
Скронева зона кори головного мозку
Обновлено: 15.04.2024
- у лобовій частці(lobus frontalis):
- передцентральну борозн (sulcus precentralis);
- верхню лобову борозна (sulcus frontalis superior);
- нижню лобову борозна (sulcus frontalis inferior);
- передцентральну звивину (gyrus precentralis);
- верхню лобову звивину (gyrus frontalis superior);
- середню лобову звивину (gyrus frontalis medius);
- нижню лобову звивину (gyrus frontalis inferior), яка за допомогою передньої гілки та висхідної гілки бічної борозни (ramus anterior et ramus ascendens sulci lateralis) поділяється на:
- покришкову частину (pars opercularis);
- трикутну частину (pars triangularis);
- очноямкову частину (pars orbitalis).
- у тім’яній частці(lobus parietalis):
- зацентральну борозну (sulcus postcentralis);
- внутрішньотім’яну борозну (sulcus intraparietalis);
- зацентральну звивину (gyrus postcentralis);
- верхню тім’яну часточку (lobulus parietalis superior);
- нижню тім’яну часточку (lobulus parietalis inferior), в якій знаходяться:
- надкрайова звивина (gyrus supramarginalis);
- кутова звивина (gyrus angularis);
- у скроневій частці(lobus temporalis):
- верхню скроневу борозну (sulcus temporalis superior);
- нижню скроневу борозну (sulcus temporalis inferior);
- поперечну скроневу борозну (sulcus temporalis transversus);
- верхню скроневу звивину (gyrus temporalis superior);
- середню скроневу звивину (gyrus temporalis medius);
- нижню скроневу звивину (gyrus temporalis inferior);
- поперечні скроневі звивини (gyri temporales transversi) – звивини Гешля, які містяться на присередній поверхні верхньої скроневої звивини (facies medialis gyri temporalis superioris).
употиличній частці(lobus occipitalis):
- поперечну потиличну борозну(sulcus occipitalis transversus), яка обмежовує досить варіабельні потиличні звивини (gyri occipitales);
- півмісяцеву борозну(sulcus lunatus);
- передпотиличу вирізку (incisura preoccipitalis);
- у острівцевій частці; острівцi (lobus insularis; insula):
- центральна борозна острівця (sulcus centralis insulae);
- колову борозну острівця (sulcus circularis insulae);
- звивини острівця (gyri insulae);
- довгу звивину острівця (gyrus longus insulae);
- короткі звивини острівця (gyri breves insulae);
Строение коры головного мозга: слои, клетки
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
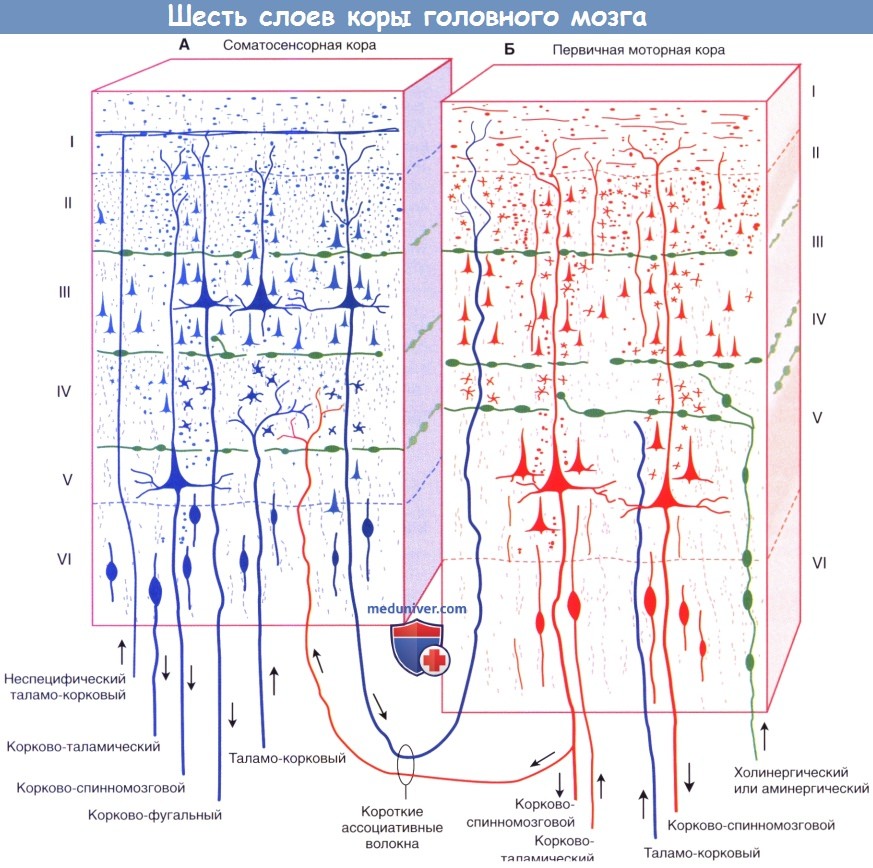
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
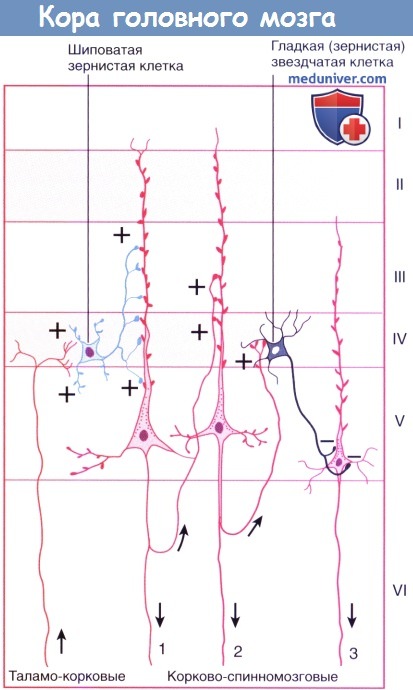
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
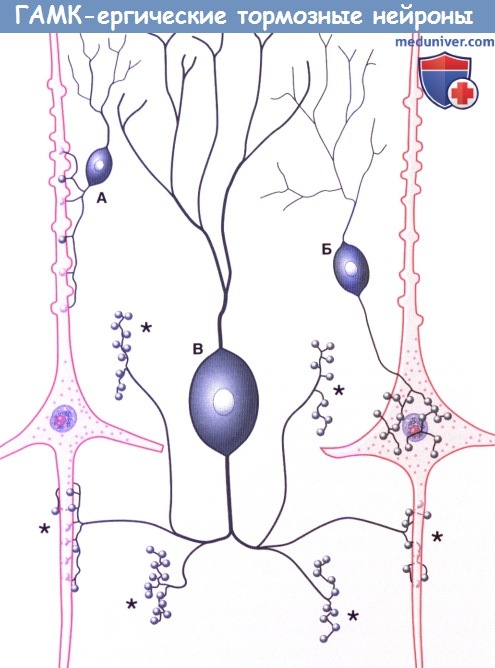
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Сенсорные зоны коры головного мозга: соматосенсорная кора, теменная долька, зрительная кора, слуховая кора
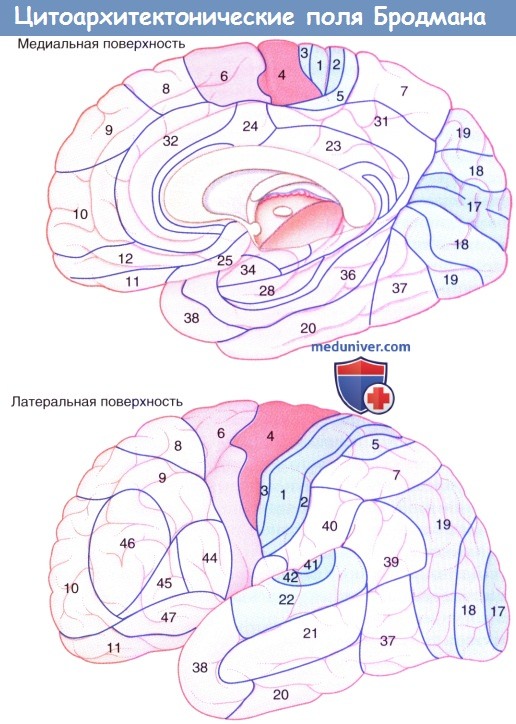
Цитоархитектонические поля Бродмана. Окрашенные зоны:
а) Моторная (красная):
4 — первичная моторная кора
6 — на медиальной поверхности, дополнительная моторная зона
6 — на латеральной поверхности, премоторная кора
б) Сенсорная (голубая):
3/1/2 — первичная соматосенсорная кора
40 — вторичная соматосенсорная кора
17 — первичная зрительная кора 18,
19 — ассоциативная зрительная кора
41, 42 — первичная слуховая кора*
22 — ассоциативная слуховая кора
(*Первичную слуховую кору не всегда можно увидеть сбоку, так как она полностью расположена на верхней поверхности верхней височной извилины.)
а) Соматосенсорная кора:
1. Отделы. Соматосенсорная, или соматоэстетическая, кора занимает всю постцентральную извилину. Части тела противоположной стороны представлены инвертированными (за исключением лица), а кисти рук, губы и язык представлены непропорционально большими. Представленные на рисунке ниже уже известные Вам гомункулы лишь схематически изображают расположение различных частей тела без учета обширных участков, перекрывающих друг друга.
2. Афферентные волокна. Помимо таламических афферентных волокон от вентрального заднего ядра, к соматосенсорной коре подходят также комиссуральные волокна от противоположной соматосенсорной коры через мозолистое тело и короткие ассоциативные волокна от прилежащей первичной моторной коры. Множество волокон от моторной коры служат коллатералями корково-спинномозговых волокон, идущих к переднему рогу спинного мозга, и участвуют в создании чувства веса при подъеме объектов (потерю этого чувства называют барагнозией).
Поражение соматосенсорной коры часто возникает при окклюзии ветви средней мозговой артерии, кровоснабжающей сенсорную кору. Корковый тип потери чувствительности в этом случае проявляется ослаблением чувствительности на противоположной стороне тела, особенно на предплечье и кисти (проявляющимся повышением порога возбудимости, ослаблением дискриминационного чувства, нарушением вибрационного чувства и чувства положения), а также нарушением распознавания более сложных видов чувствительности, несмотря на сохранность болевой, температурной и даже вибрационной чувствительности. Поражение может проявляться невозможностью распознавания знакомых объектов при помещении их в руку (астереогноз), невозможностью распознавать фигуры и цифры, рисуемые на руке (аграфестезия), или неспособностью опознать одновременно два тактильных стимула, приложенных к противоположным частям тела (экстинкция). Потерю способности распознавать размер и форму объектов в результате поражения на уровне между чувствительными рецепторами и корой обозначают термином стереоанестезия.
Сложные неврологические нарушения при повреждении теменной доли, обычно недоминантного полушария (чаще правого), обозначают общим термином агнозии (тактильная агнозия — неспособность распознать форму объекта при его ощупывании, анозогнозия—отрицание болезни или неврологических нарушений, аутопагнозия—неспособность к определению, ориентированию и распознаванию собственных частей тела). Повреждение теменной доли (чаще левой) может, кроме того, приводить к апраксии—невозможности совершения целенаправленных движений или правильного использования объекта, несмотря на сохранную моторику и осознанное восприятие.
3. Эфферентные волокна. К эфферентным волокнам соматосенсорной коры относят ассоциативные, комиссуральные и проекционные волокна. Ассоциативные волокна направляются к моторной коре на той же стороне к полю 5 и полю 40 (надкраевой извилине). Комиссуральные волокна проходят к соматосенсорной коре противоположных полушарий. Проекционные волокна опускаются в заднем отделе пирамидного пути и оканчиваются на вставочных нейронах сенсорных релейных ядер (вентральное заднее ядро таламуса той же стороны), а также проходят в заднем канатике к заднему рогу спинного мозга на противоположной стороне. Передача чувствительной информации по спинно-таламическому проводящему пути может ингибироваться (тормозными вставочными нейронами) при интенсивных физических нагрузках, тогда как проведение через задний канатик (в составе медиального лемнискового пути) может усиливаться (возбуждающими вставочными нейронами) при исследовательской активности, например при пальпации рельефной поверхности.
б) Ассоциативная соматосенсорная кора (поле 5). Этим термином обозначают поле 5, расположенное сразу за соматосенсорной корой. Активация большей части этой области происходит при совершении хватательных движений противоположной кистью под контролем зрения (дорсальный зрительный путь описан ниже).
г) Нижняя теменная долька (поля 39 и 40). Нижняя теменная долька образована полями 39 (угловая извилина) и 40 (надкраевая извилина). Оба поля отвечают за речь — функцию преимущественно левого полушария, описанную в отдельной статье на сайте; нарушение речи, вызванное поражением мозга, носит название афазия. (Поражение правого полушария может приводить к развитию неспособности понимания или использования эмоций при устной речи — апрозодии.)
д) Межтеменная кора. Повышенная активация коры в стенках межтеменной борозды возникает при выполнении задач, требующих зрительно-двигательной координации (например, при приближении и захвате объектов, идентифицированных полем зрения противоположной стороны, с последующим зрительным и тактильным пространственным анализом). К этой зоне относят также теменное глазодвигательное поле.
е) Вторичная соматосенсорная кора. На медиальной поверхности теменной покрышки островка расположена небольшая вторичная соматосенсорная кора (SII). Она получает ноцицептивные пути от таламуса и ярко светится на ПЭТ-снимках головного мозга при периферической болевой стимуляции. SII, вероятно, действует совместно с SI при обработке дискриминационной чувствительности или локализации болезненного участка.
• Срединный нерв иннервирует ладонную поверхность латеральных трех с половиной пальцев кисти, а лучевой нерв иннервирует их тыльные поверхности. При повреждении срединного нерва увеличивается зона представления в коре тыльной поверхности кисти, захватывая зону ладонной чувствительности. Расширение зоны начинается через несколько часов и прогрессивно увеличивается в течение нескольких недель. При восстановлении функции срединного нерва, кортикальная карта возвращается к норме.
• При денервации среднего пальца соответствующая корковая зона в течение нескольких часов не отвечает на стимуляцию, а затем прогрессивно (в течение нескольких недель) замещается зонами II и IV пальцев.
• При постоянной стимуляции кожи подушечки пальца (например, при соприкосновении с вращающимся диском для приготовления гранул пищи) представление этого пальца в коре может увеличиваться в два раза в течение нескольких недель, возвращаясь к норме после прекращения эксперимента.
Эти исследования показывают, что соматосенсорные карты обладают пластичностью и изменяются при нарушении периферической иннервации. Чисто анатомический подход (например, разрастание нервных волокон в центральной или периферической нервной системе) не позволяет объяснить ранние изменения, развивающиеся в течение нескольких часов: наоборот, их следует рассматривать как основу сенсорной конкуренции.
ж) Сенсорная конкуренция. В сенсорных картах, созданных на уровне задних рогов, задних канатиков, таламуса и соматосенсорной коры, выявлены признаки наложения (анатомического перекрытия). Например, таламо-корковые соматосенсорные зоны III пальца накладываются на зоны II и IV пальцев. В зоне наложения корковые колонны получают информацию от двух соседних пальцев. Вставочные корковые нейроны могут тормозить слабо возбужденные периферические колонны. В эксперименте (например, на кошках) число колонн, отвечающих на направленную таламо-корковую стимуляцию, может быть увеличено местным воздействием антагонистов ГАМК (бикукуллин), подавляющих периферическое торможение. Результат отсутствия периферического сенсорного поля может быть различным.
з) Зрительная кора (поля 17, 18, 19). К зрительной коре относят первичную зрительную кору (поле 17) и ассоциативную зрительную кору.
1. Первичная зрительная кора (поле 17). Первичная зрительная кора — место окончания коленчато-шпорного тракта, через который передается информация от ипсилатеральных половин обеих сетчаток и, следовательно, от зрительного поля противоположного глаза. Этот миелинизированный путь образует в первичной зрительной коре бледные зрительные полоски (линии Геннари) перед созданием контактов с шиповатыми зернистыми клетками в высокозернистом слое IV. Зрительные полоски (впервые обнаруженные студентом-медиком Франческо Дженнари в 1775 г.) дали альтернативное название для поля 17 — полосатая кора.
Шиповатые зернистые клетки находятся в глазных доминантных колоннах, названных так, поскольку к чередующимся колоннам попеременно подходят импульсы от левого и правого глаз. Если можно было бы отдельно промаркировать волокна от каждого глаза и посмотреть на зрительную кору снаружи, эти чередующиеся колонны образовывали бы пучки в форме витков (наподобие отпечатков пальцев), где каждый из пучков отвечал бы на стимуляцию только одного из глаз. Коленчато-шпорный тракт представлен в коре таким образом, что соответствующие точки от двух сетчаток расположены в соседних колоннах на одной линии. Такое расположение идеально подходит для бинокулярного зрения, так как серия этих колонн образует модули, в краях которых происходит обработка информации от обоих глаз.
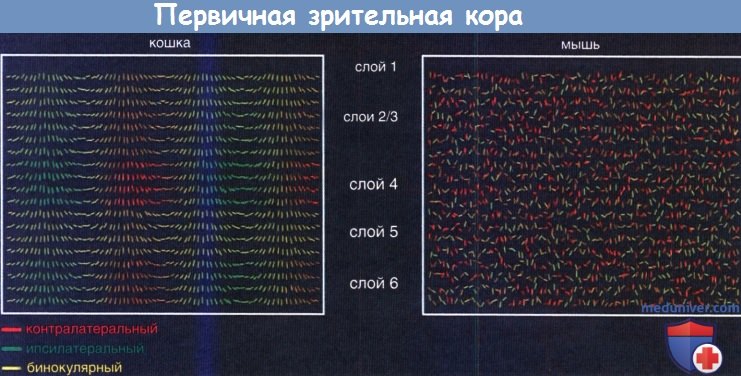
Представление избирательного действия нейронов первичной зрительной коры (V1), получающей информацию от латерального коленчатого тела.
Запись нейрональной активности взрослой кошки (слева) показывает высокую избирательность для специфических ориентаций стимулов (показаны наклоном линий) и различную степень доминирования противоположного (красный) и ипсилатерального (зеленый) глаз с множеством клеток, регулирующих деятельность обоих глаз.
Как ориентация, так и глазодоминирование (нейроны, в большей степени отвечающие на стимуляцию от одного глаза, чем от другого) распределены по колоннам. В колоннах, реагирующих на определенную ориентацию стимула, участвуют все клеточные слои, тогда как глазодоминирование происходит преимущественно в слое 4, где большое число клеток отвечает за монокулярное зрение. Поле V1 у мыши (справа) не имеет колончатого строения для ориентации или глазодоминирования.
Однако все же имеется высокая избирательность нейронов к ориентации, а также в некоторой степени отмечено глазодоминирование, но со смещением к противоположному глазу.
2. Пластичность первичной зрительной коры. Базовая схема и соотношение глазных доминантных колонок закладывается и устанавливается еще до рождения и сохраняется у животных, обитающих в полной темноте. При развитии в детстве слепоты на один глаз соответствующие корковые колонны остаются небольшими, а размеры корковых колонн видящего глаза увеличиваются.
3. Ассоциативная зрительная кора (поля 18 и 19). Ассоциативная зрительная кора представлена зонами 18 и 19, имеющими также общее название перистриарная, или экстрастриарная, кора. К ней подходят афферентные волокна преимущественно от поля 17, а также небольшое количество прямых таламических волокон от подушки таламуса. Клеточные колонны отвечают за обработку отдельных признаков. Часть колонн обрабатывает геометрические формы, часть отвечает за обработку цвета, другая часть — за бинокулярное (пространственное) зрение, а некоторые колонны — за более сложные задачи (распознавание лиц).
Активация клеточных колонн поля 7 у обезьян происходит, когда важный объект (например, фрукт) появляется в противоположной половине зрительного поля. Через ассоциативные волокна возбужденные клеточные колонны повышают частоту фоновой активности колонн во фронтальном глазодвигательном поле и премоторной коре, но без инициации движения. Этот эффект называют скрытым вниманием, или скрытой ориентацией. Внимание (ориентация) становится открытым, когда животное отвечает саккадическими (высокочастотными содружественными движениями глаз) и хватательным движением в направлении объекта (или без него). При поражении поля 7 двигательные реакции на важные объекты возникают с опозданием, и хватательные движения противоположной руки становятся неточными.
При появлении в поле зрения объекта, представляющего особый интерес, у добровольцев активируются все дополнительные зоны коры. Дорсолатеральная префронтальная кора (ДЛПФК, ориентировочно соответствует полю 46 по Бродману) — важная зона принятия решений, особенно решения о приближении или отступлении, показана на рисунке ниже и также упомянута ниже. На отдельном рисунке показан участок коры передней поясной извилины. Эта область подробно рассмотрена в отдельной статье на сайте, но упомянута здесь в связи с тем, что ее возбуждение дорсолатеральной корой возникает, когда человек уделяет внимание зрительной задаче.
• В относительно латеральном отделе расположены модули, определяющие форму всех типов объектов, включая форму букв. Это центр общей (категориальной/классической) идентификации объектов (например, собаки как таковой, без уточнения).
• В среднем отделе расположены модули, специфическая функция которых заключается в распознавании человеческих лиц.
• В относительно медиальном отделе находится отдел распознавания цвета, необходимый для определения всех цветов, кроме черного и белого. Ахроматопсия (цветовая слепота может развиться при поражении любого отдела зрительного проводящего пути) может возникать вследствие стойкого снижения артериального давления в обеих задних мозговых артериях, вызванного, например, эмболией в верхушку основного ствола базилярной артерии, сопровождающейся развитием инфаркта мозга. Такие пациенты видят мир черно-белым (в оттенках серого).
Угрожающие взгляды или лица приводят к возбуждению зонами 20 и 38 миндалевидного тела, особенно в правом полушарии; правое миндалевидное тело, в свою очередь, возбуждает отвечающую за страх правую орбитофронтальную кору, выделенную сиреневым цветом на рисунке ниже.
Этот путь участвует в обработке зрительной информации на ранних этапах, что приводит к стабилизации зрительной картины (внутренней модели внешнего мира, сохраняющейся благодаря обработке вестибулярной, соматосенсорной и зрительной информации), несмотря на постоянные движения глаз, а также облегчает восприятие зрительной картины и придает различные смыслы в зависимости от модели поведения.
и) Слуховая кора (поля 41,42 и 22). Первичная слуховая кора расположена в передней поперечной височной извилине Гешля, описанной в отдельной статье на сайте. Извилина Гешля соответствует полям 41 и 42 на верхней поверхности верхней височной извилины; большая часть информации от медиального коленчатого тела направляется к полю 41. Колончатая организация коры предположительно представлена в виде одночастотных полос, т.е. каждая из полос соответствует определенной частоте тона. Высокие частоты активируют латеральные полосы в извилине Гешля, а низкие частоты — медиальные полосы. В связи с неполным перекрестом центральных слуховых путей в стволе мозга (глава 20) каждое ухо имеет двустороннее представительство в коре. По данным исследований, первичная кора одинаково отвечает на монофоническую стимуляцию каждого слухового пути, однако кора противоположной стороны лучше отвечает на одновременную стимуляцию обоих слуховых путей.
Ассоциативная слуховая кора соответствует полю 22, обрабатывающему речь. Одновременная обработка зрительной и слуховой информации происходит в полимодальной коре, ограниченной верхней височной бороздой (соединение полей 21 и 22).
Удаление слуховой коры с одной стороны (например, при опухолевом процессе) не приводит к заметному ухудшению звуковосприятия. Единственный значительный дефект — потеря стереоакузии: при исследовании у пациента возникают трудности с определением локализации и расстояния от источника звука.
Видео физиология целенаправленного поведения (поведенческий акт) - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 27.2.2021

Вважається, що медіальна ділянка скроневої частки кори головного мозку відіграє важливе значення для декларативної та довготривалої пам’яті, але майже нічого не відомо про її роль у бінарних рішеннях та метакогніції.
Група вчених провела два експерименти. Спочатку досліджуваним показували продукт, наприклад, шоколадний батончик, і просили оцінити, наскільки сильно вони хотіли б з’їсти його зараз, після цього їм давали наїстися обраним. Дослід повторили 20 разів. Потім переходили до наступного тесту: показували фото двох продуктів і пропонували обрати один. Респонденти повинні були позначити, що їм подобається більше, з допомогою повзунка: чим ближче він був до одного з продуктів, тим сильніше вони були впевнені у своєму рішенні. Таким чином перевірили 190 пар різних закусок.

В тестах брали участь 12 чоловіків та жінок, всі страждали важкою формою епілепсії. Раніше їм імплантували внутрішньочерепні електроди, які точно визначають, активність якої зони мозку порушена. Вони також показують, як функціонують окремі нейрони, тому науковці використовували їх для проведення тестів. З допомогою електродів вони спостерігали, як працює мозок респондентів.
Команда зафіксувала активність 830 нервових клітин у скроневій зоні. Професор Унру-Пінейро, один з авторів дослідження, розповів, що частота імпульсів у деяких нейронах змінювалася в залежності від того, наскільки досліджувані були впевнені у своїх рішеннях.
Вчені змогли вперше спостерігати кореляцію між активністю нейронів і впевненістю в зробленому виборі. Також вони довели, що нейронна активність змінюється разом з реакцією респондента – і цей зв’язок зберігається до тих пір, поки вибір, перед яким стоїть людина, актуальний.

Дослідники відмітили, що вивчені нейрони знаходяться в області мозку, яка частково пов’язана з пам’яттю. Тому ймовірно, що надалі ми запам’ятовуємо не лише те, яке рішення прийняли, а й не, наскільки були впевнені у ньому. Професор Флоріан Морманн, один з науковців, які займалися дослідженням, припускає, що такий процес навчання може рятувати нас від неправильного вибору в майбутньому.
Дослідники з Боннського університету розповіли, що початково шукали зовсім інший феномен – вони збиралися визначити, яку роль скронева доля відіграє в тому, як ми оцінюємо предмети при виборі. Вчені не змогли підтвердити зв’язок суб’єктивної оцінки та активності нейронів, але дослідили, що деякі клітини мозку показують, наскільки людина впевнена у прийнятому рішенні.
Читайте также: