
Среди представителей саркодовых паразитический образ жизни ведет фораминиферы
Обновлено: 22.04.2024
Фораминиферы отряд простейших подкласса корненожек(рассматривается также как самостоятоятельный класс саркодовых, входящий в надкласс корненожек). Известны с кембрия. Размеры обычно 0,1—1 мм, редко до 20 см. наружный скелет в виде раковинок, у большинства известковых, изредка хитиноидных или состоящих из агглютинированных посторонних частиц (песчинок и т. п.). Раковинки однокамерные и многокамерные, расположены в один или два ряда, по спирали, иногда ветвящиеся. Через устье и поры раковинок выдаются тончайшие ветвящиеся и анастомозирующие псевдоподии (ризоподии). Для них характерно чередование полового и бесполого размножения. свыше 4000 современных видов (с ископаемыми около 30 тыс. видов). Все они морские, преимущественно бентосные организмы (за исключением 2 планктонных семейств Globigerinidae и Globorotaliidae). Раковины образуют значительную, часть океанических илов, морскоих осадков и осадочных пород.
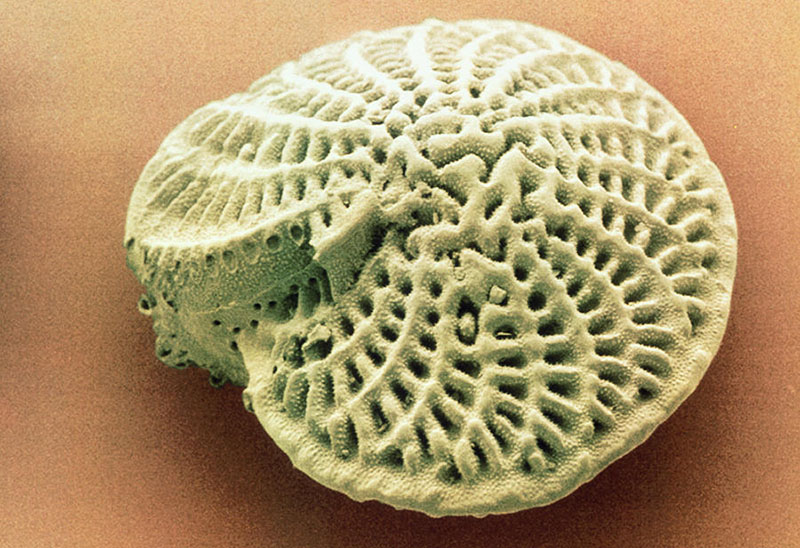
Фораминиферы строение раковина
Латинское название Foraminiferida
Фораминиферы Общая характеристика
Фораминиферы (лат. foramen – отверстие, дыра и fero – носить) – одноклеточные животные организмы, относимые к Простейшим, с сетью тонких разветвленных и соединяющихся между собой (анастомозирующих) псевдоподий (ложноножек). Это большая и разнообразная группа современных и ископаемых форм, насчитывающая в настоящее время более 34 000 видов, из которых около 4000 являются современными. Средние размеры их колеблются от 0,1 до 1 мм, однако некоторые формы могут достигать размера 10 мм и более. Цитоплазматическое тело фораминифер заключено в раковину различного состава и строения. Внутренняя полость раковины сообщается с внешней средой при помощи устья. Псевдоподии либо выходят только из устья раковины, либо образуют вокруг раковины сложную сеть, отходящую от наружной части цитоплазмы. Псевдоподии способны сокращаться и вытягиваться. В них осуществляется непрерывный ток цитоплазмы. По одной и той же псевдоподии одни струи ее текут в центростремительном (к раковине), другие – в центробежном направлении. Псевдоподии служат для улавливания и частично переваривания пищи, а также для передвижения животного.
Фораминиферы имеют сложный жизненный цикл, характеризующийся чередованием бесполого и полового поколений. Это в основном морские организмы. Небольшая часть фораминифер приспособилась к жизни в солоноватоводных бассейнах, а некоторые из них известны даже в пресноводных бассейнах. Большинство фораминифер являются бентосными формами (свободноживущими или прикрепленными), обитающими на разных глубинах. Некоторые ведут паразитический образ жизни на прикрепленных животных (мшанках, губках, кораллах). Планктонные формы значительно менее многочисленны; они живут на различных глубинах водного столба, будучи наиболее многочисленными в верхних пятидесяти метрах и особенно между 6 и 30 м от поверхности. Вертикальная миграция может быть суточной или в течение индивидуального развития.
Эти организмы распространены во всех морских бассейнах. Остатки их известны начиная с докембрия. Филогенетическое развитие сопровождалось усложнением строения раковины, которое было связано с прогрессивным усложнением строения и физиологических функций цитоплазматического тела.
Строение и физиология клетки

Форам.
Морфология раковин
Тип строения и терминология элементов раковины. Под типом строения подразумеваются характер последовательности нарастания камер и отчасти число камер, составляющих раковину. При этом один и тот же тип строения может иметь раковины как с агглютинированной, так и с секреционной стенкой. По числу камер все фораминиферы подразделяются на однокамерные, двухкамерные и многокамерные.
Однокамерные раковины имеют самую различную форму – трубчатую, трубчатую разветвленную или неправильно завернутую, звездчатую, древовидную, сферическую, полусферическую или субсферическую, колбовидную, лепешковидную. Они сообщаются с внешней средой с помощью одного или нескольких отверстий – устьев. У сферических и близких к ним форм имеется одно устье, расположенное в устьевой части (дистальном конце) раковины; часть раковины, противоположная устьевой, называется начальной (проксимальный конец). У трубчатых, звездчатых и других форм устьями являются свободные концы трубки.
Двухкамерные раковины состоят из начальной шарообразной камеры (пролокулюм) и второй трубчатой или ложнотрубчатой камеры. Трубчатая камера представляет собой настоящую трубку с собственной стенкой на всем протяжении, у ложнотрубчатой камеры в месте ее сочленения со смежным оборотом или с субстратом собственная минеральная стенка отсутствует, а имеется лишь тонкая органическая мембрана. Вторая – трубчатая или ложнотрубчатая – камера двухкамерных раковин может быть свернута в плоскую или коническую спираль или в неправильный клубок, у некоторых форм вторая камера представляет собой прямую несвернутую часть раковины или может быть свернутой зигзагообразно.
Стенка раковины у фораминифер, способ образования, состав и структура
Палеонтологи придают большое значение изучению состава и структуры стенки раковин фораминифер, рассматривая их в качестве ведущего признака в систематике этой группы организмов.
По составу и способу образования различают три типа раковин: органические, агглютинированные и секреционные известковые. Много новых данных по структуре раковин было получено за последние десятилетия благодаря использованию электронной микроскопии и рентгеноструктурного анализа.
Размножение, жизненный цикл, полиморфизм раковин
Система и характеристика высших таксонов
Система фораминифер находится в последнее время в состоянии активной разработки, но до сих пор отсутствует единое представление о таксономическом ранге этой группы и классификации ее на уровне высших таксонов. В настоящей работе принимается один из новых вариантов классификации этой гоуппы согласно изданию (Маслакова и др., 1995), учитывающий взгляды авторов предшествующих классификационных схем и новые представления. Здесь фораминиферы рассматриваются в ранге класса типа саркодовых. В составе класса выделяются 8 подклассов и 29 отрядов. При построении системы использовалась совокупность всех возможных критериев, т. е. морфологического, онтофилогенетического, геохронологического и экологического, которые определяют сочетание признаков, свойственных тому или иному таксону.
Основой классификации по-прежнему остается морфология раковины, поскольку, судя по имеющимся, хотя и немногочисленным, данным по цитологии и жизненным циклам, прогрессивное усложнение мягкого тела сопровождалось в эволюции этой группы многообразием и усложнением строения раковины. Наиболее важными признаками, положенными в основу выделения подклассов, являются определяющиеся природой цитоплазмы химический состав, способ образования и микроструктура стенки раковины. В качестве дополнительных признаков при выделении подклассов использованы такие особенности строения секреционных известковых раковин, как отсутствие или наличие и характер пористости в стенке раковины, наличие каналовой системы у специализированных форм, особая форма камер, специфический характер устья, образ жизни (экологический тип: бентосный или планктонный). Приняты во внимание данные по цитологии (особенности ядерного аппарата, присутствие в цитоплазме планктонных форм везикулярного ретикулума и криптосом), а также особенности жизненных циклов и характер гамет.
Для характеристики отрядов использовано сочетание таких признаков, как отсутствие или. наличие септации (однокамерные, двухкамерные и многокамерные раковины с учетом вторичного появления однокамерных форм у многих секреционных известковых форм), тип строения раковины или их определенное сочетание, преобладающий характер устья, форма камер, наличие каналовой системы, характер ранних стадий онтогенеза. При этом учитывалось, что таксономическое значение некоторых морфологических признаков не оставалось постоянным, а менялось в ходе исторического развития фораминифер (обычно повышалось).
Класс Foraminifera Orbigny, 1826
Саркодовые, тело которых заключено в различную по способу образования и составу раковину: органическую, агглютинированную или секреционную известковую (преимущественно кальцитовую или арагонитовую). Раковины имеют одно или несколько устьев.
Жизненные циклы с чередованием полового и бесполого поколений, с промежуточной редукцией; гаметы обычно двухжгутиковые, часто разножгутиковые (один из жгутиков с двумя рядами ресничек), реже трехжгутиковые, могут быть вторично-амебоидными.
Ядерный аппарат со сложными изменениями в течение жизненного цикла; лишь у низших форм часть цикла одноядерна, а у остальных многоядерность сохраняется в течение всего жизненного цикла (кроме зиготы). При этом у наиболее продвинутых форм возникает ядерный дуализм агамонтов (шизонтов), реже гамонтов.
Преимущественно морские бентосные свободноживущие, реже прикрепленные организмы; небольшая часть перешла к планктонному образу жизни, изредка наблюдается переход к паразитизму. Докембрий – современные.
Класс включает 8 подклассов: Allogromiata, Textulariata, Fusulinata, Miliolata, Lagenata, Rotaliata, Spirillinata, Globigerinata.
Экология и тафономия
Являются преимущественно морскими организмами, но кроме типично морских бассейнов некоторые их представители заселяют солоноватоводные, даже иногда пресные водоемы. Так, известны находки фораминифер в солоноватоводных и пресных бассейнах Сахары, ряд видов заходит на различное расстояние в эстуарии рек. Некоторые формы сохранились в виде реликтов в водоемах, отделившихся от моря и сильно опресненных (оз. Эльтон). Известны находки фораминифер в грунтовых водах и соленых колодцах Средней Азии. Предполагается, что это реликты миоценового моря, когда-то заходившего на территорию теперешних Каракумов. В условиях пониженной солености раковины фораминифер тонкие, при повышенной солености – толстые. Для фораминифер, живущих в условиях значительного опреснения, характерна утрата извести вплоть до образования чисто органической раковины.
Существуют две биономические группы фораминифер: бентосная и планктонная.
Этапы развития
Развитие фораминифер происходило в направлении усложнения морфологии раковины и появления различных типов стенки, а соответственно и новых таксонов. На основании смены систематического состава фораминифер в истории развития этой группы организмов установлены семь основных этапов:
- 1) раннепалеозойский, включающий ордовик и силур, к ним может быть; добавлен кембрий и частично докембрий;
- 2) среднепалеозойский (девон–ранний карбон);
- 3) позднепалеозойский (средний карбон – пермь);
- 4) – триасово-раннемеловой;
- 5) позднемеловой;
- 6) палеогеновый;
- 7) неоген-антропогеновый
Стратиграфическое значение
Фораминиферы имеют большое значение для разработки детальной стратиграфии, местной, межрегиональной и субглобальной корреляции. Это определяется тем, что их раковины в ряде случаев присутствуют в породе в больших количествах, благодаря чему возможны подробная послойная характеристика и датировка разреза. Кроме того, представители ряда групп фораминифер, такие, как фузулиниды, эндотириды, нуммулитиды, милиолиды, глобигериниды и другие, быстро эволюционировали, в связи с чем можно расчленять разрез на стратоны различного ранга по отдельным видам или их комплексам. Небольшие размеры раковин фораминифер позволяют извлекать их в достаточном количестве и из кернового материала, что придает им особую ценность при проведении буровых работ – расчленении и сопоставлении разрезов скважин.
Разные группы фораминифер дают возможность детально расчленять разрезы и выделять различные по широте распространения стратоны. Если по бентосным фораминиферам разрабатываются главным образом местные и региональные схемы, то по планктонным – субглобальные, простирающиеся в пределах одного климатического пояса. На основании изучения видов устанавливаются характерные комплексы для групп ярусов, отдельных ярусов, подъярусов; при более детальных исследованиях выделяются слои с видами или зоны, отвечающие обычно части подъяруса или подъярусу. При частом отборе образцов, когда улавливаются даже незначительные изменения фауны, и при детальном изучении филогении той или иной группы могут быть выделены филозоны, отражающие определенные этапы в эволюции этих организмов.
Хотя находки фораминифер известны начиная с докембрия и некоторое стратиграфическое значение имеют их более примитивные представители при расчленении силура и девона, но достаточно дробная зональная стратиграфия по этой группе ископаемых разработана начиная с карбона.
Схемы расчленения мезозойских и кайнозойских отложений первоначально были разработаны на материале континентов, в последние десятилетия более дробные схемы получены при исследовании из океанических скважин.
Фузулиниды имеют большое практическое значение для детального расчленения каменноугольных и пермских отложений таких регионов, как Поволжье, Западное Приуралье, Западная Европа, Северная Америка и др. Для разработки стратиграфии триасовых и юрских отложений главную роль играют лагениды (в триасе они составляют 70 % всех известных видов), а также милиолиды, эпистоминиды и в меньшей степени роталииды. Наиболее дробно стратиграфия триасовых отложений разработана для юга Западной Европы (особенно для Альп), юга бывшего СССР, Аляски. Для юры дробные стратиграфические схемы с выделением зон созданы для платформенных областей, в частности, для Восточно-Европейской и Западно-Сибирской платформ, и менее дробные для геосинклинальных областей.
В начале раннего мела в значительной степени наследуется систематический состав фораминифер позднеюрского времени, и руководящими формами на протяжении берриаса – баррема являются также лагениды, роталииды, эпистоминиды, некоторые атаксофрагмииды и литуолиды. Начиная с апта широкое распространение получают планктонные фораминиферы, которые делают возможным разработку зональной стратиграфии для отложений геосинклинальных областей. В областях развития платформенных отложений главная роль принадлежит роталиидам (Gavelinellidae и др.), лагенидам, эпистоминидам и агглютинированным формам.
Чрезвычайно велико практическое значение планктонных форм для разработки зональной стратиграфии верхнемеловых и палеогеновых отложений. Швейцарским микропалеонтологом Г. Болли в 1966 г. была предложена субглобальная зональная схема для меловых и кайнозойских отложений тепловодного пояса. Со временем эта схема совершенствовалась другими исследователями.
Наряду с планктоном большую роль в расчленении верхнемеловых и кайнозойских отложений играет и бентос. Так, зональная схема расчленения верхнего мела п-ова Мангышлак по аномалинидам широко применяется и для расчленения разрезов Восточно Европейской платформы; многочисленны работы по расчленению палеогеновых отложений Западной Европы и юга бывшего СССР по нуммулитидам.
Стратиграфия неогеновых и четвертичных отложений также разрабатывается как по планктонным, так и по бентосным фораминиферам. Среди последних наиболее важны милиолиды, различные роталииды (Rotaliidae, Elphidiidae, Nonionidae, Cassidulinidae).
Результаты изучения используются не только в стратиграфии, но и при решении вопросов фациального анализа, палеогеографии и палеозоогеографии. Отдельные экологические типы приурочены к определенным условиям разных зон морей и океанов. Это особенно хорошо устанавливается при исследовании современных фораминифер, полученные результаты с определенными поправками могут быть перенесены на ископаемый материал для восстановления палеообстановок.
Методика изучения
Раковины встречаются практически во всех породах морского происхождения, но преобладают в карбонатных глинах, мергелях, органогенно-обломочных известняках. Процесс исследования фораминифер состоит из сбора образцов в поле, технической обработки пород и научной обработки материала.

Саркодовые
Общая Характеристика Саркодовых
Саркодовые, что значит плазматические, представляют собой самый большой класс простейших, включающий более 11000 видов и отличающийся большим разнообразием форм.
Класс саркодовых объединяет животных, ведущих весьма различный образ жизни. Большинство — жители моря, но немало среди них и пресноводных форм. Некоторые Саркодовые приспособились к обитанию в почве и в моховых подушках торфяных болот и лесов. Паразитические формы относительно редки, не превышают 1,3% от свободноживущих.
Саркодовым свойственно все время менять форму тела, что связано с образованием псевдоподий. Ранее непостоянство формы тела объясняли отсутствием клеточной оболочки, которую невозможно было практически различить в оптический микроскоп. Однако при помощи электронного микроскопа у них можно обнаружить тончайшую мембрану, или плазмалемму, имеющую фибриллярное строение. Цитоплазма простейших образует два слоя: наружный, более плотный — эктоплазму и внутренний, более жидкий — эндоплазму. У них эти слои легко переходят в процессе жизни друг в друга. Плазмалемма также способна нарушаться и восстанавливаться. По-видимому, это непостоянство слоев цитоплазмы и отсутствие плотной стабильной оболочки обусловливают неопределенность формы тела, столь характерную для саркодовых.
Многие саркодовые образуют в той или иной форме внутренний скелет или наружный скелет в виде раковин.
Органеллами движения служат псевдоподии, или ложноножки. Псевдоподии представляют собой временные плазматические выросты различной формы.
Псевдоподии их — не только органеллы движения, но и органеллы захвата пищи. У многих плавающих форм псевдоподии могут быть только ловчими образованиями.
Питаются бактериями, водорослями и различными прстейшими. Переваривание захваченной пищи происходит в особых органеллах — пищеварительных вакуолях, которые образуются в цитоплазме. Это пузырьки жидкости с пищеварительными ферментами, выделяемыми цитоплазмой вокруг пищевых частиц. У пресноводных сар- кодовых обычно имеются также выделительные органеллы — сократительные вакуоли.
Таксон Саркодовые разделяется на 5 отрядов:
Первые три отряда (амебы, раковинные амебы и фораминиферы) многими зоологами (В. А. Догель и др.) объединяются в подкласс корненожек (Rhizopoda) на основании наличия некоторых общих признаков. В таком случае следующие два отряда — лучевики и солнечники — рассматриваются не как отряды, а как подклассы наряду с подклассом Rhizopoda.
ОТРЯД 1. АМЕБЫ (AMOEBIDA)
Амебы — преимущественно обитатели пресных водоемов, встречаются также и в морях, в микроводоемах прибрежного песка и почвы. В почве амебы наряду с другими простейшими обнаружены в активном состоянии. Они питаются почвенными бактериями.
В пресных водоемах на растениях, на гниющих листьях и различных Подводных предметах можно найти несколько различных видов амеб, в том числе особенно крупную Amoeba proteus (рис. 3). Подробнее >>
ОТРЯД 2. РАКОВИННЫЕ АМЕБЫ (TESTACEA)
Раковинные амебы живут исключительно в пресных водах, наиболее многочисленны они среди влажных мхов на болотах.
Тело раковинных амеб, в отличие от голых амеб, одето раковиной, в остальном строение представителей этих двух отрядов очень сходно. Раковина обычно состоит из хитиноидного вещества, которое выделяется эктоплазмой. У некоторых форм раковина более твердая: она образована пластиночками из кремнезема или в ее органическую основу включены песчинки. Раковина всегда состоит из одной камеры и представляет собой шапочку, колпачок или бутылочку, на нижней поверхности которой находится отверстие — устье. Через устье высовываются псевдоподии амебы. Подробнее >>
Фораминиферы
Рассматривается как самостоятельный тип или класс, входящий в надкласс корненожек. Известны с кембрия. Размеры обычно 0,1—1 мм, редко до 20 см. Наруж. скелет в виде раковинок, у большинства известковых, изредка хитино- идных или состоящих из агглютинированных посторонних частиц (песчинок и т. п.). Раковинки однокамерные и многокамерные, расположены в один или два ряда, по спирали, иногда ветвящиеся. Через устье и поры раковинок выдаются тончайшие ветвящиеся и анастомозирую- щие псевдоподии (ризоподии). Для фораминиферы характерно чередование полового и бесполого размножения. Свыше 1000 современных видов (с ископаемыми около 30 тыс. видов). Все фораминиферы— морские, преимущественно бентосные организмы (за исключением 2 планктонных семейств Globigerinidae и Globorotali- idae). Раковины Ф. образуют значит, часть океанических илов, морских осадков и осадочных пород. Подробнее >>
ОТРЯД 4. ЛУЧЕВИКИ (RADIOLARIA)
Лучевики, или радиолярии,— богатая видами группа исключительно морских организмов (свыше 14000 видов, современных около 1000 видов). Они ведут только плавающий (планктонный) образ жизни. Особенно много радиолярий в теплых морях с достаточно большой соленостью, в которых они могут преобладать в планктоне. Многие из них имеют шаровидную форму тела с лучеобразно расположенными псевдоподиями (рис. 14).
Центральная часть тела лучевиков, состоящая из эндоплазмы и части эктоплазмы, содержит одно или несколько ядер. Она заключена в харак-терную для лучевиков центральную хитин о и дну ю капсулу. Цитоплазму внутри капсулы называют капсулярной, а вне ее — в н е- капсулярной. Нередко внекапсулярная плазма содержит пигмент и различные секреты (рис. 14, А; 15). Центральная капсула имеет отверстия, через которые внутрикапсулярная плазма сообщается с внекапсу- лярной. Внекапсулярная плазма толстым слоем окружает центральную капсулу, она сильно вакуолизирована (рис. 14, А), что способствует уменьшению плотности животного и может рассматриваться как приспособление к планктонному образу жизни.
Во внекапсулярпой плазме, а отчасти и внутри центральной капсулы у большинства лучевиков обитают симбиотические^ одноклеточные водоросли зоохлореллы. Они потребляют углекислоту, образуемую радиоляри- ями, а последние используют выделяемый водорослями кислород.
Псевдоподии радиолярий берут начало во внутрикапсулярной плазме, проходят через поры центральной капсулы, через внекапсулярную плазму и радиально расходятся во все стороны. Они длинные, тонкие, часто вет-вящиеся, образующие сплетения и анастомозы, т. е. как и у фораминифер, это типичные ризоподии. Питаются радиолярии жгутиконосцами, инфузо-риями, мелкими водорослями, которых они захватывают своими ризопо- дпями. Пищеварительные вакуоли образуются внутри пучка рпзоподий или во внекапсулярпой плазме. Сократительные вакуоли у радиолярий от-сутствуют.
Некоторые радиолярии не имеют иного скелета, кроме центральной капсулы. У большинства радиолярий имеется более или менее сложно устроенный скелет, состоящий из кремнезема или сернокислого стронция. В простейшем случае он состоит из кремневых иголочек, более или менее густо расположенных на поверхности внекапсулярной плазмы. У ряда форм скелет состоит из правильно расположенных радиальных игл (рис. 14, Б\ 15), у других форм скелетные иглы могут соединяться перемычками, образующими сложную решетку, покрывающую тело (рис. 14, В и Г). У многих радиолярий скелет сильно усложнен и имеет причудливую форму.
Радиолярии живут на различных глубинах (до 7000 м), у некоторых видов имеются приспособления, при помощи которых они могут подниматься или опускаться в толще воды. Так, у радиолярии Acanthometra (рис. 15) скелет состоит из правильно расположенных (радиально) игл. К иглам прикрепляются мускульные волоконца — миофриски, котоые другим концом соединены с поверхностью плазмы.
Когда волоконца сокращаются, они растягивают поверхностный слой эктоплазмы и объем радиолярии увеличивается, а ее плотность уменьшается, при этом животное поднимается в верхние слои воды. При расслаблении мио- фрисков объем животного уменьшается, а плотность его увеличивается и оно погружается медленно в воду (рис. 14,Б, Вг; 15).
А — Thalassicolla (внутри — центральная капсула, окруженная вакуолизированной внекапсулярной плазмой, снаружи — многочисленные псевдоподии); Б и Б, — Acantharia, изменяющая объем вследствие сокращения миофрисков; В — скелет Heliosphaera; Г — Theopilium; Д — колониальная радиолярия Collozoum (много центральных капсул, окруженных общей внекапсулярной плазмой).
Радиолярии размножаются как бесполым, так и половым путем. При бесполом размножении тело радиолярии делится надвое, сначала митотически делится ядро, затем — центральная капсула и минеральный скелет, позднее всего делится внекапсулярная плазма. При задержке деления внекапсулярной плазмы образуются колонии, содержащие множество центральных капсул в единой массе студенистого вещества (рис. 14, Д). Колонии возпикают в результате незавершенного процесса бесполого размножения.
У многих радиолярий (Acantharia, Spumellaria и др.) прослежено об-разование маленьких гамет с двумя жгутиками. У Acantharia наблюдались копуляция гамет и формирование зиготы. Правильного чередования бесполого и полового размножения у радиолярий не наблюдалось.
Подобно тому как скелеты фораминифер входят в со- Практпческое став ила морского дна и участвуют в образовании осадочных пород, скелеты радиолярий образуют на дне океана радиоляриевый ил, часто на очень больших глубинах.
Скелеты радиолярий, состоящие из кремнезема, хорошо сохраняются. Некоторые горные породы состоят сплошь из их остатков. Эти породы из-вестны под названием горной муки или трепела и употребляются для шлифовки металлических изделий и других технических целей. В России залежи трепела имеются на Урале, в Поволжье и других районах.
Скелеты радиолярий, так же как и скелеты фораминифер, служат руководящими ископаемыми при определении возраста осадочных пород. Подробнее >>
ОТРЯД 5. СОЛНЕЧНИКИ (HELIOZOA)
Солнечники — преимущественно пресноводные животные, внешне во многом похожие на радиолярий. Они имеют шаровидную форму тела с прямыми, расходящимися по радиусам псевдоподиями, напоминающими солнечные лучи.
В наших пресных водах обычны крупный солнечник Actinosphaerium eichhorni, до 1 мм в диаметре, Actinophrys sol и др.
В отличие от радиолярий у солнечников нет центральной капсулы. Цитоплазма резко разделяется на зернистую эндоплазму, содержащую одно (Actinophrys) или несколько (Actinosphaerium) ядер, и более прозрачную и сильно вакуолизированную эктоплазму, в которой находятся одна или две сократительные вакуоли.
Псевдоподии солнечников начинаются в эндоплазме, они прямые, не анастомозирующие. В середине псевдоподии проходит более плотная осевая нить, покрытая снаружи слоем жидкой цитоплазмы. Подобные псевдоподии называют аксоподиями. Аксоподии могут удлиняться и укорачиваться, а также изгибаться. Благодаря этому солнечники могут захватывать аксоподиями довольно крупную добычу.
У большинства солнечников нет твердого скелета, но у некоторых имеются мелкие кремневые иглы. Подробнее >>
Саркодовые — группа одноклеточных организмов, которая ранее рассматривалась в качестве класса или подтипа. К ним относятся простейшие, не имеющие постоянной формы тела, они покрыты мембраной, состоящей из белков и липидов. Микроорганизмы не имеют оболочки, но они могут образовывать раковину или внутренний скелет.
Общее строение
Класс саркодовые представляет собой большую группу микроорганизмов, который объединяет более 11 тыс. видов, отличающихся большим разнообразием форм. Он включает простейших, которые ведут различный образ жизни.

Большую часть составляют морские обитатели, но существуют и пресноводные микроорганизмы. Некоторые простейшие могут существовать в грунте, мхе торфяных болот и лесов. Реже встречаются паразитические представители, численность которых составляет чуть больше одного процента от остальных микроорганизмов.
Тип саркодовые постоянно изменяет внешний вид, так как происходит формирование псевдоподий (ложноножек). Несколько лет назад биологи объясняли изменчивость простейших отсутствием покрытия вокруг клетки, которое трудно было рассмотреть даже в оптический прибор.
Если использовать электронный микроскоп, то можно увидеть тончайшую мембрану, которая имеет фибриллярное строение. Плазмалемма саркодовых представителей формирует два слоя:
- наружную плотную эктоплазму;
- внутреннюю жидкую эндоплазму.
В течение жизненного цикла оба слоя легко взаимодействуют между собой, кроме того, клеточная мембрана способна разрушаться и восстанавливаться. По-видимому, такая изменчивость слоев плазмалеммы и отсутствие плотного покрытия объясняет неопределенность конфигурации тела саркодовых.

Часть микроорганизмов обладает внутренним или наружным скелетом в виде раковин. Передвижение простейшие осуществляют с помощью псевдоподий, или ложноножек. Они представляют собой отростки разной конфигурации и предназначены также для захвата пищи.
В питание микроорганизмов входят водоросли и обыкновенные простейшие, которые перевариваются вакуолями, образующимися в цитоплазме. Это пузырьки жидкости с пищеварительными ферментами, которые выделяет плазмалемма вокруг пищевых частиц.
У пресноводных микроорганизмов имеются органы выделения — сократительные вакуоли. Саркодовые делятся на 5 отрядов:
- Амебы.
- Раковинные амебы.
- Фораминиферы.
- Лучевики.
- Солнечники.
Первые 3 отряда относятся к корненожкам, а лучевики и солнечники рассматриваются не как отряды, а как подклассы.
Отряд корненожек
Самым простым микроорганизмом, относящимся к корненожкам, считаются голые амебы. Но особенно часто встречаются обыкновенные простейшие, или протей.
Амеба обыкновенная
Обыкновенная амеба обитает в пресных водах, маленьких прудах и канавах с илистым дном, кроме того, микроорганизмы легко размножаются в условиях лаборатории. Это самый крупный представитель среди существующих простейших. Когда он передвигается, то может вырасти до 0,5 мм и его видно невооруженным глазом.
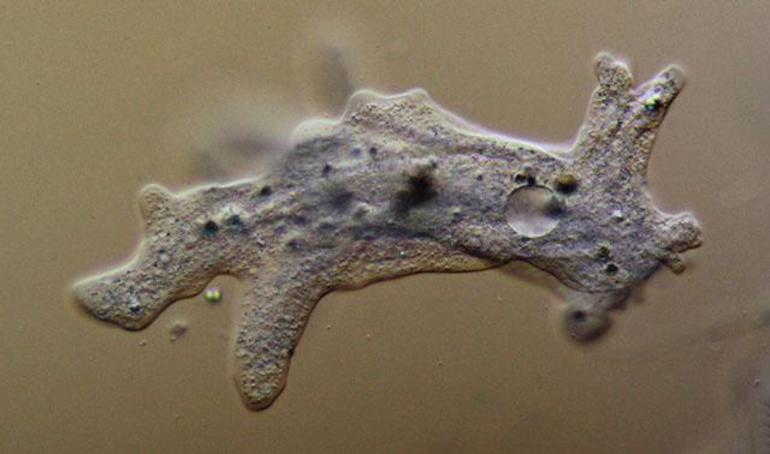
Если смотреть за амебой через микроскоп, то можно заметить несколько длинных ложноножек, напоминающих лопасти. Форма тела саркодовых постоянно изменяется, часть псевдоподий втягивается внутрь, а некоторые, наоборот, удлиняются. Ложноножки в нескольких точках закрепляются к почве, что помогает амебе передвигаться.
Если простейшее сталкивается с пищевой органикой, то оно обхватывает всю частицу и помещает ее внутрь плазмалеммы вместе с жидкостью. Так в теле образуются пузырьки с пищей, которые называют пищеварительными вакуолями, и в них происходит процесс переваривания органики.
Кроме того, в цитоплазме обыкновенного простейшего периодически появляется прозрачный пузырек. Это сократительная вакуоль, которая выполняет для микроорганизма важные жизненные задачи. Она набирает в себя воду из цитоплазмы, увеличиваясь в размерах, а затем сокращается и выбрасывает ее через отверстие наружу.
Вся эта процедура происходит в течение 5—8 минут. В теле амебы количество органических и неорганических веществ выше, чем в окружающей жидкости, поэтому вода поступает в протоплазму микроорганизма. Благодаря сократительной вакуоли, ее избыток выбрасывается наружу. Если бы этого не происходило, то амеба просто растворилась бы в воде.
Вместе с жидкостью выделяются продукты обмена веществ, то есть сократительная вакуоль принимает участие в процессе выделения. С водой в микроорганизм поступает кислород, таким образом, органоид помогает простейшему организму дышать. Как и в любой клетке, в теле амебы находится ядро, которое практически не видно, поэтому для его обнаружения специалисты применяют красители.
Размножение микроорганизма происходит единственным способом — это деление на две клетки при свободном передвижении. Оно происходит через центр ядра, которое разделяется на две половины. Периодичность этого процесса зависит от температуры обитания, питания и происходит один раз через 1—2 суток.
Паразитические микроорганизмы
Некоторые виды простейших могут вести паразитический образ жизни, поселившись в кишечнике позвоночных и беспозвоночных животных. У человека в кишечной полости обитают пять видов микроорганизмов. Четыре из них считаются безобидными, но пятый, который носит название дизентерийной амебы, может вызывать тяжелое заболевание — амебиаз.
Этот вид простейших представляет собой очень мелкие микроорганизмы (20—30 мк), которые живут в толстом кишечнике. Основной их характеристикой считается высокая подвижность. Амеба существует практически во всех уголках мира, но заболевание амебиаз встречается довольно редко, только в южных и теплых регионах.
Это объясняется тем, что наличие паразитического микроорганизма не всегда сопровождается заболеванием. В этом случае дизентерийная амеба выполняет функцию носительства, то есть она передвигается в кишечной среде, питается бактериями, но не вызывает патологических изменений. Но иногда микроорганизм активно начинает проникать в стенки кишечника.
Такое действие вызывает у человека тяжелую форму кровавого поноса. В этот период амеба меняет объект питания, начиная поедать красные кровяные клетки вместо бактерий. Если не пройти курс лечения, то амебиаз перейдет в хроническую форму, вызывая у человека сильное истощение.
Дизентерийные микроорганизмы могут жить только в организме человека, а попадая в почву или воду, они сразу погибают. Заражение осуществляется непостоянными формами существования — цистами. Когда простейшие из толстого кишечника попадают в прямую кишку, то они меняют свою форму. Они втягивают ложноножки, выкидывают пищевые частицы и становятся круглыми, а эктоплазма образует тонкое, но прочное покрытие.
Этот процесс называется инцистирование. Особенностью этого момента считается деление ядра последовательно два раза, образуя характерные четырехъядерные цисты. В таком виде они с фекалиями выводятся наружу, где продолжают существовать в течение 2—3 месяцев. Если паразитическое простейшее попадает в организм человека, то оно эксцистируется.
Тело разделяется два раза, при этом ядро не затрагивается. В итоге образуются четыре одноядерных микроорганизма, которые приступают к активной жизнедеятельности. К паразитическим микроорганизмам относят и ротовую амебу, которая обитает в полости рта человека.
Подкласс лучевики
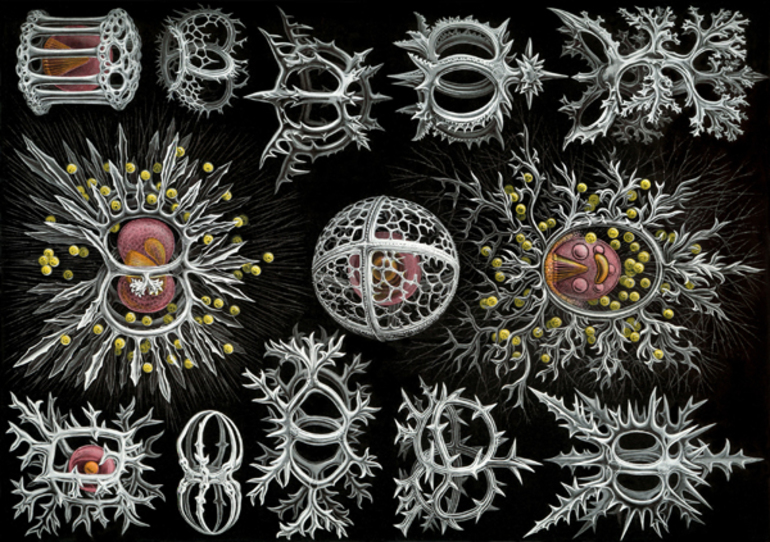
Самый многочисленный по числу видов морской подкласс в биологической систематике саркодовых составляют лучевики, или радиолярии. Он насчитывает более 7—8 тыс. видов. Лучевики относятся к планктонным организмам, которые в основном живут в морских водах тропических и субтропических зонах.
Размеры тела радиолярий колеблются в пределах от 50 мк до 1 мм и более. Существуют некоторые объединения саркодовых, обладающих гораздо крупным телом. Значительная часть радиолярий имеет форму сферы с характерной капсулой в центре. Это органическая мембрана, расположенная вокруг цитоплазмы с ядром.
Основная их масса — это слизь, образующая мощный слой. Кроме слизи, в цитоплазме лучевиков встречаются капли жира. Все эти образования облегчают саркодовые и помогают им свободно передвигаться в морском пространстве. У некоторых радиолярий внутри цитоплазмы встречаются одноклеточные водоросли из отряда панцирных жгутиковых.
Это доказывает процесс симбиоза простейших организмов с растительным миром. Водоросли в теле простейших получают питательные вещества и углекислый газ, который образуется в результате дыхания. Подкласс лучевиков делится на четыре отряда:

- Спумеллярии.
- Назеллярии.
- Акантарии.
- Феодарии.
Все они отличаются типичными формами скелетных образований. Недостаточно изучены процессы размножения лучевиков, хотя ученые длительное время занимались исследованием этих простейших. Проблема в том, что радиолярии долго не живут в лабораторных условиях, поэтому не удается получить полные результаты.
Скелетное образование не позволяет им делиться на две части как в случае с амебами. Скорее всего, формирование зародышей с одним ядром у них происходит как при бесполом размножении фораминифер. Этот процесс у радиолярий требует продолжения дальнейших исследований.
Самым обширным отрядом среди корненожек являются обитатели моря — фораминиферы (Foraminifera). В составе современной морской фауны известно свыше 1000 видов фораминифер. Небольшое число видов, представляющих, вероятно, остаток морской фауны, обитает в подпочвенных соленых водах и солоноватых колодцах Средней Азии.
Подобно раковинным корненожкам, все фораминиферы имеют раковину. Строение скелета достигает здесь большой сложности и огромного разнообразия.
В океанах и морях фораминиферы распространены повсеместно. Их находят во всех широтах и на всех глубинах, начиная от прибрежной литоральной зоны и кончая самыми глубокими абиссальными впадинами. Все же наибольшее разнообразие видов фораминифер встречается на глубинах до 200—300 м. Подавляющее большинство видов фораминифер является обитателями придонных слоев, входят в состав бентоса. Лишь очень немногие виды живут в толще морской воды, являются планктонными организмами.

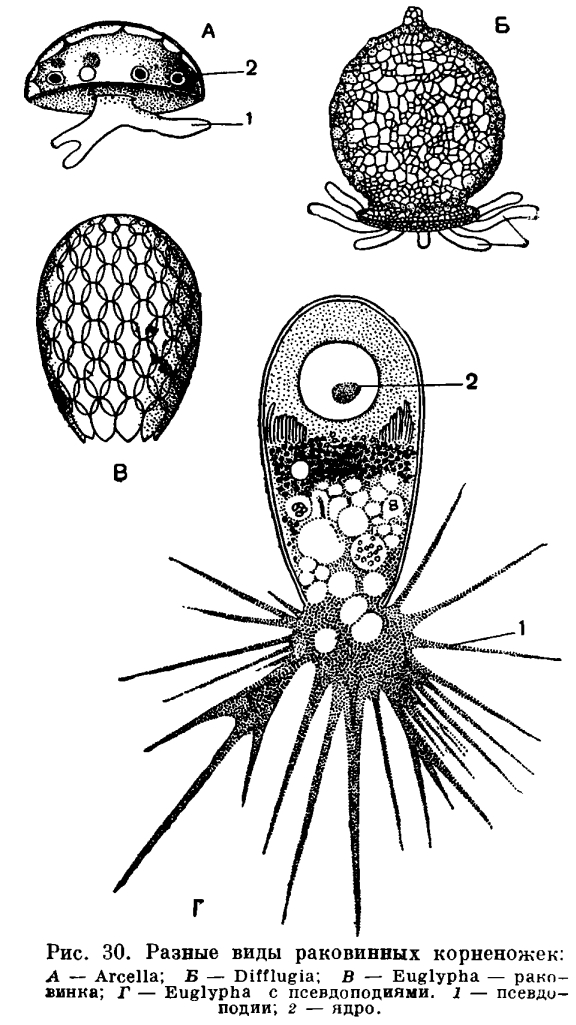
Среди огромного разнообразия строения раковин фораминифер можно различить по составу их два типа. Одни из них состоят из посторонних телу корненожки частиц — песчинок. Подобно тому как мы это видели у диффлюгии (рис. 30), фораминиферы, обладающие такими агглютинированными раковинами, заглатывают эти посторонние частицы, а затем выделяют их на поверхности тела, где они закрепляются в тонком наружном кожистом слое цитоплазмы. Такой тип строения раковины имеют часто встречающиеся представители родов Hyperammina, Astrorhiza (рис. 32, 3—7) и др. Например, в некоторых районах наших северных морей (море Лаптевых, Восточно-Сибирское море) эти крупные фораминиферы, достигающие 2—3 см длины, почти сплошным слоем покрывают дно.
Число видов фораминифер с агглютинированной раковиной относительно невелико (хотя число особей этих видов может быть огромным). Большая часть обладает известковыми раковинами, состоящими из углекислого кальция (СаС03).
Эти раковины выделяются цитоплазмой корненожек, которые обладают замечательной особенностью концентрировать в своем теле кальций, содержащийся в морской воде в небольших количествах (соли кальция в морской воде составляют немногим более 0, 1%). Размеры известковых раковин разных видов фораминифер могут быть очень различны. Они варьируют в пределах от 20 мк до 5—6 см. Это примерно такое же соотношение размеров, как между слоном и тараканом. Наиболее крупных из фораминифер, раковина которых имеет 5—6 см в диаметре, уже нельзя назвать микроскопическими организмами. Наиболее крупные (роды Cornuspira и др.) живут на больших глубинах.
Однокамерные фораминиферы имеют одну-единственную полость внутри раковины, которая сообщается с наружной средой устьем. Форма однокамерных раковин разнообразна. У одних (например, Lagena) раковинка напоминает бутылочку с длинным горлышком, иногда снабженную ребрами (рис. 32, 2).
Очень часто происходит спиральное закручивание раковины, и тогда внутренняя полость ее становится длинным и тонким каналом (например, Ammodiscus, рис. 32, 8, 9).
Большинство известковых раковин корненожек является не однокамерными, а многокамерными. Внутренняя полость раковины поделена перегородками на ряд камер, число которых может достигать нескольких десятков и сотен. Перегородки между камерами не сплошные, в них имеются отверстия, благодаря чему протоплазматическое тело корненожки не расчленено на части, а представляет собой единое целое. Стенки раковинок не у всех, но у многих фораминифер пронизаны мельчайшими порами, которые служат для выхода наружу псевдоподий. Об этом подробнее будет сказано ниже.
Число, форма и взаимное расположение камер в раковинке может быть очень различным, что и создает огромное разнообразие фораминифер (рис. 32). У некоторых видов камеры расположены в один прямой ряд (например, Nodosaria, рис. 32, 12), иногда же их расположение двурядное (Textularia, рис. 32, 22). Широко распространена спиральная форма раковины, когда отдельные камеры расположены по спирали, причем по мере приближения к камере, несущей устье, размеры их возрастают. Причины этого постепенного нарастания размеров камер станут понятны, когда мы рассмотрим ход их развития.
В спиральных раковинках фораминифер имеется несколько оборотов спирали. Наружные (более крупные) обороты могут быть расположены рядом с внутренними оборотами (рис. 32, 17, 18) так, что все камеры видны снаружи. Это эволютный тип раковины. У других форм наружные (более крупные) камеры целиком или частично охватывают внутренние камеры (рис. 33, 1). Это инволютный тип раковины. Особую форму строения раковины находим у фораминифер милиолид (семейство Miliolidae, рис. 32, 19). Здесь камеры сильно вытянуты параллельно продольной оси раковины и расположены в нескольких пересекающихся плоскостях. Вся раковина в целом оказывается продолговатой и по форме несколько напоминает тыквенное зерно. Устье расположено на одном из полюсов и обычно снабжено зубцом.
Большой сложностью строения отличаются раковинки, относящиеся к циклическому типу (роды Archiacina, Orbitolites и др., рис. 33, 2, 34). Число камер здесь очень велико, причем внутренние камеры располагаются по спирали, наружные же — концентрическими кольцами.
В чем заключается биологическое значение столь сложного строения многокамерных раковинок корненожек? Специальное исследование этого вопроса показало, что многокамерные раковинки по сравнению с однокамерными обладают гораздо большей прочностью. Основное биологическое значение раковинки — это защита мягкого протоплазматического тела корненожки. При многокамерном строении раковинки эта функция осуществляется весьма совершенно.
Внутренняя полость раковины заполнена цитоплазмой. Внутри раковинки помещается и ядерный аппарат. В зависимости от стадии размножения (о чем будет сказано несколько ниже) ядро может быть одно или их несколько. Из раковинки через устье наружу выдаются многочисленные очень длинные и тонкие псевдоподии, ветвящиеся и анастомозирующие между собой. Эти особые свойственные фораминиферам ложные ножки называют ризоподиями. Последние образуют вокруг раковинки тончайшую сеточку, общий диаметр которой обычно значительно превосходит диаметр раковинки (рис. 34). У тех видов фораминифер, у которых имеются поры, ризоподии выдаются наружу через поры.

Выполненные за последние годы с применением цейтраферной киносъемки исследования показали, что цитоплазма, входящая в состав ризоподии, находится в непрерывном движении. Вдоль ризоподии в центростремительном (к раковине) и центробежном (от раковины) направлениях довольно быстро текут токи цитоплазмы. По двум сторонам тонкой ризоподии цитоплазма как бы струится в противоположных направлениях. Механизм этого движения до сих пор остается не выясненным.
Размножение фораминифер происходит довольно сложно и у большинства видов связано с чередованием двух разных форм размножения и двух поколений. Одно из них бесполое, второе — половое. В настоящее время процессы эти изучены у многих видов фораминифер. Не вдаваясь в детали, рассмотрим их на каком-нибудь конкретном примере.

На рисунке 35 изображен жизненный цикл фораминиферы Elphidium crispa. Этот вид представляет собой типичную многокамерную фораминиферу со спирально закрученной раковинкой. Начнем рассмотрение цикла с многокамерной корненожки, обладающей маленькой зародышевой камерой в центре спирали (микросферическое поколение).
В цитоплазме корненожки первоначально имеется одно ядро. Бесполое размножение начинается с того, что ядро последовательно несколько раз делится, в результате чего образуется множество небольшого размера ядер (обычно несколько десятков, иногда свыше сотни). Затем вокруг каждого ядра обособляется участок цитоплазмы и все протоплазматическое тело корненожки распадается на множество (по числу ядер) одноядерных амебообразных зародышей, которые выходят через устье наружу. Сразу же вокруг амебовидного зародыша выделяется тонкая известковая раковинка, которая и явится первой (эмбриональной) камерой будущей многокамерной раковины. Таким образом, при бесполом размножении на первых стадиях своего развития корненожка является однокамерной. Однако очень скоро к этой первой камере начинают добавляться следующие. Происходит это так: из устья сразу выступает наружу некоторое количество цитоплазмы, которая тотчас же выделяет раковинку. Затем наступает пауза, в течение которой простейшее усиленно питается и масса протоплазмы его увеличивается внутри раковины. Затем вновь часть цитоплазмы выступает из устья и вокруг нее образуется очередная известковая камера. Этот процесс повторяется несколько раз: возникают все новые и новые камеры, пока раковина не достигнет характерных для данного вида размеров. Таким образом, развитие и рост раковины носит ступенчатый характер. Размеры и взаимное положение камер определяется тем, какое количество протоплазмы выступает из устья и как эта протоплазма располагается по отношению к предшествующим камерам.
Мы начали рассмотрение жизненного цикла Elphidium с раковинки, обладавшей очень маленькой эмбриональной камерой. В результате бесполого размножения получается раковинка, эмбриональная камера которой значительно крупнее, чем у той особи, которая приступала к бесполому размножению. В результате бесполого размножения получаются особи макросферического поколения, которые существенно отличаются от дающего им начало микросферического поколения. В данном случае потомство оказывается не вполне похожим на родителей.
Они развиваются в результате полового размножения макросферического поколения. Происходит это следующим образом. Как и при бесполом размножении, половой процесс начинается с деления ядра. Количество образующихся при этом ядер значительно больше, чем при бесполом размножении. Вокруг каждого ядра обособляется небольшой участок цитоплазмы, и таким путем формируется огромное количество (тысячи) одноядерных клеток. Каждая из них снабжена двумя жгутиками, благодаря движению которых клетки активно и быстро плавают. Эти клетки являются половыми клетками (гаметами). Они сливаются друг с другом попарно, причем слияние затрагивает не только цитоплазму, но и ядра. Этот процесс слияния гамет и есть половой процесс. Образующаяся в результате слияния гамет (оплодотворения) клетка носит название зиготы. Она дает начало новому микросферическому поколению фораминиферы. Вокруг зиготы сразу же по ее образовании выделяется известковая раковина — первая (эмбриональная) камера. Затем процесс развития и роста раковины, сопровождающийся увеличением числа камер, осуществляется по тому же типу, как и при бесполом размножении. Раковина получается микросферической потому, что размер зиготы, выделяющей эмбриональную камеру, во много раз меньше одноядерных амебовидных зародышей, образующихся при бесполом размножении. В дальнейшем микросферическое поколение приступит к бесполому размножению и вновь даст начало макросферическим формам.
На примере жизненного цикла фораминифер мы встречаемся с интересным биологическим явлением закономерного чередования двух форм размножения — бесполого и полового, сопровождающегося чередованием двух поколений — микросферического (развивается из зиготы в результате оплодотворения) и макросферического (развивается из одноядерных амебоидных зародышей в результате бесполого размножения).
Отметим еще одну интересную особенность полового процесса фораминифер. Известно, что у большинства животных организмов половые клетки (гаметы) бывают двух категорий. С одной стороны, это крупные, богатые протоплазмой и запасными питательными веществами неподвижные яйцевые (женские) клетки, а с другой — мелкие подвижные сперматозоиды (мужские половые клетки). Подвижность сперматозоидов обычно связана с наличием у них активно двигающегося нитевидного хвостового отдела. У фораминифер, как мы видели, морфологических (структурных) различий между половыми клетками нет. По строению своему все они одинаковы и благодаря наличию жгутиков обладают подвижностью. Здесь нет еще структурных различий, которые позволили бы различать мужские и женские гаметы. Такая форма полового процесса является исходной, примитивной.
Как уже было сказано, огромное большинство современных видов фораминифер являются придонными (бентическими) организмами, встречающимися в морях всех широт от прибрежной зоны вплоть до самых больших глубин мирового океана. Изучение распределения корненожек в океане показало, что оно зависит от ряда факторов внешней среды — от температуры, глубины, солености. Для каждой из зон типичны свои виды фораминифер. Видовой состав фораминифер может служить хорошим показателем условий среды обитания.

Широко распространенные в современных морях и океанах фораминиферы были богато представлены и в прежние геологические периоды начиная с самых древних кембрийских отложений. Известковые раковинки после размножения или смерти корненожки опускаются на дно водоема, где входят в состав отлагающегося на дне ила. Процесс этот совершается десятки и сотни миллионов лет; в результате на дне океана образуются мощные отложения, в состав которых входит несметное количество раковинок корненожек. При горообразовательных процессах, которые совершались и совершаются в земной коре, как известно, некоторые области дна океана поднимаются и становятся сушей, суша опускается и становится дном океана. Большая часть современной суши в различные геологические периоды была дном океана. Это относится в полной мере и к территории Советского Союза (за исключением немногих северных районов нашей страны: Кольский полуостров, большая часть Карелии и некоторые другие). Морские донные отложения на суше превращаются в горные осадочные породы. Во всех морских осадочных породах присутствуют раковинки корненожек. Некоторые же отложения, как например меловые, в основной своей массе состоят из раковин корненожек. Столь широкое распространение фораминифер в морских осадочных породах имеет большое значение для геологических работ, и в частности для геологической разведки. Фораминиферы, как и все организмы, не оставались неизменными. В течение геологической истории пашей планеты происходила эволюция органического мира. Изменялись и фораминиферы. Для разных геологических периодов истории Земли характерны свои виды, роды и семейства фораминифер. Известно, что по остаткам организмов в горных породах (окаменелостям, отпечаткам и т. п.) можно определить геологический возраст этих пород. Для этой цели могут быть использованы и фораминиферы. Как ископаемые они благодаря своим микроскопическим размерам представляют очень большие преимущества, так как могут быть обнаружены в очень небольших количествах горной породы. При геологической разведке полезных ископаемых (в особенности при разведке нефти) широко используется метод бурения. При этом получается колонка породы небольшого диаметра, охватывающая все слои, через которые прошел бур. Если эти слои представляют собой морские осадочные породы, то в них при микроскопическом анализе всегда обнаруживаются фораминиферы. Ввиду большой практической важности вопрос о приуроченности определенных видов фораминифер к тем или иным осадочным породам известкового возраста разработан с большой степенью точности.
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева . 1970 .
Читайте также: