
Семейство парвовирусов у животных
Обновлено: 18.04.2024
Парвовирусы (Parvoviridae; лат. parvus маленький + вирусы) — семейство, объединяющее самые мелкие и просто организованные ДНК-содержащие вирусы позвоночных и насекомых.
Семейство Парвовирусов состоит из трех родов: парвовирусов (Parvovirus), аденоассоциированных (аденосателлитных) вирусов (см. Аденоассоциированные вирусы) и денсовирусов (Densovirus). Роль Парвовирусов в патологии человека не изучена. Размер (диаметр) вирионов Парвовирусов колеблется от 18 до 26 нм. Им присуща кубическая симметрия икосаэдральной) типа, капсид состоит из 32 капсомеров, имеющих в поперечнике 3—4 нм. Молекулярный вес (масса) вирионов 5,5—6,2 X 106 дальтон, плавучая (удельная) плотность 1,38—1,46 г/см 3 , обнаруживаемые порой в культурах пустые капсиды имеют плотность 1,33—1,36 г/см 3 ;, константа седиментации 110—120 S.
В отличие от других ДНК-содержащих вирусов геном П. представлен однонитчатой ДНК с мол. весом 1,5—2,2 X 106 дальтон, составляющей 19—39% от общего веса вириона.
Белки П. представлены тремя полипептидами, молярное соотношение к-рых 10:1:1, причем на один вирион приходится ок. 60 молекул основного полипептида с мол. весом 62 000 — 66 000 дальтон, два других полипептида имеют мол. вес 72 000 — 87 000 и 46 000—55 000 дальтон.
Парвовирусы высокоустойчивы к физическим и химическим воздействиям; они выдерживают нагревание при t° 60° в течение 1 часа, а иногда и дольше (аденоассоциированные вирусы — до 4 час.), не инактивируются хлороформом, эфиром и анионными детергентами, устойчивы в кислой среде. Инактивируются УФ-облучением. Ряд П. обладает гемагглютинирующими свойствами; часто реакция гемагглютинации (см.) идет лишь с эритроцитами определенных видов животных, причем в нек-рых случаях только при t° 4°. В эксперименте показано, что П., находящиеся в кровяном русле животных, могут адсорбироваться на поверхности эритроцитов.
Размножаются П. в ядре инфицированной клетки, проявляя зависимость от фаз клеточного деления. Так, при изучении репродукции типового вируса этого семейства рода Parvovirus — латентного вируса крыс (син. вирус Килхэма) — было показано, что данный процесс осуществляется лишь в клетках, находящихся в стационарной фазе. При этом эффективность формирования зрелых вирионов зависит от интенсивности клеточного метаболизма. Это подтверждается уменьшением способности клеток крысиного эмбриона поддерживать образование латентного вируса крыс после предварительного рентгеновского или УФ-облучения, угнетающего синтез клеточных ДНК. Аналогичный эффект был получен и при использовании 5-фторурацила, также обладающего ингибирующим воздействием на клеточные ДНК. Аденоассоциированные вирусы размножаются только при наличии вирусов-помощников, в качестве к-рых выступают как аденовирусы и вирусы герпеса, так и нек-рые П., напр, латентный вирус крыс.

Сравнительная электронограмма латентного вируса крыс рода Parvovirus (1) и аденовируса (2); негативное контрастирование; X 260 000.
Латентный вирус крыс имеет небольшие размеры (рис.). Этот вирус вызывает у потомства естественно зараженных крыс врожденные заболевания. Внутриутробное инфицирование может привести к рассасыванию, плода, тяжелому гепатиту у новорожденных или к развитию более легких видов заболевания, допускающих выживание. В последнем случае у части выжившего потомства развивается гипоплазия мозжечка. Кроме того, при гистологическом исследовании в эндотелиальных клетках капилляров зараженных новорожденных крысят обнаруживают характерные тельца-включения.
Вирусы рода Parvovirus выделены от человека, а также млекопитающих и птиц, у к-рых в естественных условиях инфекция зачастую протекает в латентной форме. Порой лишь только тщательное обследование животных выявляет широкое распространение у них П. Выделить Парвовирусов удается из различных органов больных животных. Напр., П. кошек выделяется из содержимого желудка, ткани кишечника и селезенки взрослых животных, а у плодов — из мозжечка.
Патогенность вирусов в экспериментальных условиях ограничивается естественно-восприимчивыми хозяевами, но могут поражаться и близкие к ним виды. Так, П. кошек были выделены от тигра. В эксперименте латентный вирус крыс оказался патогенным для хомяков 1—4-дневного возраста, к-рые при заражении в мозг погибали на 3—6-й день. При инфицировании небольшими дозами вируса хомяки выживали, но отставали в росте. Куриные эмбрионы к П. невосприимчивы. Парвовирусы, патогенные для определенных видов животных, проявляют тропизм к соответствующим клеткам, на к-рые они оказывают цитопатическое действие.
Библиография: Феннер Ф. и др. Биология вирусов животных, пер. с англ., т. 1 — 2, М., 1977; Шалунова Н. В. и др. Совершенствование методов выявления латентной парвовирусной инфекции у крыс, Вестн. АМН СССР, № 7, с. 56, 1977: Berns К. I. Molecular biology of the adeno-associated viruses, Curr. Top. Microbiol. Immunol., v. 65, p. 1, 1974; Siegl G. The parvoviruses, Wien—N. Y., 1976.
Инфекции вызванные парвовирусами: диагностика, лечение, профилактика
Парвовирусы — мелкие двадцатигранные однонитевые безоболочечные ДНК-вирусы (5,6 тыс. пар нуклеотидов; 8—26 нм) с икосаэдрическим типом симметрии нуклеокапсида. Единственный представитель парвовирусов, способный вызвать заболевание у человека, — эритровирус (В19) из одноимённого рода Erythrovirus.
Эпидемиология инфекции вызванной парвовирусами. Вирус распространён по всему миру; сезонность не характерна. Путь передачи инфекции — воздушно-капельный. Чаще всего вспышки инфекционной эритемы (пятая болезнь) регистрируют в детских учреждениях (школах и др.). Доминирование парвовируса В19 усиливается с увеличением возраста (антитела определяют у 60% взрослых людей и более).
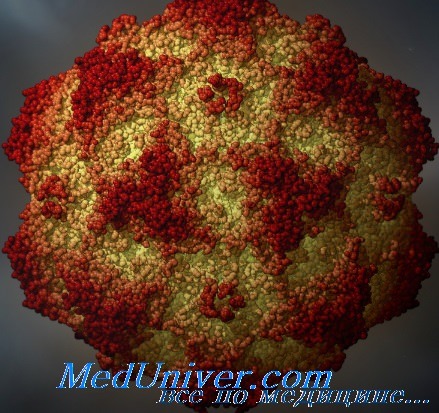
Возбудитель парвовирусной инфекции - парвовирус
Иногда (у взрослых пациентов) обнаруживают симметричный артрит мелких суставов. Угнетение эритропоэза легко переносят здоровые люди, но у пациентов с усиленным обновлением эритроцитов отмечают прогрессирующую анемию (например, апластический криз у пациентов с серповидно-клеточной анемией). Риск развития заболевания во время беременности достаточно низок, но в случае заражения возможно развитие отёчного синдрома новорождённых или даже смерть плода.
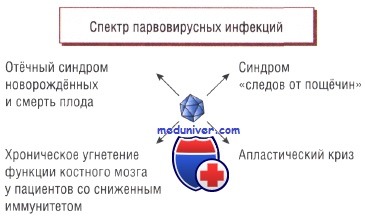
В настоящий момент не существует доказательств того, что парвовирус способен вызывать нарушения внутриутробного развития плода, но при заражении во II триместре в 10% случаев наблюдают самопроизвольное прерывание беременности.
Диагностика инфекции вызванной парвовирусами. Диагноз основан на данных клинической картины, результатах молекулярно-генетических методов. Культивировать парвовирусы достаточно сложно. Иногда используют ИФА (обнаружение IgM) и электронную микроскопию крови (в том числе пуповинной), смывов из носоглотки, амниотической жидкости.
Профилактика и контроль инфекции вызванной парвовирусами. В настоящее время вакцины против парвовирусов не существует. Для предотвращения распространения инфекции в лечебном учреждении необходимо соблюдать правила респираторной изоляции.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Полиомавирусы. Папилломавирусы. Виды полиомавирусов и папилломавирусов.
Семейство полиомавирусы включает вирусы полиомы, обезьян (SV40, SA12), кроликов (RKV), мышей (К) и человека (ВК, JC). Вирионы полиомавирусов представляют собой безоболочные частицы кубической симметрии диаметром 45 нм. Капсид имеет двадцатигранную структуру, состоящую из 72 капсомеров, расположенных на вершинах икосаэдра, из которых 60 являются гексонами и 12 - пентонами. Липидов и углеводов в составе вирионов не обнаружено. Полиомавирусы содержат одну циркулярную суперспирализованную ДНК размером 5 тпн.
Белки составляют 88—90% массы вириона. В вирионах SV40 и других полиомавирусов обнаружено три полипептида с молекулярной массой 21—48 Kd.
Полиомавирусы обладают высокой хозяинной специфичностью, и многие из них способны вызывать опухоли как в естественных условиях, так и при экспериментальном заражении.
Капсиды полиомавирусов содержат три белка VP1, VP2 и VP3, которые окружают одну молекулу ДНК. Основным белком полиомавирусов является полипептид VP1, на долю которого приходится примерно 70% всех белков вириона; он тесно связан с вирусной ДНК. Структурные белки VP2 и VP3 находятся в вирионе в небольшом количестве: VP3 составляет шестую-седьмую часть молекулярного количества VP1, a VP2 — четвертую часть молекулярного количества VP3. Установлено, что VP3 является С-концевой частью белка VP2. Белки VP2, VP3 вируса SV40 гомологичны соответствующим белкам вируса полиомы на 34%, а основной белок капсида (VP1) - на 54%.
Вирусная ДНК объединена с клеточными гистонами Н2А, Н2В, НЗ и Н4 в форме хроматина. Икосаэдральный капсид, содержащий 360 молекул главного капсидного белка VP1, организован в 72 пентаметра, одиночные молекулы VP2 или VP3 связаны с каждым пептаметром. Полиомавирусы известны как мелкие ДНК опухолевые вирусы. Эти вирусы могут трансформировать клетки первичной культуры, превращая их в перевиваемые линии и вызывать опухоли у животных.

Синтез ДНК полиомавирусов начинается с одного уникального места репликации и продолжается двунаправленно, вдоль циркулярной родительской ДНК. Двунаправленная репликация, пройдя полный круг циркулярной геномной ДНК, завершается образованием потомства циркулярной ДНК, которое отделяется.
Вирионы формируются в ядре и освобождаются с гибелью клеток. Некоторые клетки проявляют характерный цитопати-ческий эффект с вакуолизацией цитоплазмы. В инфицированной клетке могут синтезироваться 10 000—100 000 вирионов.
Около 60% сывороток КРС, включая сыворотки эмбрионов и новорожденных телят, содержат палиомавирус КРС, который хорошо размножается в культуре клеток почки обезьян. В Голландии около 60% ветеринарных работников имели антитела к этому вирусу.
Папилломавирусы
Мелкие ДНК вирусы, которые индуцируют бородавки у различных высших позвоночных, включая человека. Папилломавирусам свойственна латенция. Спонтанная регрессия папиллом является необычным явлением. Она не предсказуема, и папилломы могут появиться вновь. Папилломавирусы различаются по спектру хозяев и нуклеотидной последовательности ДНК. Папилломавирусы включают вирусы папилломы кроликов (папиллома Шоупа, прототипный вирус), папиллом человека, крупного рогатого скота, лошадей, оленей, овец, коз, собак и хомяков. Новым типом вируса считается вирус, у которого гомология ДНК с другими вирусами того же вида < 50%. Если гомология >50%, но < 100%, вирус считается новым подтипом и обозначается порядковым номером. Согласно этому критерию идентифицировано 6 типов папилломавирусов КРС, 2 типа у лошадей и 77 типов у человека. Папилломавирусы обнаружены у других видов животных (обезьян, собак, оленей, слонов, мышей и др.). ДНК папилломавирусов различных видов животных обладает лишь небольшой степенью гомологии. Различают две группы ПВ КРС: вирусы 1, 2 и 5 связаны иммунологически, имеют одинаковый размер генома и сходную нуклеотидную последовательность ДНК; вирусы 3, 4 и 6 имеют небольшие геномы и сходную нуклеотидную последовательность ДНК и связаны лишь отдаленно.
За немногими исключениями, папилломавирусы не размножаются в культуре клеток, но могут вызывать трансформацию клеток in vitro. Частота трансформации клеток обычно не превышает 2—3%.
ДНК многих папилломавирусов инфицирует животных. Инфекционные вирионы образуются только в эпителиальных клетках на конечной стадии их дифференциации.
Папилломавирусы — безоболочные сферические вирионы икосаэдрической симметрии диаметром 45 нм. Капсид состоит из двух структурных белков. Основной капсидный белок L1 (~55 КД) составляет около 80% массы вирионного белка. Минорный белок L2 имеет массу ~70 КД. Геном представлен циркулярной двухцепочечной суперспирализованной ДНК размером 8 тпн с ковалентно замкнутыми концами, обладающей инфекционностью. Вирулентная ДНК связана с клеточными гистонами, образуя хроматинподобный комплекс. Папилломавирусы могут образовывать дефектные интерферирующие частицы (Диг).
Репродукция вируса происходит в ядре клетки. В процессе репликации транскрибируется только одна цепь вирусной ДНК. Интегрированная эписомальная ДНК вируса может быть онкогенной.
Репликация папилломавирусов тесно связана с ростом и дифференциацией клеток кожи или слизистой оболочки. Вирусиндуцированная гиперплазия, вызванная продуктами ранних вирусных генов, ведет к усилению деления базальных клеток и задерживает их созревание. Они составляют основную массу папиллом. Поздние вирусные гены кодируют капсидные белки. Вирионы образуются в стадии клеточной дифференциации клеток и, накапливаясь в большом количестве, вызывают цитопатологию.
Вирионы связываются с клеточными рецепторами, проникают в клетки путем эндоцитоза, транспортируются в ядро клетки, где освобождается вирусный геном. При продуктивной инфекции транскрипция генома разделяется на раннюю и позднюю стадии, которые кодируются областями, контролируемыми отдельными промоторами, расположенными на одной из цепей ДНК. Половина генома, содержащая поздние гены, транскрибируется с образованием мРНК, на которых синтезируются ферменты, связанные с репликацией вируса. Поздние мРНК, на которых синтезируются вирионные белки, транскрибируются на другой половине вирусного генома после начала его синтеза. Потомство вирусной ДНК служит дополнительными матрицами, увеличивающими продукцию структурных белков.
Среди папилломавирусов животных наиболее изучен вирус папилломы кроликов. В естественных условиях он вызывает образование доброкачественных опухолей эпителиального происхождения в различных участках тела диких американских и домашних кроликов. Членистоногие — механические переносчики вируса. Экспериментальную инфекцию легко вызывать у домашних кроликов и зайцев. У зараженных животных развиваются сходные опухоли, которые чаще, чем у диких американских кроликов, превращаются в злокачественные. Различные изменения содержат сходные антигены. Антигенное родство между вирусом папилломы кроликов и папилломавирусами крупного рогатого скота, собак, а также человека не установлено. Вирус папилломы кроликов имеет выраженную антигенную активность и индуцирует синтез нейтрализующих и комплементсвязывающих антител. С помощью МАТ в белке вируса папилломы 1 типа крупного рогатого скота выявлены конформационные и линейные эпитопы. Белок L1 является потенциальным иммуногеном, так как содержит иммунодоминантные, нейтрализующие эпитопы. Жизненный цикл вируса совершается в созревающих кератиноцитах, и вирионы освобождаются со слущивающимися клетками. Вероятно, поэтому иммунный ответ на вирусные антигены в инфицированном организме является минимальным. Т-клеточный ответ кажется наиболее важным, тогда как антитела помогают предотвращать распространение инфекции и снижают возможность реинфицирования. Регрессия инфекции и устойчивость к реинфицированию, вероятно, являются типоспецифическим явлением.
Папилломы развиваются после проникновения вируса в кожу. Развивается гиперплазия с последующей дегенерацией и гиперкеритозом. Эти изменения обычно начинаются через 4—6 недель после заражения.
Фибропапилломы, как правило, сохраняются 4—6 месяцев перед спонтанной регрессией; множественные бородавки обычно регрессируют одновременно. Эта стадия характеризуется цитопатологией размножения вируса и образованием кристаллических агрегатов вирионов в местах кожных поражений.
Папилломавирусы КРС 1 и 2 и ДНК, выделенная из них, могут трансформировать клетки in vitro. В отличие от других трансформирующих вирусов, ДНК папилломавирусов остается эписомальной и редко включается в геном клетки. Гены, экспрессируемые в таких клетках, кодируют белки, связанные с репликацией и регуляцией транскрипции вируса, так же как белки, которые непосредственно влияют на трансформацию клеток. Совместно с кофакторами, папилломавирусы могут вызывать карципомы у КРС, кроликов и человека.
Уровень ВН-антител, по-видимому, коррелирует с регрессией кожных изменений и защитой против реинфекции.
Специфическая профилактика папилломавирусных инфекций не разработана. Вакцинация белками вирусного капсида, полученными на основе рекомбинантной ДНК технологии, была обнадеживающей. Трудность состоит в том, что вакцина должна содержать много типов вируса, т.к. перекрестная (межтиповая) защита отсутствует.
Антитела к капсидным вирусным белкам L1 и L2 могут нейтрализовать внеклеточный вирус и защитить чувствительных животных от первичной инфекции. Идеальная вакцина, однако, должна вызывать регрессию патологических изменений, т.е. обладать терапевтическим действием и предохранять от любого проявления инфекции.
Решение задачи специфической профилактики осложняется отсутствием типоспецифических диагностических серологических тестов и невозможностью размножения папилломавирусов в культуре клеток.
ПВ КРС включают различные типы, инфицирующие кожные покровы (BPV1 и BPV2) или слизистые (BPV4). По структуре генома ПВ КРС подобен ПВ человека.
Иммунитет к ПВ КРС может быть вызван введением очищенного вируса или экстракта бородавок КРС. Он коррелирует с высоким титром нейтрализующих антител и является в основном типоспецифическим. Для того чтобы вакцина имела широкий защитный спектр, она должна включать антигены нескольких типов вируса.
Парвовирусы. Строение и виды парвовирусов.
Семейство парвовирусов (parvus - маленький) включает в себя три рода: парвовирусы, депендовирусы и эритровирусы.
К роду парвовирусы относятся: вирус крыс, или вирус Килхема (прототипный вирус), парвовирусы свиней, крупного рогатого скота, лошадей, собак, кроликов, кур, вирусы энтерита норок и гусей, алеутской болезни норок, панлейкопении кошек, мелкий вирус мышей, а также вирусы, происхождение которых не выяснено. В род эритровирусы включен парвовирус человека (В 19), вызывающий апластические кризы при хронической анемии и являющийся этиологическим агентом инфекционной эритемы детей. Возможными представителями рода являются парвовирусы обезьян и бурундуков. Род депендовирусов объединяет аденоассоциированные вирусы (ААВ) человека и обезьян (пять типов), крупного рогатого скота, собак и птиц. Типовой вид — ААВ типа I. Возможными представителями рода являются ААВ лошадей и овец. Депендовирусы, в отличие от других парвовирусов — дефектны, то есть размножаются только в присутствии вирусов-помощников, роль которых выполняют аденовирусы или герпесвирусы.
Недефектные парвовирусы вызывают у животных энтериты, гепатиты, миокардиты, геморрагическую энцефалопатию, панлейкопению, гибель эмбрионов и плодов, отставание в росте, подавляют гемопоэз, лимфопоэз и иммунный ответ. Дефектные парвовирусы не оказывают какого-либо клинически выраженного эффекта.
Вирионы парвовирусов представляют собой безоболочечные изометрические частицы с кубической симметрией диаметром 25 нм. Это довольно мелкие вирусы животных. Капсид парвовирусов состоит из 32 капсомеров диаметром 3—4 нм. Парвовирусы содержат одноцепочечную линейную ДНК размером 5,2 тн (19—32% массы вирионов).
Парвовирусы содержат 63—81% белков. Сравнительным анализом некоторых автономных парвовирусов обнаружено различие в их структурных белках. Парвовирусы собак (ПВС), свиней (ПВСв), кошек (ПВК) и мышей (ПВМ) содержат по три структурных полипептида VP1, VP2, VP3 с молекулярной массой соответственно 79—82; 65—66 и 62—63 кД. Парвовирус крупного рогатого скота содержит четыре полипептида VP1, VP2, VP3 и VP4 с молекулярной массой соответственно 74,5; 67, 62 и 57 кД. Следует отметить, что у парвовируса свиней, выделенного при диарее (штамм Н-45), также обнаружено четыре структурных полипептида (81, 70, 66 и 62 кД). Основным белком у парвовирусов млекопитающих является VP3, на долю которого приходится около 80% массы вирионных белков.

Капсид содержит 60 молекул белка VP2 (65 кД) и небольшое количество молекул VP1 (84 кД). VP1 и VP2 образуются альтернативным сплайсингом одной мРНК и полной последовательности VP2. Третий структурный белок VP3 образуется в полных (ДНК-содержащих) капсидах путем отщепления 15—20 аминокислот от аминоконца VP2. Зависимость размножения парвовируса от стадии клеточного цикла указывает на зависимость его репликации от репликации клеточной ДНК. Вирионы не содержат полимеразы, но способны кодировать такой фермент. Вместо этого используется клеточная полимераза И для транскрипции вирусной ДНК в двухспиральную промежуточную структуру, которая затем используется как матрица для других клеточных ферментов, катализирующих транскрипцию вирусных мРНК. Альтернативный сплайсинг структур приводит к увеличению отдельных видов мРНК, которые транслируются в большое количество различных белков. Наиболее обильный пул представлен мРНК, кодируемыми 3'-половиной генома, управляющей синтезом структурных белков. Неструктурный белок NS1, кодируемый 5'-половиной генома, также образуется в очень большом количестве и выполняет многие функции:
1) он связан с ДНК и требуется для репликации вирусной ДНК;
2) выполняет роль геликазы;
3) служит эндонуклеазой;
4) интерферирует с репликацией клеточной ДНК.
Эта активность ведет к остановке деления клеток в S фазе. Механизм репликации генома является удивительно комплексным. 3'-конец геномной (-) ДНК служит полупраимером для начала синтеза двуцепочечнои циркулярной ДНК в качестве репликативного предшественника.
Одной из особенностей репликации геномной ДНК являлось образование димерной формы репликативного предшественника с последующим синтезом тетрамерной структуры, состоящей из двух полностью позитивных и двух полностью негативных цепей вирусной ДНК, которые с помощью эндонуклеазы превращаются в одноцепочечные молекулы, в том числе в потомство вирусной ДНК.
Парвовирусы размножаются в ядре клеток, находящихся в поздней S фазе или ранней G2 фазе цикла клеточного деления. Такая особенность репликации парвовирусов является основой многих аспектов патогенеза. Это прежде всего поражение тканей в период эмбрионального развития и у молодых животных, а также клеток лимфоидных органов и эпителия кишечника, которые постоянно находятся в стадии деления. Авирулентный штамм вируса алеутской болезни норок легко адаптировался к размножению в клеточной линии FS (клетки селезенки кошки).
Парвовирус собак может размножаться в клетках собак и кошек, но парвови-рус кошек — только в клетках кошек. Парвовирусы кошек и норок не различаются антигенно в РТГА и РН, но отличаются от парвовируса собак. Эти три вируса различаются нуклеиновой последовательностью геномной ДНК.
Антигенная структура парвовирусов недостаточно изучена. Большинство из них обладают ГА-активностью. Общие представления об антигенной структуре разных видов парвовирусов основаны на сравнительном изучении степени их родства в РН, РСК, РТГА методом флуоресцирующих антител и иммунопреципитации. У многих парвовирусов выявлено антигенное сходство вирионных белков вследствие наличия протяженных гомологичных участков в структуре их геномов.
По данным МФА и иммунопреципитации, антигенные связи выявлены у ПВМ, ПВСв, ПВС, ПВК. При анализе в РН антигенное родство было выраженным у ПВС и ПВК, а у остальных парвовирусов менее отчетливым.
Слабое антигенное родство обнаружено у ПВС и ПВСв. Белки ПВКРС (три серотипа) антигенно отличались от белков остальных парвовирусов. Установлено генетическое и биологическое родство между вирусами панлейкопении кошек (ПВК) и вирусом энтерита норок (ВЭН), а также между парвовирусами собак, норок и енотов. Высказано предположение, что ПВС, ВЭН и парвовирус енотов являются хозяинными вариантами вируса панлейкопении кошек.
Антисыворотка к каждому вирионному белку ПВСв нейтрализует инфекционность полных вирионов и взаимодействует со всеми тремя структурными белками данного вируса. Эти результаты свидетельствуют о том, что вируснейтрализующие антигенные детерминанты имеются во всех структурных белках ПВСв. Тесная антигенная взаимосвязь между структурными белками обнаружена также у вируса алеутской болезни норок и, возможно, является характерной особенностью других парвовирусов. Полагают, что капсидные белки парвовирусов обладают не менее чем двумя антигенными участками, представленными несколькими перекрывающимися эпитопами. Однако в опытах с парвовирусом человека В19 отмечена иная ситуация. Продукция ВН-антител наблюдалась лишь при иммунизации животных капсидными препаратами, содержащими одновременно VP1 и VP2. Капсиды, содержащие только VP2, не вызывали образования таких антител, несмотря на то, что уникальный участок VP2 экспонирован на поверхности капсида и содержит эпитопы нейтрализации.
Парвовирус собаки вирус панлейкопении кошек (ПЛК) имели лишь небольшие различия в структуре генома (гомология 99%). Тем не менее вирус ПЛК размножался только в культуре клеток кошек, тогда как парвовирус собак - в большинстве культур клеток собак и кошек. Выявлены специфические для клеток кошек детерминанты, определяющие круг хозяев. Введение двух нуклеотидных остатков парвовируса собак в область гена капсидного белка VP1 вируса ПЛК придало ему способность размножаться в культуре клеток собак.
До недавнего времени было принято считать, что вирус алеутской болезни норок не нейтрализуется специфическими антителами естественного хозяина in vivo и in vitro. Однако после обработки вируса н-бутанолом, фильтрации через мелкопористые мембраны (диаметр пор 0,05 мкм) основная масса его (>95%) быстро нейтрализуется антителами при 37°С, хотя резистентная фракция вируса сохраняет инфекционность. С помощью моноклональных антител эпитопы нейтрализации вируса обнаружены на вирионных белках 75 и 85 кД.
Парвовирусы в естественных условиях поражают животных многих видов и птицу. Парвовирусы, патогенные для одного вида животных, как правило, не вызывают заболевания у животных других видов. Исключение составляют ПВ плотоядных и птиц. Заболевания парвовирусной этиологии широко распространены в мире и наносят существенный ущерб здоровью животных. К таким болезням прежде всего следует отнести парвовирусную инфекцию свиней (ПВИСв), собак (ПВИС), вирусный энтерит гусей (ВЭГ), вирусный энтерит норок (ВЭН), алеутскую болезнь норок (АБН) и панлейкопению кошек. Против большинства из этих болезней разработаны достаточно эффективные средства активной профилактики.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Забота о братьях наших меньших открывает в человеке лучшие качества, воспитывает и укрепляет чувство ответственности, нежность и преданность. К сожалению, иногда радость от этой заботы омрачают неприятности в виде различных заболеваний питомцев, причем в некоторых случаях довольно серьезных и опасных для их жизни. К числу таких болезней относится острое инфекционное заболевание, характеризующееся разнообразными клиническими проявлениями, название которому парвовирусная инфекция.
Чувствительность собак к парвовирусу зависит от их возраста. Наиболее восприимчивы к заболеванию щенки, но могут болеть и взрослые животные. Источником инфекции служат больные собаки, которые выделяют вирус во внешнюю среду с фекалиями, а также вирусоносители. Вместе с тем вирус довольно легко может быть перенесен на подошве обуви хозяина собаки и членов его семьи.
Специалисты выделяют три формы парвовирусной инфекции собак:
— сердечную — у собак в возрасте от 3-8 недель, характеризующуюся острым поражением миокарда;
— комбинированную — преимущественно у собак в возрасте 6-16 недель, выраженную различными поражениями сердечно — сосудистой, пищеварительной и дыхательной систем организма животного.

В связи с угнетением в процессе течения заболевания защитной функции желудочно-кишечного тракта, внутрь организма собаки проникают патологические агенты бактериальной природы, усугубляющие и осложняющие и без того тяжёлый ход болезни. В результате острой сердечнососудистой недостаточности, возникающей при энтерите, ткани и органы собак начинают испытывать недостаток кислорода и питательных веществ, что сказывается и на лёгочной ткани: в ней очень быстро развивается отёк. Прогрессирующим обезвоживанием организма из-за частых рвот и поносов собаки нарушается правильная и упорядоченная работа сердца. По совокупности факторов в результате активизации патогенной микрофлоры, а также быстрого увеличения концентрации вируса, в тканях и органах организма животного накапливаются токсины, вызывающие его общую интоксикацию и лихорадочное состояние со скорым летальным исходом.
В этой связи важно помнить, что ранняя постановка правильного диагноза ровно на половину является гарантией благополучного исхода заболевания.
Предварительно парвовирусную инфекцию диагностируют на основании эпизоотологических данных и таких клинических признаков, как внезапность появления болезни, наличие рвоты, геморрагический энтерит, обезвоживание организма, а иногда лейкопения. Из патологоанатомических изменений чаще всего наблюдают острое катаральное геморрагическое воспаление в тонком отделе кишечника. Окончательный диагноз устанавливают лабораторными методами.
Основным материалом для исследований при прижизненной диагностике парвовирусов являются пробы фекалий, дефибрированная кровь от животных, а также мазки со слизистой прямой кишки. Материал доставляют в лабораторию в день его взятия или на следующий день, сохраняя при комнатной температуре до 30 мин, при температуре от +2 до +8 градусов – до 12ч. Допускается сохранение материала при температуре не выше минус 16 градусов в течение 30 дней. Нужно также учесть следующее: если животное было вакцинировано, то исследования проводятся не ранее, чем через 10 дней после вакцинации.
При посмертной диагностике гистологическому анализу подвергаются незамороженные участки тонкого и толстого отделов кишечника, взятые от павших или вынужденно убитых животных в первые часы после гибели. Срок гистологического исследования — 7 дней.
Читайте также: