирусных РНК от механизма синтеза клеточных РНК состоит в том, что в качестве матрицы в первом случае используется РНК, а во втором — ДНК.
Для транскрипции РНК на РНК-матрице необходима вирионная РНК-зависимая РНК-полимераза. Репликация вирусной РНК требует, прежде всего, синтеза комплементарной РНК, которая затем служит матрицей для производства большого количества вирусной РНК.
Когда вирусная РНК имеет отрицательную полярность (орто-, парамиксо-, рабдо-, фило-, борна-, арена- и буньявирусы), комплементарная РНК будет иметь положительную полярность и РНК-полимераза, подобно вирионной транскриптазе, используется для первичной транскрипции мРНК.
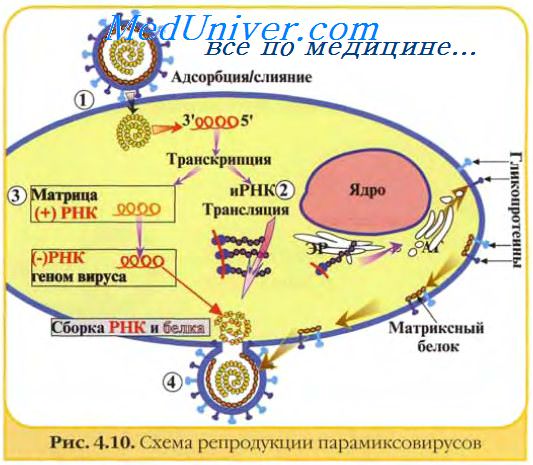
Так как большинство транскриптов, синтезируемых на каждой вирусной (-)цепи РНК, являются молекулами субгеномной РНК, некоторые полноразмерные цепи служат матрицами для синтеза (репликации) вирусной РНК. Некоторые вирусы для транскрипции и репликации используют различные РНК-полимеразы, тогда как в других случаях одни и те же ферменты могут выполнять различные функции.
У многих РНК-вирусов, (пикорна-, калици-, астро-, тога-, флави-, корона-, артери-, нодавирусы) комплементарная РНК является отрицательно полярной. На одной комплементарной РНК-матрице может транскрибироваться одновременно несколько молекул вирусной РНК, а на каждом РНК-транскрипте начинается продукция полимеразы. Образуется структура, известная как реплика-тивный посредник, — частично двуцепочечная структура с одноцепочечными хвостами.
Для начала репликации РНК пикорнавирусов и калицивирусов, а также ДНК аденовирусов требуется небольшой белок, связанный ковалентно с 5'-концом вновь синтезированных (+) или (—) цепей РНК, так же как с родительской вирионной РНК, но не с мРНК.
Вновь синтезированные (+)РНК могут иметь разное назначение: включаться в репликативный комплекс и служить матрицей для синтеза комплементра-ных (—)РНК; выполнять функции мРНК; включаться в качестве генома в новые вирионы. Механизм, определяющий судьбу вновь синтезированных (+)РНК, не известен.
Ретровирусы имеют геномную (+) одноцепочечную РНК. В отличие от других РНК-вирусов, они реплицируются посредством ДНК-посредника. Вирионная обратная транскриптаза, используя РНК-молекулу как праймер, создает односпиральную ДНК-копию. Затем, функционируя как рибонуклеаза, тот же самый фермент удаляет родительскую молекулу РНК из ДНК-РНК-гибрида и копирует одноцепочечную ДНК-цепь, чтобы образовать линейную двуцепочечную ДНК, которая содержит дополнительную последовательность, известную как длинный концевой повтор (LTR) на каждом конце.
Эта двуцепочечная ДНК затем циркулирует и интегрирует с клеточной хромосомальной ДНК. Вирусная РНК транскрибируется с интегрированной (провирусной) ДНК.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Тип геномной вирусной ДНК определяет особенности ее репликации.
1. Двунитевая ДНК – репликация происходит по обычному механизму полуконсервативной репликации: нити разделяются, и на каждой из них достраивается комплементарная ей нить.
3. У вирусов, геном которых представлен однонитевой РНК, ее репликация происходит по следующей схеме: вначале на вирионной РНК (вРНК) синтезируются комплементарные ей РНК (кРНК). Этот процесс катализируется специфической РНК-репликазой I. Затем на кРНК синтезируется комплементарная ей, но идентичная исходной вирионная РНК (вРНК), этот процесс также катализируется специфической репликазой II. Таким образом, репликация идет по схеме (рис. 80. II):
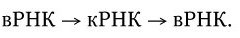
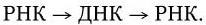
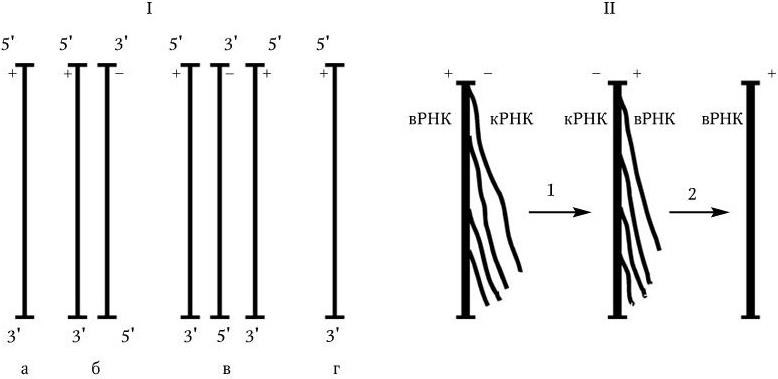
Рис. 80
. Механизм репликации однонитевого вирусного ДНК-генома (I) и однонитевого вирусного РНК-генома (II):
а – геномная однонитевая ДНК; б – репликативная форма; в – промежуточная репликативная форма; г – однонитевая дочерняя геномная ДНК; вРНК – вирионная (геномная) РНК; кРНК – комплементарная РНК. Объяснение в тексте
5. Размножение вируса гепатита В также протекает с участием обратной транскриптазы, но вначале клеточная РНК-полимераза синтезирует на вирусной ДНК прегеномную РНК, после чего вирусная ревертаза синтезирует на ней минус-цепь ДНК, которая достраивается плюс-цепью ДНК. В виде двунитевой ДНК вирус интегрируется в хромосому клетки-хозяина, где на ней транскрибируется вирусная РНК.
Существуют некоторые общие закономерности размножения вирусов. Во-первых, все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов, размножаются в цитоплазме. Для своего размножения вирусы гриппа А и В и ретровирусы проникают в ядро, что связано с особенностями поведения их генома. Во-вторых, размножение всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает в ядре, где происходит транскрипция и репликация их геномных нуклеиновых кислот, и в цитоплазме, где происходит трансляция вирусных белков, их процессинг и морфогенез вирионов. Лишь размножение вирусов группы оспы происходит в цитоплазме клетки, поскольку они обладают собственными системами транскрипции.
Другая особенность размножения вирусов заключается в том, что их нуклеокапсидные белки синтезируются на свободных полирибосомах (не связанных с мембраной), а суперкапсидные белки – на рибосомах, ассоциированных с мембранами (на шероховатых мембранах). Кроме того, белки некоторых вирусов подвергаются протеолитическому процессингу и гликозилированию. Различают два типа протеолитического процессинга: каскадный и точечный.
При точечном протеолизе разрезанию подвергается один (реже несколько) из вирусных полипептидов. Разрезание происходит, как правило, в определенном участке полипептида. Такой тип протеолиза необходим для того, чтобы определенный белок вируса приобрел свою специфическую активность. Например, суперкапсидный белок вируса гриппа – гемагглютинин – разрезается на две субъединицы: боEльшую и меньшую. В результате меньшая субъединица приобретает способность сливаться с мембранами клетки-мишени и ее лизосомами. Благодаря этому вирус гриппа приобретает способность проникать в клетку. Такой точечный протеолиз наблюдается у ортомиксовирусов, парамиксовирусов, ротавирусов, вирусов группы оспы и др. Точечный протеолиз, как и каскадный, жизненно важен для вируса.
Наконец, еще одна особенность вирусов, обладающих суперкапсидом, заключается в том, что суперкапсидные белки подвергаются в ходе своей транспортировки на наружную поверхность клеточной мембраны гликозилированию.
Читайте также: