
Serotypes of salmonella typhi
Обновлено: 18.04.2024
Salmonella includes a group of gram-negative bacillus bacteria that causes food poisoning and the consequent infection of the intestinal tract. While some of the infections can be easily treated, some of the strains have been shown to resist antibiotic treatment and are therefore deadly. For this reason, infections should not be underestimated.
There are two major species that include:
Classification
The genus Salmonella is closely related to Escherichia coli bacteria and is suggested to have diverged from the bacteria (E. coli) about 150 million years ago. As such, it has adapted and can be found in several niches in the environment.
Several methods of classification of Salmonella have been suggested so far. No single method/approach has been unanimously agreed on.
The following is one of the most recent classifications as used by the Center for Disease Control (CDC) as per recommendations by the World Health Organization (WHO):
- Domain: Bacteria - As bacteria, Salmonella are prokaryotes with a simple cell structure that lacks membrane bound organelles.
- Order: enterobacteriales - Gram-negative rods (Bacillus) that typically move by using flagella and do not form endospores/microcysts
- Family: Enterobacteriaceae - This is the only family in Order enterobacteriales and is composed of rod-shaped gram-negative bacteria.
- Genus: Salmonella
- Species: S. bongori and S. enterica
- Subspecies: S. bongori has a single subspecies referred to as Subspecies V.
The following are subspecies of Salmonella enterica:
- enterica I
- salamae II
- arizonae IIIa
- diarizonae IIIb
- houtenae IV
- indica VI
* In addition to the subspecies, there are also various serotyes of Salmonella that have been suggested to range from 2,200 to about 4,400 serotypes/serovar.
* Serotype grouping is based on cell surface antigens.
Serotypes (Kauffman Classification)
With regards to Salmonella serotypes, the bacteria has been shown to possess three types of antigen. These include antigen H (flagella antigen), antigen O (somatic antigen) and Vi (capsular). These antigens play an important role when it comes to grouping or serotyping the organisms.
-
- This antigen is composed of lipopolysaccharide. Also refered to as somatic antigen, O antigen occurs on the outer membrane and is typically determined by the sugar sequence.
- H antigen - This includes the proteins that are found on the flagella of the bacteria. The H antigen occurs as either phase 1 or phase 2 (or both in some cases). While they may occur in either of these forms, the organisms have also been shown to change from one phase to the other. Currently, studies have identified well over 1800 serovars in this classification.
- Vi antigen - Vi is found in a few serovars and is a superficial antigen that overlies the O antigen. As such, it is an additional antigen found in such organisms as Salmonella typhi and Salmonella paratyphi C where it plays an important role in confirming serotype determination.
* Serotyping is conducted using a pure culture of the organisms isolated on non-selective agar. Some of the media that may be used include: Triple Sugar Iron (TSI), Tryptic Soy Agar (TSA) or Nutrient Agar.
* Agglutination tests involve using polyvalent and monovalent antisera.
Salmonella Microbiology
Evolution and Ecological Niche
According to scientific studies, Salmonella has evolved (from E. coli) for over 150 million years through genetic alterations, which has resulted in changes in pathogen ecology.
By evolving into a complex group composed of well over 2300 genetically/phenotypically diverse serotypes, the bacteria has been shown to be capable of infecting a wide range of hosts (both vertebrates and invertebrates).
Over the course of their evolution, Salmonella has also proven to be capable of adapting to and surviving in different habitats in the environment and are therefore referred to as environmental Salmonellae.
The following are some of the ecological niches in which different species and serovars of Salmonella can be found:
Water - Species like S. enteriditis have been shown to survive in water. While various other serotypes have been shown to be able to live and survive in such water bodies as streams, lakes, and river etc, their survival and lifespan is largely dependent on a variety of factors such as temperature, level of oxygen, contamination (animal faces etc.) as well as competing flora among others.
For instance, for some species, reproduction has been shown to be largely favored by warm temperature of the water as well as contamination by animal feces which provided a source of nutrients. In these environments, aquatic fauna such as frogs can act as reservoirs, playing a secondary role in the spread of the organisms.
Sewage - Serovar like S. paratyphi B have been shown to live and multiply in sewage sludge at about 10 degrees Celsius. In one particular study in Sweden, the bacteria could not be attributed to any animal source and was therefore suggested to live freely in this particular niche.
In the event that such sewage is discharged into other environments such as rivers, sea and soil, it can spread and continue to multiply. This is also one method through which they can ultimately infect animals and human beings.
Birds and wild animals - The levels of Salmonella carried by different birds have been shown to vary. For instance, whereas free-living pigeons carry about 17 percent of Salmonella (S. typhimurium) sport and breeding pigeons have been shown to carry much higher levels of the bacteria while wild ducks were found to carry much lower levels.
While transmission to humans from some birds has been reported, most of the birds, including gulls have largely been shown to play a vector role, transferring the bacteria from one site to the next.
Wild and zoo animals have also proven to be sources of exotic and rare serovars. The carrying rate of these animals is largely dependant on the type of animal and their habitats. In reptiles, diseases are not frequently reported as is the case with some birds and wild animals.
* Agricultural and domestic animals have been shown to contribute in contamination especially through the human food chain. Given their exposure to the bacteria in their environment, poultry have been shown to be a significant source of Salmonella.
Salmonella species can also be found in:
- Animal feeds
- Dairy foods
- Aquatic flora
* Salmonella survive in simple celled organisms like amoeba. In these organisms, the bacteria uses a secretion system to protect itself from enzymes that can degrade it.
Metabolism
Salmonella bacteria are facultative anaerobes that are capable of fermenting glucose, mannitol, and sorbotol.
As such, a majority of Salmonella bacteria have the following characteristics:
- Can grow aerobically or anaerobically - This means that they can also grow in the presence of oxygen. While they are capable of using oxygen for respiration, they can also survive through anaerobic respiration by fermenting organic compounds. Here, however, the fermentative pathway is the final electron acceptor in the process.
- Prefer using oxygen for a greater yield of energy during respiration - As such, a majority of Salmonella thrive when oxygen is present. However, studies have shown that in the presence of readily fermentable substances, a good number of these bacteria will refer fermentation. In this case, the sugars have been shown to repress respiratory enzymes, which in turn favors fermentation while minimizing respiration. In the absence of these sugars, as well as other non-fermentable substances, there is an increase of respiratory enzymes that enhances respiration.
- Compared to anaerobiosis, the breakdown rate of sugar during aerobiosis is smaller.
* While a majority of Salmonella bacteria ferment glucose, mannitol and sorbotol, S. arizonae are capable of fermenting lactose.
* The fermentation of sugars by Salmonella results in the production of acids or gas.
Salmonella are also catalase positive and oxidase negative, characteristics that are also used for the purposes of determining the presence of the bacteria.
Some of the other characteristics used to identify the presence of the bacteria in biochemical tests/reactions include:
- Does not hydrolyze urea
- H 2 S positive
- Reduce nitrate to nitrite
- Lysine-Decarboxylase positive
- Arginine-Dihydrolase variable
- Voges-Proskauer positive
* Catalase is an important enzyme found in Salmonella (as well as all other living organisms that are exposed to oxygen). In these organisms, the enzymes are involved in the decomposition of hydrogen peroxide to produce oxygen and water. This helps protect against the cell oxidative damage.
Salmonella Infection
Naturally, infection is acquired through ingestion of water or foods contaminated with the bacteria. However, it may also be acquired through contact with any of the carriers mentioned above.
With over 2500 serovars of Salmonella identified today, more than 1500 have been shown to belong to subspecies enterica. This group is also composed of the majority of bacteria that infect different types of hosts.
Different types of Salmonella affect different hosts, which has led members of the subspecies to be divided into three major groups based on the type of host they infect (wide host specificity).
Unrestricted serovars - This group includes serovars S. typhimurium and S. enteritidis that infects human beings, poultry, swine, mouse, and cattle.
Infections caused by these organisms:
- Enterocolitis (human beings)
- Septicemia in mice
- Asymptomatic in cattle and poultry
Host adapted - includes serovars S. dublin and S. cholerasuis. Hosts for these bacteria include cattle, pigs, chicken, mouse and rarely infect human beings.
- Enterocolitis in cattle (as well as septicemia)
- Fatal systemic infections in swine
- Bacteremia in human beings as well as in mice
Host restricted - This group includes serovars S. typhi, S. gallinarum, and S. abortusequi. These bacteria are found in horses, human beings and poultry.
Infections in the host include:
* With regards to host specificity, typhi and paratyphi serovars only cause diseases in human beings.
Virulence Factors (Physiology)
Apart from host specificity, several other factors play an important role in the successful infection of the host.
- Endotoxin - Like many other Gram-negative bacteria, some species of Salmonella like Salmonella typhi produce endotoxin (lipopolysaccharides (LPS)), which is a toxic substance that is produced when the outer membrane of the organism is disrupted. This enhances Salmonella infection and inflammation in the affected site.
- Capsule - S. typhi and various strains of S. paratyphi have been shown to possess a capsule as the outermost envelope. These capsules play an important role in the survival of the bacteria given that they cannot be easily removed. As a result, they enhance the infection and have even been associated with resistance to treatment in some cases. Apart from capsules, Salmonella has also been shown to produce outer membrane proteins that allow them to survive in macrophages.
- Adhesives - Apart from capsules that protect the bacteria and enhance their survival, some Salmonella produce such adhesives as fimbrials (and non-fimbrial) that allow the bacteria to remain attached to the surfaces of the infected site. Through reinforced attachment (through these adhesives) bacterial infection is enhanced.
- Flagella - Given that Salmonella tends to affect the intestinal tract, flagella help them move through the intestinal mucus from one site to another.
Human Infections
In human beings and some animals, Salmonella is of clinical importance as they cause salmonellosis and enteric fever. Salmonella infection (salmonellosis) often results in diarrhea, fever (because of inflammation) as well as abdominal cramps that occur 1 to 3 days after infection.
While the infection can last a few days and pass without any treatment, some instances, particularly severe diarrhea, require treatment.
Some infection can spread to the blood stream resulting in toxic shock that can cause death if not treated.
Those at high risk of this infection include:
- Young Children
- Patients with poor/compromised immunity
- Older adults
* Also known as typhoid fever, enteric fever is caused by S. typhi and S. paratyphi and presents the following symptoms:
- General weakness
- Headache
- Cough
- Loss appetite
- Diarrhea and constipation
Pathogenesis
The process through which Salmonella infects and affects human beings in salmonellosis is different from enteric fever.
In salmonellosis, infection starts with ingestion of the bacteria followed by colonization of the lower intestines. This colonization is followed by the invasion of the mucosal layer where acute inflammation takes place.
With regards to enteric fever, oral infection is followed by two weeks of an incubation period after which the bacteria gets to the mucosa layer through the M cells. The infection then proceeds to the local macrophages that transport the bacteria to the mesentarial lymph nodes.
Ultimately, the bacteria can spread to such organs as the lung and liver among others resulting in such complications as empyema, cystitis and typhoid hepatitis.
Prevention and Treatment
For the most part, Salmonella infections are the result of drinking contaminated water, eating uncooked meat, poultry and seafood, uncooked eggs and contaminated fruits and vegetables. It can also be caused through contact with certain pets such as amphibians and reptiles.
Most infections can be prevented by minimizing contact with such pets, particularly for children as well as good hygiene. Here, hygiene involves washing hands with water and soap before eating (and generally keeping them clean), washing all foods before cooking as well as sufficiently cooking meat products that potentially carry the bacteria as well as washing hands after touching pets. These are important prevention tips to avoid infection and the spreading of Salmonella infections.
In some cases, treatment is required. In such cases, treatment involves the use of antibiotics and antimobility drugs as well as replacing fluids and electrolytes. Such drugs as loperamide are used for the purposes of relieving cramping among patients. However, this has been associated with such side effects as diarrhea resulting from the infection.
Microscopy with Gram Stain
A Salmonella sample can be obtained directly from the patient (feces) or from contaminated water/foods. The bacteria may be cultured first using the appropriate agar/media to increase the number of cells.
Sample Preparation
Requirements
- Prepare a smear at the center of the glass slide using a cotton swab stick or wire loop. Make sure that the slide, sample and the cotton swab/wire loop are clean to prevent contamination
- Air dry the slide and heat fix (passing over Bunsen flame approximately 3 times and avoid overheating)
- Place the slide on a staining rack and add a few drops of crystal violet onto the sample, gently wash with water
- Add a few drops of Gram iodine (mordant) for between 30 seconds and 1 minute, gently wash with water
- Add a few drops of alcohol (95% alcohol) for about 10 seconds, gently wash with water
- Add a few drops of safranin (counterstain) and rinse with water
- Use a tissue paper to remove excess water by touching the sides of the slide
- View the slide under the microscope starting with lower power
Observation
When viewed under the microscope, Salmonella bacteria, such as Salmonella newport will appear as pink rods. This indicates that it is a Gram-negative bacteria.
See also Eubacteria page, closely related to E. Coli bacteria and also see Coliform

report this ad
References
C.J. Murray. Salmonellae in the environment. Rev. sci. tech. Off. int. Epiz., 1991, 10 (3), 765-785.
Hiyoshi et al. Typhoidal Salmonella serovars: ecological opportunity and the evolution of a new pathovar. Volume 42, Issue 4, 1 July 2018, Pages 527–541.
Shen, Y. Zhang, Food Microbiology Laboratory for the Food Science Student. Springer International Publishing AG 2017.
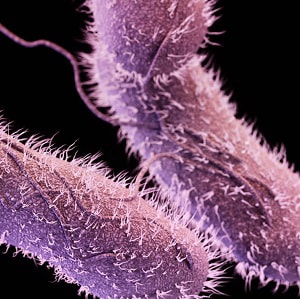
Medical illustration of non-typhoidal Salmonella
Serotypes are groups within a single species of microorganisms, such as bacteria or viruses, which share distinctive surface structures. For instance, Salmonella bacteria look alike under the microscope but can be separated into many serotypes based on two structures on their surface:
- The outermost portion of the bacteria’s surface covering, called the O antigen; and
- A slender threadlike structure, called the H antigen, that is part of the flagella.
The O antigens are distinguished by their different chemical make-up. The H antigens are distinguished by the protein content of the flagella. Each O and H antigen has a unique code number. Scientists determine the serotype based on the distinct combination of O and H antigens.
Different Serotypes
Salmonella have many different serotypes. Some serotypes are only found in one kind of animal or in a single place. Others are found in many different animals and all over the world. Some can cause especially severe illnesses when they infect people; while others cause milder illnesses.
- The bacteria’s surface are covered with lipopolysaccharide (LPS). The outermost portion of the LPS is the O antigen.
- Flagella are whip-like tails that bacteria use to move around. Flagella is the whole structure, while the slender threadlike portion of the flagella is called the H antigen.

New technology is transforming how we detect and investigate outbreaks. Watch the video to learn more.
Some groups of people, such as older adults, people with weakened immune systems, and children under five years old have a higher risk for Salmonella infection. Infections in these groups can be more severe, resulting in long-term health consequences or death. 1
More than 2,500 serotypes have been described for Salmonella; but, because they are rare, scientists know very little about most of them. Less than 100 serotypes account for most human infections. What we learn about the more common serotypes can help us better understand illness and the natural history of all the Salmonella strains.
Serotypes and Outbreaks
Serotyping is a subtyping test based on differences in microbial (e.g., viral or bacterial) surfaces. Serology refers to the antibodies that form because of a viral or bacterial infection. Serotyping is sometimes referred to as serology, but this is technically inaccurate.
Since the 1960s, public health scientists in the US have used serotyping to help find Salmonella outbreaks and track them to their sources. Laboratory experts serotype the Salmonella from infected people. When cases with one serotype increase, they suspect an outbreak and disease detectives start their investigation.
Serotyping has been the core of public health monitoring of Salmonella infections for over 50 years. Now, scientists use DNA testing to further divide each serotype into more subtypes and to detect more outbreaks. With the next generation of sequencing technology, advancements continue as the laboratory can find information about the species, serovar, and subtype of bacteria in just one test. Currently, at least two scientists must generate these three important pieces of information using three separate tests or more. 2
Salmonella and Antibiotic Resistance
Resistance to two clinically important drugs, ceftriaxone (a cephem) and ciprofloxacin (a quinolone), has climbed in non-typhoidal Salmonella since 1996. In 2011, about 5% of Salmonella tested by CDC were resistant to five or more types of drugs. 3
CDC has posted a series of interactive graphs that allow users to see the percentage of Salmonella human isolates resistant to various antibiotics tracked, by year, through the National Antimicrobial Resistance Monitoring System (NARMS). This graph includes the option to view all Salmonella isolates or any one of five common serotypes with resistance to antibiotics used to treat Salmonella infections: Enteritidis, Typhimurium, Newport, Heidelberg, and I 4,[5],12:i: -.
Murielle Baltazar 1 2 , Antoinette Ngandjio 1 , Kathryn Elizabeth Holt, Elodie Lepillet, Maria Pardos de la Gandara 3 , Jean-Marc Collard, Raymond Bercion 4 , Ariane Nzouankeu, Simon Le Hello, Gordon Dougan, Marie-Christine Fonkoua, and François-Xavier Weill
Author affiliations: Institut Pasteur, Paris, France (M. Baltazar, E. Lepillet, M. Pardos de la Gandara, S. Le Hello, F.-X. Weill) ; Centre Pasteur du Cameroun, Yaoundé, Cameroon (A. Ngandjio, A. Nzouankeu, M.-C. Fonkoua) ; University of Melbourne, Melbourne, Victoria, Australia (K.E. Holt) ; Wellcome Trust Sanger Institute, Cambridge, UK (K.E. Holt, G. Dougan) ; Scientific Institute of Public Health, Brussels, Belgium (J.-M. Collard) ; Institut Pasteur de Bangui, Bangui, Central African Republic (R. Bercion)
Abstract
We identified 3 lineages among multidrug-resistant (MDR) Salmonella enterica serotype Typhi isolates in the Gulf of Guinea region in Africa during the 2000s. However, the MDR H58 haplotype, which predominates in southern Asia and Kenya, was not identified. MDR quinolone-susceptible isolates contained a 190-kb incHI1 pST2 plasmid or a 50-kb incN pST3 plasmid.
Typhoid fever, which is caused by Salmonella enterica serotype Typhi, is endemic to the developing world; there were an estimated 26.7 million cases in 2010 (1). The incidence of typhoid fever in sub-Saharan Africa was an estimated 725 cases/100,000 persons in 2010, despite a lack of incidence studies conducted in West and central Africa (1). Antimicrobial susceptibility data are also scarce for this part of Africa. This issue is problematic because treatment with appropriate antimicrobial drugs is essential for recovery in the context of the global emergence of multidrug resistance.
In the Indian subcontinent and Southeast Asia, the multidrug-resistant (MDR) Salmonella Typhi H58 clone, which was named after its haplotype (a combination of defined chromosomal single-nucleotide polymorphisms [SNPs]) (2,3), has spread rapidly and become endemic and predominant. During the 1990s, this clone acquired a large conjugative incHI1 pST6 plasmid encoding resistance to ampicillin, chloramphenicol, and co-trimoxazole (4,5); also in the 1990s, this MDR clone became resistant to quinolones and showed decreased susceptibility to ciprofloxacin because of point mutations in the chromosomal gyrA gene (2). The H58 clone has also spread to eastern Africa, where it has been the most prevalent haplotype (87%) in Kenya since the early 2000s (6).
During 1997–2011, high incidence of MDR Salmonella Typhi was reported in some countries near the Gulf of Guinea in Africa, including Nigeria (7), Ghana (8,9), Togo (10), and the Democratic Republic of the Congo (11). During 1999–2003, a British surveillance system reported a prevalence of 19% (49/421) for MDR Salmonella Typhi isolates among imported cases of typhoid fever acquired in Africa, particularly in Ghana (12). However, nothing is known about the genotypes of these isolates, including whether they belong to the spreading MDR H58 clone.
We report data for the occurrence, genotypes, and characterization of the resistance mechanisms of MDR Salmonella Typhi isolates. These isolates were obtained from the French National Reference Center for Salmonella (FNRC-Salm), Institut Pasteur (Paris, France), and Centre Pasteur du Cameroun (Yaoundé, Cameroon).
The Study
Almost all Salmonella Typhi strains isolated in France are referred to the FNRC-Salm. Most isolates were obtained from travelers or immigrants, most of whom were infected in Africa and Asia. In Cameroon, the Centre Pasteur du Cameroun collects Salmonella Typhi isolates from several hospitals in Yaoundé.
During 1996–2013, a total of 1,746 Salmonella Typhi isolates were collected through the French national surveillance system and subjected to antimicrobial susceptibility testing; 408 were acquired in sub-Saharan Africa (n = 237) and northern Africa (n = 171), and 55 (13.5%) of those acquired in Africa were MDR (Table). All but 1 of the MDR isolates were acquired in sub-Saharan Africa (Table). The proportion of MDR isolates increased from 0% during 1996–1999 to 22.3% (n = 23) during 2010–2013. Only 4 isolates from Africa were resistant to nalidixic acid, including 1 isolate resistant to ciprofloxacin. Because these isolates acquired after 2010 were not MDR isolates, they were not studied further.
In Yaoundé, the proportion of MDR isolates was high (45.5 %, 29/61) in the first survey during 2000–2004. However, this proportion increased to 75.5% (37/49) during 2010–2013.
We studied 61 isolates (Technical Appendix). Of these, 46 were MDR: 29 acquired in Africa and detected at FNRC-Salm before 2010; 2 acquired in France during 2009 at an African festive meal (13); 12 randomly selected acquired in Yaoundé during 2002–2007; 2 acquired in the Central African Republic, and 1 acquired in Morocco (2). The remaining 15 comparison strains (MDR or drug susceptible) that belonged to various haplotypes and were acquired in Africa and Asia during 1958–2009.

Figure 1. Characteristics of 50 Salmonella enterica serotype Typhi isolates. The dendrogram was generated by using BioNumerics version 6.6 software (Applied Maths, Sint-Martens-Latem, Belgium) and shows results of cluster analysis on the basis.
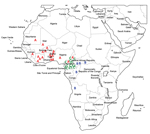
Figure 2. Distribution of multidrug-resistant Salmonella enterica serotype Typhi isolates by genetic lineage (A, B, or C), Gulf of Guinea region, Africa. Location within the country of infection/isolation was assigned at random. UAE.
With the exception of the isolate from Morocco (ISP-03-07467) (2), none of the isolates had the H58 haplotype or contained the associated incHI1 pST6 MDR plasmid. We found 3 other lineages with different geographic distributions and MDR plasmids (Figures 1, 2).
Lineage A consisted mostly of haplotype H56 isolates and more rarely H42 (which differs from H56 by 2 SNPs) and was found only in the western part of the Gulf of Guinea region. Lineage B consisted of haplotype H55 isolates and was found in the eastern and southern parts of the Gulf of Guinea region. Lineage C consisted of haplotype H77 isolates and was found only in Cameroon. All 3 lineages had distinctive CRISPR1 spacer contents. XbaI-PFGE, which used a similarity value of ≥90% as a cutoff, correctly grouped (i.e., concordant with haplotyping and CRISPR results) all but 2 of the MDR isolates from Africa (Figure 1; Technical Appendix).
The 3 lineages contained a large (≈190 kb) conjugative MDR incHI1 pST2 plasmid that differed among lineages. Resistance to trimethoprim was encoded by different class 1 integron gene cassettes: dfrA15, dfrA7, and dfrA1 for incHI1 plasmids of lineages A, B, and C, respectively. All incHI1 plasmids from lineage A encoded resistance to chloramphenicol, and none of those from lineage C encoded such resistance. A second smaller (50-kb) MDR plasmid belonging to the incN incompatibility group (pST3 by plasmid multilocus sequence typing), was present mostly in lineage C isolates, but was also found in 1 lineage A isolate (02-1739) (Technical Appendix).
Conclusions
Analysis of older isolates and previously published data (2) showed that susceptible Salmonella Typhi strains of haplotypes H42, H56, and H77 had already been identified in Senegal in 1962, Tunisia in 1969, and Cameroon in 1958, respectively. This finding suggests that the MDR isolates from lineages A and C are derived from local Salmonella Typhi populations in Africa, rather than being recently imported from other regions to which this bacterium is endemic. Haplotype H55 was previously restricted largely to the Indian subcontinent and eastern Africa (2); it was detected in association with an incHI1 pST2 plasmid in India during the mid-1970s (5). Therefore, lineage B might have been was imported into central Africa from eastern Africa/southern Asia.
A previous study also reported isolation of an MDR clone in the Democratic Republic of the Congo in 2004 that was resistant to quinolones, showed decreased susceptibility to ciprofloxacin, and belonged to the Asian H58 lineage (2). Because only a limited number of isolates from central Africa were tested in our study, studies of a larger collection of isolates might provide more information about bacterial genotypes/MDR plasmids circulating in central Africa.
Despite intrinsic limitations of a laboratory surveillance system for typhoid fever that is used mostly for travelers and immigrants and has the bias of preferential links caused by colonial history and choices of tourist destinations, we documented emergence of 3 MDR Salmonella Typhi lineages in the Gulf of Guinea area. Two lineages found in Guinea and Cameroon were local lineages that acquired MDR conjugative plasmids, either a large incHI1 pST2 plasmid or a smaller incN pST3 plasmid. The H58 lineage, which is currently predominant in Asia and eastern Africa, was not detected among MDR isolates from West and central Africa.
At the time of this study, Ms. Baltazar was a predoctoral student at the FNRC-Salm, Paris, France. She is currently a doctoral candidate at the University of Limoges, Limoges, France. Her primary research interest is mechanisms of antimicrobial drug resistance.
Acknowledgments
We thank all the corresponding laboratories of the FNRC-Salm network for participating in this study.
This study was supported by the Institut Pasteur, the Réseau International des Instituts Pasteur, the Institut de Veille Sanitaire, and the French Government Investissement d'Avenir Program (Integrative Biology of Emerging Infectious Diseases, Laboratory of Excellence, grant ANR-10-LABX-62-IBEID).
References
Buckle GC , Walker CL , Black RE . Typhoid fever and paratyphoid fever: systematic review to estimate global morbidity and mortality for 2010. J Glob Health . 2012 ; 2 : 010401 . DOI PubMed Google Scholar
Roumagnac P , Weill FX , Dolecek C , Baker S , Brisse S , Chinh NT , Evolutionary history of Salmonella Typhi. Science . 2006 ; 314 : 1301 – 4 . DOI PubMed Google Scholar
Holt KE , Parkhill J , Mazzoni CJ , Roumagnac P , Weill FX , Goodhead I , High-throughput sequencing provides insights into genome variation and evolution in Salmonella Typhi. Nat Genet . 2008 ; 40 : 987 – 93 . DOI PubMed Google Scholar
Phan MD , Kidgell C , Nair S , Holt KE , Turner AK , Hinds J , Variation in Salmonella enterica serovar Typhi IncHI1 plasmids during the global spread of resistant typhoid fever. Antimicrob Agents Chemother . 2009 ; 53 : 716 – 27 . DOI PubMed Google Scholar
Holt KE , Phan MD , Baker S , Duy PT , Nga TV , Nair S , Emergence of a globally dominant IncHI1 plasmid type associated with multiple drug resistant typhoid. PLoS Negl Trop Dis . 2011 ; 5 : e1245 . DOI PubMed Google Scholar
Kariuki S , Revathi G , Kiiru J , Mengo DM , Mwituria J , Muyodi J , Typhoid in Kenya is associated with a dominant multidrug-resistant Salmonella enterica serovar Typhi haplotype that is also widespread in Southeast Asia. J Clin Microbiol . 2010 ; 48 : 2171 – 6 . DOI PubMed Google Scholar
Akinyemi KO , Coker AO . Trends of antibiotic resistance in Salmonella enterica serovar typhi isolated from hospitalized patients from 1997 to 2004 in Lagos, Nigeria. Indian J Med Microbiol . 2007 ; 25 : 436 – 7 . DOI PubMed Google Scholar
Mills-Robertson F , Addy ME , Mensah P , Crupper SS . Molecular characterization of antibiotic resistance in clinical Salmonella typhi isolated in Ghana. FEMS Microbiol Lett . 2002 ; 215 : 249 – 53 . DOI PubMed Google Scholar
Gross U , Amuzu SK , de Ciman R , Kassimova I , Gross L , Rabsch W , Bacteremia and antimicrobial drug resistance over time, Ghana. Emerg Infect Dis . 2011 ; 17 : 1879 – 82 . DOI PubMed Google Scholar
Dagnra AY , Akolly K , Gbadoe A , Aho K , David M . Emergence of multidrug resistant Salmonella strains in Lome (Togo) [in French] . Med Mal Infect . 2007 ; 37 : 266 – 9 . DOI PubMed Google Scholar
Lunguya O , Lejon V , Phoba MF , Bertrand S , Vanhoof R , Verhaegen J , . Salmonella Typhi in the Democratic Republic of the Congo: fluoroquinolone decreased susceptibility on the rise. EPLoS Negl Trop Dis. 2012 ; 6:e1921. Doe: . Pub 2012 Nov 5. DOI Google Scholar
Cooke FJ , Day M , Wain J , Ward LR , Threlfall EJ . Cases of typhoid fever imported to England, Scotland and Wales (2000–2003). Trans R Soc Trop Med Hyg . 2007 ; 101 : 398 – 404 . DOI PubMed Google Scholar
Loury P , Tillaut H , Faisant M , Paillereau N , Marquis M , Mari C , Cluster of typhoid fever cases in Ille-et-Vilaine (France), April 2009 [in French] . Bull Epidémiol Hebd. 2010 ; 44 : 446 – 8 .
Fabre L , Le Hello S , Roux C , Issenhuth-Jeanjean S , Weill FX . CRISPR is an optimal target for the design of specific PCR assays for Salmonella enterica serotypes Typhi and Paratyphi A. PLoS Negl Trop Dis . 2014 ; 8 : e2671 . DOI PubMed Google Scholar
Figures
Table
1 These authors contributed equally to this article.
2 Current affiliation: University of Limoges, Limoges, France.
3 Current affiliation: Rockefeller University, New York, New York, USA.
4 Current affiliation: Institute Pasteur de Dakar, Dakar, Senegal.
Please use the form below to submit correspondence to the authors or contact them at the following address:
François-Xavier Weill, Centre National de Référence des Escherichia coli, Shigella et Salmonella, Unité des Bactéries Pathogènes Entériques, Institut Pasteur, 28 Rue du Docteur Roux, 75724 Paris CEDEX 15, France
Comment submitted successfully, thank you for your feedback.
There was an unexpected error. Message not sent.
The conclusions, findings, and opinions expressed by authors contributing to this journal do not necessarily reflect the official position of the U.S. Department of Health and Human Services, the Public Health Service, the Centers for Disease Control and Prevention, or the authors' affiliated institutions. Use of trade names is for identification only and does not imply endorsement by any of the groups named above.
Marie Accou-Demartin, Valérie Gaborieau, Yajun Song, Philippe Roumagnac, Bruno Marchou, Mark Achtman, and François-Xavier Weill
Author affiliations: Author affiliations: Institut Pasteur, Paris, France (M. Accou-Demartin, F.-X. Weill) ; Hôpital Purpan, Toulouse, France (V. Gaborieau, B. Marchou) ; University College Cork, Cork, Ireland (Y. Song, M. Achtman) ; Centre de Coopération International en Recherche Agronomique pour le Développement, Montpellier, France (P. Roumagnac)
Abstract
We report Salmonella enterica serotype Typhi strains with a nonclassical quinolone resistance phenotype (i.e., decreased susceptibility to ciprofloxacin but with susceptibility to nalidixic acid) associated with a nonsynonymous mutation at codon 464 of the gyrB gene. These strains, not detected by the nalidixic acid disk screening test, can result in fluoroquinolone treatment failure.
The Study
Figure 1. Scattergrams for 685 Salmonella enterica serotype Typhi isolates correlating the zone diameters around the 5-μg ciprofloxacin disk with those of the 30-μg nalidixic acid disk. Circle sizes are proportional to the.
In France, laboratory surveillance of typhoid fever infections is performed by the FNRC-Salm through its network of ≈1,500 hospital and private clinical laboratories. Almost all Salmonella Typhi isolates in France are referred to the FNRC-Salm, and almost all are acquired abroad, mainly in Africa and Asia. Until 2009, Cip DS Salmonella Typhi was monitored with the 30-μg Nal screening test. A total of 685 Salmonella Typhi isolates collected during 1997–2009 were reanalyzed to identify Nal S –Cip DS Salmonella Typhi isolates. The scattergram correlating the zone diameters around the 5-μg ciprofloxacin disk with those of the 30-μg Nal disk showed 4 subpopulations, which were labeled A (554 isolates), B (11 isolates), C (119 isolates), and D (1 isolate) (Figure 1). The characteristics of these populations are shown in Table 1 and Table 2. The QRDRs of gyrA, gyrB, parC, and parE genes were studied on 133 isolates selected to represent diversity in terms of year of isolation, geographic origin, and MICs. To analyze the isolate characteristics, we used the following approaches: sequencing (5), denaturing high performance liquid chromatography (4), and Luminex-based genotyping assays (12). QRDR DNA sequences were compared with those of Salmonella Typhi strain Ty2 (GenBank accession no. AE014613). In subpopulation A, 75 isolates had wild-type QRDR sequences, whereas 2 isolates had a gyrB mutation at codon 465 leading to amino acid substitution Gln to Leu. Their Nal MICs were 2 and 4 μg/mL, respectively, and those of Cip were 0.04 μg/mL and 0.08 μg/mL, respectively. Notably, both isolates were acquired in Mexico during 1998 and 2009, respectively. In subpopulation C, the lowest MIC values for Cip (0.06 μg/mL) were associated with a mutation at codon 87 of the gyrA gene, whereas MICs did not increase with the additional mutation in the parE gene. Subpopulation D consisted of 1 isolate, highly resistant to ciprofloxacin, which was acquired by a traveler in India in 2004. This isolate contained 2 NS mutations in the gyrA gene and 1 in the parC gene.
Figure 2. XbaI pulsed-field gel electrophoresis (PFGE) profiles obtained from 10 Salmonella enterica serotype Typhi isolates belonging to subpopulation B. The dendrograms generated by BioNumerics version 3.5 software (Applied Maths, Sint-Martens-Latem, Belgium) show.
Conclusions
Nal S –Cip DS Salmonella Typhi isolates originating from Asia comprise ≈1% of Salmonella Typhi isolates in France but are more prevalent in the United States and the United Kingdom. The NS gyrB mutation at codon 464 was found exclusively in Nal S –Cip DS isolates; however, the effects of this mutation need to be formally demonstrated by site-directed mutagenesis. Furthermore, the involvement of an efflux system, such as AcrAB-TolC and OqxA, or the qnrC gene, have not been investigated and cannot be excluded.
Whatever the molecular mechanism of resistance of such strains, the main concern is detection of such isolates in clinical practice to prevent fluoroquinolone treatment failures. Consequently, the Nal R screening test should no longer be recommended and ciprofloxacin drug MICs should be determined for all Salmonella Typhi isolates instead. There is also a clear need to reevaluate the clinical breakpoints for this pathogen.
Acknowledgments
We thank all the corresponding laboratories of the French National Reference Center for Salmonella network for their assistance.
This work was funded by the Institut Pasteur (M.A.-D., F.-X.W.), the Institut de Veille Sanitaire (M.A.-D., F.-X.W.), and the Science Foundation of Ireland (grant no. 05/FE1/B882 to Y.S. and M.A.).
References
Crump JA , Mintz ED . Global trends in typhoid and paratyphoid fever. Clin Infect Dis . 2010 ; 50 : 241 – 6 . DOI PubMed Google Scholar
Wain J , Kidgell C . The emergence of multidrug resistance to antimicrobial agents for the treatment of typhoid fever. Trans R Soc Trop Med Hyg . 2004 ; 98 : 423 – 30 . DOI PubMed Google Scholar
Wain J , Hoa NT , Chinh NR , Vinh H , Everett MJ , Diep TS , Quinolone-resistant Salmonella typhi in Vietnam: molecular basis of resistance and clinical response to treatment. Clin Infect Dis . 1997 ; 25 : 1404 – 10 . DOI PubMed Google Scholar
Roumagnac P , Weill FX , Dolecek C , Baker S , Brisse S , Chinh NT , Evolutionary history of Salmonella Typhi. Science . 2006 ; 314 : 1301 – 4 . DOI PubMed Google Scholar
Le TA , Fabre L , Roumagnac P , Grimont PA , Scavizzi MR , Weill FX . Clonal expansion and microevolution of quinolone-resistant Salmonella enterica serotype Typhi in Vietnam from 1996 to 2004. J Clin Microbiol . 2007 ; 45 : 3485 – 92 . DOI PubMed Google Scholar
Aarestrup FM , Wiuff C , Mølbak K , Threlfall EJ . Is it time to change fluoroquinolone breakpoints for Salmonella spp.? Antimicrob Agents Chemother . 2003 ; 47 : 827 – 9 . DOI PubMed Google Scholar
Crump JA , Kretsinger K , Gay K , Hoekstra RM , Vugia DJ , Hurd S , Clinical response and outcome of infection with Salmonella enterica serotype Typhi with decreased susceptibility to fluoroquinolones: a United States FoodNet Multicenter Retrospective Cohort Study. Antimicrob Agents Chemother . 2008 ; 52 : 1278 – 84 . DOI PubMed Google Scholar
Crump JA , Barrett TJ , Nelson JT , Angulo FJ . Reevaluating fluoroquinolone breakpoints for Salmonella enterica serotype Typhi and for non-Typhi salmonellae. Clin Infect Dis . 2003 ; 37 : 75 – 81 . DOI PubMed Google Scholar
Booker BM , Smith PF , Forrest A , Bullock J , Kelchlin P , Bhavnani SM , Application of an in vitro infection model and simulation for reevaluation of fluoroquinolone breakpoints for Salmonella enterica serotype Typhi. Antimicrob Agents Chemother . 2005 ; 49 : 1775 – 81 . DOI PubMed Google Scholar
Cooke FJ , Day M , Wain J , Ward LR , Threlfall EJ . Cases of typhoid fever imported to England, Scotland and Wales (2000–2003). Trans R Soc Trop Med Hyg . 2007 ; 101 : 398 – 404 . DOI PubMed Google Scholar
Lynch MF , Blanton EM , Bulens S , Polyak C , Vojdani J , Stevenson J , Typhoid fever in the United States, 1999–2006. JAMA . 2009 ; 302 : 859 – 65 . DOI PubMed Google Scholar
Song Y , Roumagnac P , Weill FX , Wain J , Dolecek C , Mazzoni CJ , A multiplex single nucleotide polymorphism typing assay for detecting mutations that result in decreased fluoroquinolone susceptibility in Salmonella enterica serovars Typhi and Paratyphi A. J Antimicrob Chemother. 2010 Jun 1. PMID: 20511368
Gaborieau V , Weill FX , Marchou B . Salmonella enterica serovar Typhi with decreased susceptibility to ciprofloxacin: a case report [in French] . Med Mal Infect . 2010 ; 40 : 691 – 5 . DOI PubMed Google Scholar
Sjölund-Karlsson M , Howie R , Rickert R , Krueger A , Tran TT , Zhao S , Plasmid-mediated quinolone resistance among non-Typhi Salmonella enterica isolates, USA. Emerg Infect Dis . 2010 ; 16 : 1789 – 91 . PubMed Google Scholar
Читайте также:
- Циклоферон для профилактики клещевого энцефалита
- Ветеринарно санитарная экспертиза при актиномикозе
- Мероприятия по удалению патогенных микроорганизмов в присутствии источника инфекционного заболевания
- Чем лечить кишечную палочку гемолитическую у взрослого
- Кишечная палочка стафилококк клебсиелла стрептококк