Семейство ретровирусов подсемейство лентивирусов
Обновлено: 19.04.2024
Строение ретровирусов. Гликопротеины ретровирусов.
Гликопротеины ретровирусов обладают антигенными детерминантами, которые определяют индивидуальную специфичность каждого вируса. Белки наружной оболочки участвуют в различных взаимодействиях вируса и клетки. С ними связаны такие феномены, как круг хозяев, тканевая специфичность, адсорбция вириона и его проникновение в клетку, специфичность интерференции, индукция синтеза нейтрализующих антител и протективные свойства. Антитела к поверхностным гликопротеинам обладают нейтрализующей активностью. Структурной организацией гликопротеины ретровирусов принципиально не отличаются от гликопропротеинов других оболочечных вирусов. Особенно большое сходство обнаружено в случаях, когда N-концевой домен гликопротеи-на экспонирован на поверхности вириона.
В состав наружного слоя оболочки вирионов онковирусов птиц входят два гликопротеина (gp85, gp37), соединенные друг с другом дисульфидной связью и присутствующие в виде гетеродимера на вирусной оболочке. Образуются они в результате протеолиза белка предшественника 95 env. Круг хозяев вируса саркомы Рауса определяет gp85. В оболочке вирионов онковирусов мышей обнаружен один основной гликозилированный полипептид (gp70) и два минорных полипептида — р15 и р12, образующиеся в результате протеолиза первичного продукта трансляции с молекулярной массой 90 кД.
В структуре вируса лейкоза крупного рогатого скота (ВЛКРС) обнаружено шесть основных белков с молекулярной массой от 10 до 51 кД (р10, р12, р15, р24, gp30 и gp51). Четыре негликозилированных белка (р10, р12, р15 и р24) составляют сердцевину вириона, причем белок р24 присутствует в наибольшем количестве. Два наиболее крупных белка являются гликопротеинами, один из которых (р51) расположен на поверхности вириона, другой (gp30) является трансмембранным. Вирус обладает выраженной антигенной активностью и индуцирует синтез нейтрализующих и комплементсвязывающих антител. Он агглютинирует эритроциты мышей. За инфекционность и гемагглютинирующую активность ответственен gp51. В gp5l выявлено три эпитопа, ответственных за нейтрализацию вируса, что представляет интерес при изготовлении субъединичной вакцины. Наиболее важным, нейтрализующим эпитопы гликопротеина gp51, является участок, соответствующий аминокислотным остаткам 78—92. Поли- и моноклональные антитела против gp51 обладают вируснейтрали-зующей активностью, подавляют синцитий, образующую активность вируса, препятствуют его выходу из клеток и вызывают лизис инфицированных клеток в присутствии комплемента. Антитела к р24 такими свойствами не обладают. Возможно, что фузогенная активность вируса является совместной функцией белков gp51 и трансмембранного gp30.
Ретровирус аденоматоза легких овец (АЛО), в отличие от многих других ретровирусов, трансформирует эпителиальные клетки — секреторные пневмоциты и бронхиолярные эпителиальные клетки. Различают два генотипа вируса: африканский (тип 1) и северо-американский/европейский (тип 2). Ген env вируса АЛО обладает способностью трансформировать фибробласты КЭ и грызунов в культуре клеток. В геноме овцы обнаружены эндогенные ретровирусные последовательности, очень сходные с нуклеотидными последовательностями генома вируса АЛО.
Внеклеточный вирус лентивирусов овец не заражал фиброэндотелиальные клетки овец, которые легко заражались при контакте с инфицированными макрофагами.
В структуре вирусов лейкоза кошек различают белки, связанные с сердцевиной (р10, р12, р15 и р27), и два оболочечные - гликозилированный — 70 кД (gp70) и негликозилированный — 15 кД (р 15). Кроме того, различают вирусспецифические антигены, экспрессируемые на поверхности инфицированных клеток. Гликопротеин gp70 содержит единственный нейтрализующий эпитоп. Нейтрализующие антитела к этому гликопротеину контролируют латентное инфицирование вирусом лейкоза, вызывая его элиминацию из организма кошек. Вирусы лейкоза кошек подгрупп А, В и С различают по структуре главного гликопротеина оболочки gp70.
Лентивирусы относятся к неонкогенным ретровирусам. Они сходны между собой по морфологии вирионов, антигенным свойствам, организации генома, последовательности нуклеотидов, клеточному тропизму и патогенезу. Клетками-мишенями в организме хозяина в основном являются моноциты-макрофаги. Ограниченное размножение вируса происходит даже при отсутствии клинических признаков.
Лентивирусы могут вызывать как литическую, так и персистентную форму инфекции с длительным латентным периодом. Особенностью инфекции является образование ВН-антител с низкой активностью (или они вообще не образуются). Другие вирусспецифические антитела формируются нормально. Для лентивирусов характерна быстрая вариабельность генома. При серийном размножении вируса инфекционной анемии лошадей в культуре макрофагов мутации происходят во многих генах.
Прототипом подсемейства является вирус мэди-висна овец, вызывающий заболевание с инкубационным периодом до одного года. Различают два варианта вируса мэди-висна.
Штаммы мэди характеризуются высоким тропизмом к тканям легких и вызывают пневмонию овец, а штаммы висна с высоким тропизмом к нервной ткани вызывают энцефаломиелит. Однако эти различия не абсолютны. Филогенетический анализ лентивирусов мелких жвачных показал возможность межвидовой трансмиссии этих вирусов. Вирус мэди-висна содержит четыре основных белка: gpl35, р3О, р16 и р14.
В структуре вируса инфекционной анемии лошадей (ИАЛ) обнаружено шесть основных белков с молекулярной массой от 9 до 90 кД и четыре минорных полипептида. В состав наружного слоя оболочки вирионов входят два гликопротеина — gp90 и gp45, не связанные между собой дисульфидными связями. С-концевой участок gp90 и N-концевой участок gp45 вируса ИАЛ являются высококонсервативными и иммунодоминантными областями. Антигенные детерминанты для индукции ВН-антител расположены в гипервариабельной области gp90. Основная иммунная реакция хозяина направлена против высокоантигенного домена, представленного 83 аминокислотными остатками на С-конце главного сердцевинного белка р26.
Поверхностный гликопротеин gp 135 вируса артрита-энцефалита коз формируется в результате посттрансляционной модификации гликозилированного gpl55. Клеточный тропизм вируса артрита-энцефалита не ограничивается лимфоцитами и макрофагами. Установлена частичная гомология генома вируса иммунодефицита человека (ВИЧ) с геномами лентивирусов домашних животных: мэди-висна, артрита-энцефалита коз и ИАЛ. Наиболее выраженная гомология обнаружена с геномом вируса мэди-висна. Вирус иммунодефицитоподобного заболевания крупного рогатого скота генетически и антигенно родственен ВИЧ-1, а вирус иммунодефицита кошек (FIV) вызывает заболевание кошек, напоминающее СПИД.
Для ретровирусов характерна высокая изменчивость генома. Она может быть связана как с иммунологическим давлением на антигенные участки белков, так и с высокой частотой спонтанных мутаций в геноме вируса.
Это касается и лентивирусов, у которых ген, кодирующий нейтрализующие эпитопы поверхностных белков, подвержен частым мутациям. По частоте мутаций лентивирусы превосходят в 100—1000 раз другие РНК-содержащие вирусы, что позволяет им преодолевать иммунную защиту организма. Низкий аффинитет нейтрализующих антител к лентивирусам и персистенция их в макрофагах — главное препятствие в вакцинопрофилактике вызываемых ими заболеваний.
Природа механизма антигенных вариаций при персистентной ретровирусной инфекции хорошо прослеживается на модели вируса ИАЛ. Большое разнообразие антигенных вариантов вирусов ИАЛ может генерироваться в организме инфицированных животных. Существенная изменчивость генов поверхностных гликопротеидов gp90 и gp45 вируса ИАЛ обнаружена в процессе инфекции одного животного. Изоляты, выделенные с интервалом в две недели, антигенно различались между собой, белки сердцевины при этом не изменились. Изменения в гене env в процессе персистирующей инфекции приводят к изменению антигенных свойств белков оболочки вируса ИАЛ и его вариабельности. Антигенный дрейф вируса был связан с потерей двух эпитопов на gp90. Учитывая, что гены поверхностных белков лентивирусов обладают гипервариабельными участками, при создании вакцин против этих вирусов необходимо иметь иммуногены, индуцирующие образование широкого спектра нейтрализующих антител. Большие надежды в этом плане связывают с биотехнологией.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Лентивирусы способны доставлять значительное количество генетического материала в клетку хозяина и обладают уникальной среди ретровирусов способностью реплицироваться в неделящихся клетках, что делает лентивирусы удобным вектором для доставки генетического материала в молекулярной биологии.
Примерами лентивирусов являются: ВИЧ, вирус иммунодефицита обезьян, вирус иммунодефицита кошек.
Содержание
Систематика
Классифицикацию лентивирусов в пять серотипов осуществляют по таксонам позвоночных, которых заражают соответствующие серотипы (приматы, овцы и козлы, лошади, кошки, крупный рогатый скот). Лентивирусы приматов отличаются по рецептору CD4 и по отсутствию фермента dUTPазы. Некоторые группы имеют антигены gag с перекрестной специфичностью.
Морфология
Вирионы имеют оболочку, немного плейоморфные, имеют сферическую форму и диаметр около 80-100 нм. Выступы вирусной оболочки делают поверхность неровной. Нуклеоид концентрический, палочновидный либо имеет вид усеченного конуса.
Структура генома и репликация
Геном вирусов содержит три гена, которые располагаются в геномной РНК в таком порядке 5´-gag-pol-env-3´ . Также геном содержит вспомогательные гены, которые отличаются у разных вирусов (в случае ВИЧ-1 это vif, vpr, vpu, tat, rev, nef). Продукты вспомогательных генов принимают участие в регуляции репликации геномной РНК. Длинные концевые повторы имеют длину около 600 нуклеотидов, участок U3 имеет длину 450, последовательность R — 100 и участок U5 около 70 нуклеотидов.
Лентивирусы способны заражать соседние клетки при непосредственном контакте без образования внеклеточных частиц.
Антигенные свойства
Антигенные детерминанты штаммоспецифичны. Детерминанты, определяющие серотип, находятся на оболочке вируса и являются гликопротеинами. Классификация лентивирусов иногда основывается на антигенных свойствах.
Физико-химические характеристики вирионов
- Общие
- Плавучая плотность 1,16-1,18 г/см −3 в сахарозе
- Вирионы чувствительны к нагреванию, детергентам и формальдегиду
- Инфективность не снижается при радиоактивном облучении
- Вирионы содержат около 2 % нуклеиновых кислот
- Геном состоит из димеров
- Вирионы содержат по одной молекуле линейной (+) одноцепочечной РНК
- Общая длина одного мономера генома составляет 9200 нуклеотидов
- Геном имеет повторяющиеся концевые последовательности; длинные концевые повторы составляют около 600 нуклеотидов
- 5'-конец геном кэпирован, последовательность кэпа — m7G5ppp5’GmpNp
- 3'-конец каждого мономера содержит поли(А); 3'-конец имеет структуру, подобную тРНК и соединяется с лизином
- Внутри капсида содержится исключительно геномная нуклеиновая кислота
- Вирионы содержат 11 разных белков, которые составляют 60 % вирусной частицы
- Пять основных структурных белков по молекулярной массе
- 120 кДа. Gp120 гликозилированный белок оболочки SU, кодируемый вирусным геном env
- 41 кДа. Gp41 гликозилированный трансмембранный белок оболочки TM, кодируемый вирусным геном env
- 24 кДа. P24 негликозилированный белок капсида CA
- 17 кДа. P17 негликозилированный белок ядермного матрикса MA
- 7-11 кДа. Негликозилированный белок капсида NC
- Белки MA, CA и NC закодированы геном gag
- Белок размером 66 кДа. Обратная транскриптаза RT, кодируемая геном pol.
- Белок размером 32 кДа. Интеграза IN, также кодируемая геном pol.
- Белок размером 14 кДа. Протеаза PR, кодируемая геном pro.
- dUPTаза DU, функция которой неизвестна.
Применение
![]()
![]()
Механизм действия и доставка shRNA в клетки млекопитающих при помощи лентивирусных векторов для исследований в области РНК-интерференции
Лентивирусы являются удобными векторами для введения генов в системы in vitro или животные модели. Лентивирусные векторы успешно используются для доставки генно-инженерных конструкций для блокирования экспрессии специфических генов по механизму РНК-интерференции. [2] Экспрессия коротких РНК, содержащих шпильки (shRNA) снижает экспрессию заданного гена и таким образом позволяет судить о функциях данного гена в модельном объекте. Подобные исследования могут предшествовать разработке новых лекарственных препаратов для лечения заболеваний при помощи блокирования экспрессии определенных генов.
Также лентивирусные векторы используют для введения новых генов в клетки человека или животных. Например, в случае модели гемофилии на лабораторных мышах экспрессия тромобоцитарного фактора VIII дикого типа приводит к восстановлению нормального фенотипа. [3] Использование лентивирусных векторов имеет некоторые преимущства перед другими методами терапии генами. Лентивирусы заражают делящиеся и неделящиеся клетки, длительно экспрессируют трансген, и обладают низкой иммуногенностью. Лентивирусы, экспрессирующие PDGF (фактор роста тромбоцитов) успешно используются для трансфекции мышей, страдающих диабетом. [4] Возможно, подобные способы терапии генами будут в дальнейшем будут проводиться и на людях. Векторы на основе гаммаретровирусы и лентивирусов применяются уже более чем в 300 клинических испытаниях, направленных на разработку способов лечения различных заболеваний. [5]
Ретрови́русы (лат. Retroviridae ) — семейство РНК-содержащих вирусов, заражающих преимущественно позвоночных. Наиболее известный и активно изучаемый представитель — вирус иммунодефицита человека.
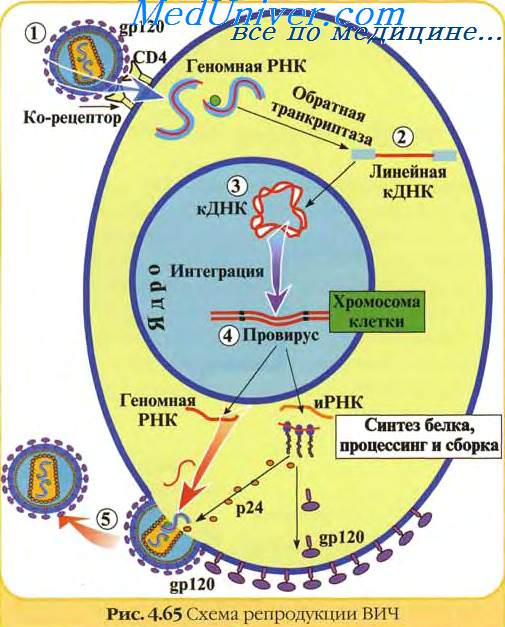
После инфицирования клетки ретровирусом в цитоплазме начинается синтез вирусного ДНК-генома с использованием вирионной РНК в качестве матрицы. Все ретровирусы используют для репликации своего генома механизм обратной транскрипции: вирусный фермент обратная транскриптаза (или ревертаза) синтезирует одну нить ДНК на матрице вирусной РНК, а затем уже на матрице синтезированной нити ДНК достраивает вторую, комплементарную ей нить. Образуется двунитевая молекула ДНК, которая интегрируется в хромосомную ДНК клетки во время клеточного деления, когда нет ядерной оболочки, (исключением является ВИЧ, ДНК которого активно проникает в ядро) и далее служит матрицей для синтеза молекул вирусных РНК. Эти РНК выходят из клеточного ядра и в цитоплазме клетки упаковываются в вирусные частицы, способные инфицировать новые клетки.
По одной из гипотез, ретровирусы могли произойти от ретротранспозонов — подвижных участков генома эукариот. [2]
Содержание
Устройство
Вирионы сферической формы размером 80 — 100 нм, покрыты внешней липопротеиновой оболочкой, имеющей ворсинки длиной 8 — 10 нм. Внутри икосаэдрального капсида находится спиральный РНП. Наружная оболочка, капсидная мембрана и нуклеоид на разрезе вириона расположены концентрически. Чувствительны к эфиру, термолабильны, относительно резистентны к УФ-лучам. Характерной чертой семейства является наличие в составе вириона РНК-зависимой ДНК- полимеразы, иначе называемой обратной транскриптазой. Это и послужило основой для названия семейства (от лат. retro — обратный). Вирионы имеют 6 структурных белков, из них 4 внутренних (капсидных) негликолизированных и 2 гликопротеина оболочки.
Основными структурными генами кодирующими трансляцию белков, из которых в последующем строится вирус, являются gag (group — specificantigens), pol (polymerase), env (envelope). К регуляторным генам относятся: tat (трансактиватор всех вирусных белков), rev (регулятор экспрессии вирионных белков), vif (вирионный инфекционный фактор), vpr (функции остаются неясными), nef (негативный фактор экспрессии), vpx (функции неизвестны)
Капсидные белки несут группоспецифические межвидовые антигены и являются основой для разделения вирусов на роды и подроды. Гликопротеиды являются типоспецифическими антигенами, участвуют в реакции нейтрализации. Геном ретровирусов представлен однонитчатой РНК с молекулярной массой 7 мегадальтон и состоит из двух копий, каждая из которых является полноценным геномом и содержит одинаковую генетическую информацию, однако неизвестно, обе ли они функциональны. Нуклеиновая кислота онковирусов имеет гомологию с клеточной ДНК своего вида хозяина. Вирионная РНК неинфекционна. Вирусная РНК транскрибируется в ковалентно связанную двунитчатую ДНК, которая интегрируется с клеточной ДНК в виде ДНК-провируса. Провирус, экстрагированный из клетки, обладает инфекционностью. Многие вирусы этого семейства вызывают неопластические процессы, главным образом лейкемии и саркомы ряда видов животных. Нормальные клетки некоторых видов животных содержат интегрированные копии соответствующих видов онковирусов. Они могут никак не проявляться или активируются некоторыми физическими и химическими факторами, а возможно, и при инфекции другими онковирусами. Часто встречаются дефектные вирусы, размножающиеся с помощью вируса-помощника. Передаются вертикально и горизонтально. Ретровирусы, наши сожители, враги и помощники [3]
Retroviridae
Семейство Retroviridae включает три подсемейства:
- Oncovirinae (онковирусы), важнейший представитель которого — T-лимфотропный вирус человека типа 1 ;
- Lentivirinae (лентивирусы), к которому относится ВИЧ ; и
- Spumavirinae (спумавирусы, или пенящие вирусы).
Ретровирус раздваивает функции своего генетического материала: инфекционную функцию, то есть функцию самораспространения, выполняет вирусная РНК, а функцию экспрессии вирусных генов и синтеза молекул РНК, которые затем перенесут генетическую информацию в другие клетки, выполняет вирусная ДНК. Попадая внутрь клетки в ходе вирусной инфекции, ретровирусная РНК превращается в ДНК путем хорошо теперь известного процесса обратной транскрипции. Эта ДНК встраивается в геномную ДНК и с этого момента становится неотъемлемой частью генома клетки. А вирус становится провирусом. Провирус для животной клетки это то же самое, что профаг для бактериальной. Кстати, идеи лизогении, по-видимому, и привели Говарда Темина, который вместе с Дэвидом Балтимором открыл обратную транскрипцию, к идее провирусного состояния ретровирусов. По информационному содержанию ДНК-вариант генома ретровируса отличается от РНК-варианта только тем, что ДНК содержит не короткие концевые повторы, а длинные концевые повторы, LTR .
Особенности трансляции РНК ретровирусов
Находясь в составе геномной ДНК, вирусные гены транскрибируются под контролем LTR
LTR, long terminal repeats. Последовательности LTR включают в себя последовательности STR. Возникновение LTR очень важно для экспрессии вирусных генов. Они содержат вирусные регуляторные транскрипционные элементы: промотор, энхансер, и другие. Например, некоторые вирусы содержат элементы, определяющие зависимость вирусной транскрипции от наличия определенных гормонов. LTR и являются теми регуляторными сигналами, которые вирус использует для эксплуатации клеточной транскрипционной машины в своих целях.
Продуктом транскрипции является полноразмерная вирусная РНК. Она должна транслироваться. И здесь вирусу необходимо решить такую проблему: нужно синтезировать много белков, а РНК одна. И в клетках эукариот РНК моноцистронны, то есть предназначены для синтеза только одного белка. Синтез белка в большинстве случаев начинается с ближайшего к кэп-сайту инициирующего кодона [4] .
Если просмотреть открытую рамку считывания от этого ближайшего инициирующего кодона, то мы увидим, что если бы вирус пользовался традиционными способами экспрессии, то он смог бы синтезировать только полипептид GAG. А дальше идет стоп- кодон. Как быть с POL и ENV? Кроме того, эти полипептиды очень длинны, а в вирусе содержатся гораздо более короткие. Проблема решается несколькими способами.
Во-первых, с помощью сплайсинга эта одна РНК превращается в нашем упрощенном варианте еще в одну, более короткую. При этом последовательности, кодирующие ENV полипептид, оказываются рядом с инициирующим кодоном, ближайшим к кэп-сайту, и начинают транслироваться.
Во-вторых, разными для разных ретровирусов способами они ухитряются обойти стоп — кодон после открытой рамки считывания GAG и синтезировать сплавленный полипетид GAG-POL, который содержит последовательности обоих групп белков.
В-третьих, полученные длинные полипептиды подвергаются процессингу и разрезаются на множество белков, которые и функционируют либо в роли регуляторных, как, например, обратная транскриптаза, либо в роли структурных, как, например, белки оболочки зрелых вирусов.
Иными словами, ретровирусы используют гибкую тоталитарную систему для весьма тонкой регуляции синтеза большого разнообразия белков под контролем одного промотора.
Род Deltaretrovirus
Т-лимфотропный вирус человека — это человеческий РНК ретровирус, который вызывает такие злокачественные новообразования лимфоидной и кроветворной тканей, как Т-клеточный лейкоз и Т-клеточную лимфому у взрослых. Возможно вовлечение этого возбудителя в определенные демиелинизирующие болезни, включая тропический спастический парапарез. Взрослый Т-лимфотропный вирус (ВТЛВ) — это штамм вируса-возбудителя указанной болезни, который поражает преимущественно взрослых. Близкородственный вирус — это вирус лейкоза крупного рогатого скота. В настоящее время известно о четырех серотипах этого вида вируса.
См. также
Ссылки
Литература
Примечания
- ↑Таксономия вирусов на сайте Международного комитета по таксономии вирусов (ICTV).
- ↑ А. Марков. Данные сравнительной геномики проливают свет на происхождение ретровирусов.[1]
- ↑ Гловер. Д (ред), 1989 Новое в клонировании ДНК/Под ред. Д.Гловера//М: Мир.- 1989.-367С.
- ↑ Kozak, 1986 (см. в статье консенсусная последовательность Козак)
Классификация вирусов по Балтимору I: дц ДНК-вирусы Caudovirales: Myoviridae • Podoviridae • Siphoviridae
покрытые оболочкой: Herpesviridae • Poxviridae
без облочки: Adenoviridae • Papovaviridae • Papillomaviridae • Polyomaviridae
без группы: Ascoviridae • Asfarviridae • Baculoviridae • Coccolithoviridae • Corticoviridae • Fuselloviridae • Guttaviridae • Iridoviridae • Lipothrixviridae • Nimaviridae • Phycodnaviridae • Plasmaviridae • Rudiviridae • Tectiviridae
MimivirusII: оц ДНК-вирусы без оболочки: Parvoviridae
без группы:Circoviridae • Geminiviridae • Inoviridae • Microviridae • NanoviridaeIII: дц РНК-вирусы Birnaviridae • Chrysoviridae • Cystoviridae • Hypoviridae • Partitiviridae • Reoviridae (Rotavirus) • Totiviridae IV: (+) оц РНК-вирусы Nidovirales: Arteriviridae • Coronaviridae • Roniviridae
Astroviridae • Barnaviridae • Bromoviridae • Caliciviridae • Closteroviridae • Comoviridae • Dicistroviridae • Flaviviridae • Flexiviridae • Leviviridae • Luteoviridae • Marnaviridae • Narnaviridae • Nodaviridae • Picornaviridae (Enterovirus, Rhinovirus) • Potyviridae • Sequiviridae • Tetraviridae • Togaviridae • Tombusviridae • TymoviridaeV: (-) оц РНК-вирусы Mononegavirales: Bornaviridae • Filoviridae • Paramyxoviridae • Rhabdoviridae
Arenaviridae • Bunyaviridae • OrthomyxoviridaeVI: оц РНК-ОТ-вирусы Metaviridae • Pseudoviridae • Retroviridae VII: дц ДНК-ОТ-вирусы Hepadnaviridae • Caulimoviridae - Вирусы по алфавиту
- Ретровирусы
- Семейства вирусов
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Ретровирусы" в других словарях:
РЕТРОВИРУСЫ — РЕТРОВИРУСЫ, представители семейства ретровирусов (Retroviridale), чей генетический код, в отличие от других живых организмов, содержит РНК (рибонуклеиновую кислоту), а не обычную ДНК (дезоксирибонуклеиновую кислоту). Для размножения ретровирусы… … Научно-технический энциклопедический словарь
ретровирусы — семейство РНК–содержащих вирусов. Диаметр вирусных частиц 80 100 нм. Капсид икосаэдрический, заключен в липопротеидную оболочку. Содержит несколько фрагментов одноцепочечной линейной РНК (мол. масса 10–12 млн Да), обратную транскрипта–зу.… … Словарь микробиологии
РЕТРОВИРУСЫ — (от лат. retro назад и вирусы) семейство РНК содержащих вирусов. Обнаружены у всех позвоночных (в т. ч. человека) и некоторых беспозвоночных. Особенность ретровирусов наличие в жизненном цикле обратной транскрипции (синтез ДНК на матрице РНК),… … Большой Энциклопедический словарь
РЕТРОВИРУСЫ — (от лат. retro обратно, назад и вирусы), семейство РНК содержащих вирусов. Диам. вирусных частиц 80 100 нм. Капсид икосаэдрический, заключён в липопротеидную оболочку. Содержат неск. фрагментов одноцепочечной линейной РНК (общая мол. м. 10 12 000 … Биологический энциклопедический словарь
ретровирусы — лейковирусы Группа РНК содержащих вирусов диаметром 70 120 нм, капсид заключен в липопротеиновую оболочку, каждая частица включает по 2 идентичные молекулы РНК и связанные с ними молекулы обратной транскриптазы; многие Р. опухолеродны (вирус… … Справочник технического переводчика
Ретровирусы — * рэтравірусы * retroviruses класс однонитчатых РНКовых вирусов, инфицирующих клетки эукариот, в которых РНК геном транскрибируется в ДНК копию (кДНК) с помощью РНК зависимой ДНК полимеразы (обратной транскриптазы, см.). Эта двунитчатая кДНК… … Генетика. Энциклопедический словарь
РЕТРОВИРУСЫ — вирусы с необычным способом репликации генетического материала. Для цикла репродукции этого большого семейства вирусов характерен обратный поток генетической информации: вместо обычной транскрипции (т.е. переписывания) дезоксирибонуклеиновой… … Энциклопедия Кольера
ретровирусы — (от лат. retro назад и вирусы), семейство РНК содержащих вирусов. Обнаружены у всех позвоночных (в том числе человека) и некоторых беспозвоночных. Особенность ретровирусов наличие в жизненном цикле обратной транскрипции (синтез ДНК на матрице… … Энциклопедический словарь
ретровирусы — (Retraviridae; син. вирусы лейкозов) семейство вирусов, вирионы которых содержат нуклеокапсид и имеют наружную оболочку; геном представлен однонитчатой молекулой рибонуклеиновой кислоты; в вирионе Р. содержится ревертаза (обратная транскриптаза); … Большой медицинский словарь
Лентивирусы (Lentivilinae) - подсемейство вирусов сем. ретровирусов, включающее вирус висны и родственные ему вирусы; Л. относят к медленным вирусам.
Лентивирусы в отличие от онкогенных ретровирусов способны заражать непролиферирующие клетки. Эта способность в значительной степени зависит от белка Vpr , который облегчает перенос провируса в клеточное ядро, вызывает задержку пролиферации клетки в периоде G2 или дифференцировку некоторых клеток- мишеней. Белок Vpx структурно сходен с белком Vpr , но функция белка Vpx окончательно не установлена. Белок Vpu стимулирует разрушение CD4 в эндоплазматическом ретикулуме , а также выход вирусов из клетки. Вирус иммунодефицита человека
Wikimedia Foundation . 2010 .
Смотреть что такое "Лентивирус" в других словарях:
лентивирус — лентивирус … Орфографический словарь-справочник
СИНДРОМ ПРИОБРЕТЕННОГО ИММУННОГО ДЕФИЦИТА — вирусная инфекция человека, сопровождающаяся глубокими нарушениями иммунной системы организма, который оказывается беззащитным перед болезнетворными микробами. Это заболевание обычно приводит к гибели больного от разнообразных инфекций или иных… … Энциклопедия Кольера
СПИД — вирусная инфекция человека, сопровождающаяся глубокими нарушениями иммунной системы организма, который оказывается беззащитным перед болезнетворными микробами. Это заболевание обычно приводит к гибели больного от разнообразных инфекций или иных… … Энциклопедия Кольера
ВИЧ — У этого термина существуют и другие значения, см. Вич. Вирус иммунодефицита человека … Википедия
ВИЧ-инфекция — ? Вирус иммунодефицита человека Стилизованное изображение сечения ВИЧ Научная классификация Надцарство: Вирусы (неклеточ … Википедия
ВИЧ-1 — ? Вирус иммунодефицита человека Стилизованное изображение сечения ВИЧ Научная классификация Надцарство: Вирусы (неклеточ … Википедия
ВИЧ-2 — ? Вирус иммунодефицита человека Стилизованное изображение сечения ВИЧ Научная классификация Надцарство: Вирусы (неклеточ … Википедия
Вирус иммунодефицита человека — ? Вирус иммунодефицита человека Стилизованное изображение сечения ВИЧ Научная классификация Надцарство: Вирусы (неклеточ … Википедия
ЛЖВ — ? Вирус иммунодефицита человека Стилизованное изображение сечения ВИЧ Научная классификация Надцарство: Вирусы (неклеточ … Википедия
Ретровирусы. Семейство ретровирусов. Виды ретровирусов.
Ретровирусы содержат две идентичные молекулы геномной РНК и РНК-зависимую ДНК-полимеразу (обратную транскриптазу, ревертазу). Ретровирусы выделены от множества видов животных и проявляют разнообразный спектр патогенного потенциала. Семейство включает вирусы 7 родов: альфа-, бета-, гамма-, дельта-, эпсилонретровирусы, лентивирусы и спумавирусы. Семейство включает вирусы, патогенные для человека и многих видов животных.
Большинство ретровирусов обладают выраженным тропизмом к клеткам лимфОретикулярной и гемопоэтической системы. Ретровирусы у специфических для них видов животных встречаются повсеместно. В борьбе с ретровирусными инфекциями основной мерой является предотвращение передачи вируса.
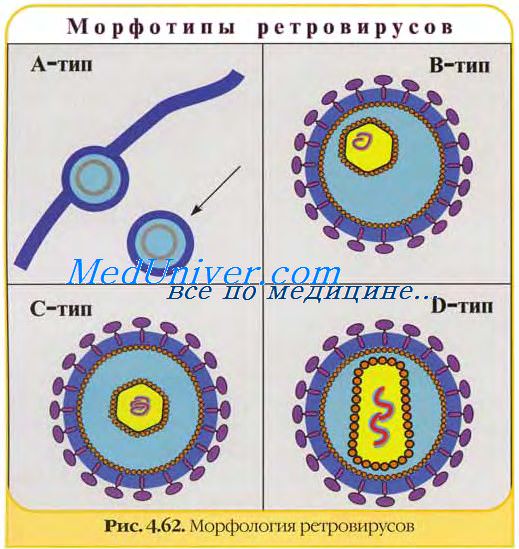
Вирионы ретровирусов представляют собой округлые оболочечные частицы диаметром 80—100 нм, обладающие уникальной трехслойной структурой. Центральная часть вириона представлена нуклеопротеиновым комплексом, который включает около 30 молекул ревертазы и имеет спиральную симметрию. Эта структура окружена икосаэдрическим капсидом диаметром около 60 нм, покрытым оболочкой, происходящей из мембраны клетки, от которой отходят гликопротеиновые пепломеры. Лентивирусы имеют на поверхности примерно 72 шишкоподобных пепломера длиной около 10 нм с яйцевидным концевым уплотнением.
Ретровирусы имеют диплоидный геном, представляющий инвертированный димер из двух молекул линейной позитивной полярности, оцРНК; каждая молекула содержит 7-11 тн и имеет полиА последовательность на З'-конце и КЭП-структуру на 5'-конце. Детальная организация геномов разных ретровирусов широко варьирует.
![ретровирусная инфекция]()
Геном ретровирусов уникален в следующих отношениях:
1) является единственным диплоидным;
2) вирусная РНК синтезируется и изменяется с помощью механизма, изменяющего клеточную мРНК;
3) это единственный геном, связанный со специфическим переносом функции РНК целиком к первичной репликации;
4) это единственная оц(+)РНК, которая не является мРНК вскоре после инфекции;
5) это единственный геном, кодирующий обратную транскриптазу, которая сама по себе уникальна.Среди многих своих функций обратная транскриптаза служит в качестве РНК-зависимой ДНК полимеразы, а ДНК-зависимая ДНК полимераза, интеграза и РНКаза, каждая в меру своей определенной функции, представляют различную часть белковой молекулы. Геном недефектных ретровирусов содержит три разных гена, каждый из которых кодирует два и более белка. Gag ген кодирует вирионные коровью белки, pol ген кодирует обратную транскриптазу, a env ген кодирует вирионные пепломерные белки. Геном кодирует также несколько определенных вирионных компонентов.
Геном быстро трансформирующих ретровирусов содержит четвертый ген — вирусный онкоген (v-onc). Присутствие онкогена обычно связано с делецией в другом месте генома, обычно в env гене, так что большинство v-onc содержащих вирусов не способны синтезировать полностью оболочку и поэтому являются дефектными по репликации. Они всегда находят связь с недефектными вирусами, которые реплицируются полностью и функционируют в качестве помощников. Вирус саркомы Рауса является исключением. Его геном содержит вирусный онкоген v-src и также полные gag, pol и env гены, и поэтому является репликативнокомпетентным.
Vpu, vpr и vpx найдены в лентивирусах приматов. Vpu способствует созреванию вирусных гликопротеинов и освобождению вирионов почкованием; Vpr -слабый активатор транскрипции; Vpx, по-видимому, способствует репликации в Т-лимфоцитах и макрофагах.
При продуктивной инфекции вирионы формируются и освобождаются почкованием через плазматическую мембрану. Некоторые ретровирусы вызывают образование опухолей.
Спумавирусы являются необычными ретровирусами во многих отношениях. Они выделены от человека, обезьян, КРС, кошек, морских львов и хомяков. Они представляют проблему, когда контаминируют клеточные культуры, но их связь с заболеваниями неизвестна. Существует мнение, что пенящие вирусы ближе стоят к ДНК гепаднавирусам.
Эндогенный ретровирус свиней (PERV) найден у всех исследованных пород свиней. Известны три субгруппы этого вируса: А, В и С. Все свиньи содержат множественные провирусы (около 50) в своем геноме. PERVлатентно инфицирует многие культуры клеток свиньи и, реже, человека. Патогенность этого вируса не установлена.
Онковирусы, в отличие от лентивирусов, реплицируются в делящихся клетках. В составе ретровирусов обнаружено 11-13 белков. Часть из них локализована в сердцевине, а главный внутренний белок Р27 - Р30 - в мембране, окружающей РНК. В составе липопротеиновой оболочки 2-5 гликопротеинов, из которых более крупный образует головку шипика и является главным компонентом оболочки, а более мелкие белки образуют отросток, погруженный в билипидный слой. Различные белки ответственны за групповую и типовую специфичность. Подгрупповые и типоспецифические антигены связаны с наружными гликопротеинами и выявляются в РН, РСК, ИФ и РИД. Группоспецифические антигены связаны с белками сердцевины. Их обнаруживают в РСК, ИФ и РИД. Главный внутренний белок Р27 - РЗО - основной группоспецифический антиген, общий для всех ретровирусов птиц или млекопитающих, относящихся к одной группе.
Согласно предложенной ранее номенклатуре структурные белки ретровирусов обозначали следующим образом: МА - матриксный, СА - капсидный, NC -белок нуклеокапсида, SU - поверхностный, ТМ - трансмембранный, PR- протеаза, RT - ревертаза, IN - белок интеграции.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.Читайте также: