
Взаимоотношения растений и паразита
Обновлено: 23.04.2024
Зрительные сигналы. Такие сигналы воспринимаются органами зрения. Примеры зрительных сигналов: яркая окраска венчика цветка, привлекающая опылителя; оранжевые пятна на брюшке жерлянки, предупреждающие о горьком вкусе; пестрая окраска ос и шмелей, говорящая об их защищенности. Большую роль во взаимоотношениях растений и насекомых-опылителей играет не только окраска, но и величина цветка или всего соцветия. К примеру, у растений из семейства сложноцветные или зонтичные сигнал подает не один цветок, а все соцветие, которое видно насекомому с большого расстояния. К зрительным сигналам относятся угрожающие позы и знаки животных (вздыбленная шерсть, оскал).
Химические сигналы. Этот способ взаимного оповещения организмов очень широко распространен в природе. Сигнальные вещества животных называются феромонами. Животные выделяют феромоны в окружающую среду и таким путем влияют на поведение особей того же вида. Эти вещества действуют при очень низких концентрациях, так как обладают сильным запахом. Различают несколько видов феромонов.
Половые феромоны облегчают встречу особей одного вида для спаривания, они важны для всех животных. Общеизвестны специфические феромоны для мечения территории, которые содержатся в моче собак, волков, представителей кошачьих и др.
Агрегационные феромоны помогают животным собраться вместе в большую группу (например, с их помощью собирается вместе население муравейника).
Выделение феромонов тревоги стимулирует уход особей с опасного участка территории (такие феромоны обнаружены у рыб, ящериц).
В настоящее время наиболее изучены феромоны насекомых. Причем оказалось возможным синтезировать многие сигнальные вещества и использовать их в качестве приманок. Специальные ловушки, внутри которых находятся источники выделения половых и агрегационных феромонов, позволяют заманивать и уничтожать вредителей сада или леса экологически безопасным способом, без применения гербицидов.
Большую роль во взаимоотношениях растений и насекомых играют запахи, привлекающие опылителей или, напротив, отпугивающие фитофагов.

1. Какие сигналы используют организмы при взаимоотношениях?
2. Как используются знания о феромонах в сельском хозяйстве?
2. Что такое аллелопатия и какова ее роль в природе?
Звуковые сигналы используют для управления поведением птиц. Трансляция записанного на магнитофон сигнала тревоги или крика хищной птицы помогает отпугнуть птиц от полей, аэродромов или других мест, где их присутствие нежелательно. Запись голоса курицы-наседки может улучшить аппетит инкубаторских цыплят.
Home Фитопатология Основы иммунитета Взаимоотношения растении с их паразитами
Взаимоотношения растении с их паразитами
В отличие от других хозяйственно-ценных признаков растений (таких, как урожайность или скороспелость) на проявление устойчивости влияют не только генетические особенности растения и условия выращивания, но и генетические особенности паразита, вызывающего заболевание. Выращивание на больших площадях устойчивого сорта часто вызывает накопление таких форм паразита, которые способны поражать его, подобно тому как широкое применение в медицине антибиотиков вызвало накопление форм патогенных бактерий, не чувствительных к этим антибиотикам.
Причины потери устойчивости сортов. Большинство фитопатогенных организмов — грибов, бактерий и вирусов — размножается в природе бесполым способом — делением клеток, образованием различных спор и т. д. При таком размножении ядра в клетках потомков возникают в результате деления ядер в клетках родителей и генетически не отличаются от родительских. Генетически однородное вегетативное потомство вирусной частицы, бактериальной клетки или грибной споры называется клоном. Вследствие того что в благоприятных условиях бесполое размножение микроорганизмов происходит очень быстро, клоны могут достигать громадных размеров и состоять из миллионов, миллиардов и даже триллионов клеток. Клоны грибов, споры которых распространяются воздушными течениями (ржавчинных, мучнисторосяных), могут занимать огромные географические районы. Те клоны, которые не способны заражать какой-либо сорт растения-хозяина, не обладают вирулентностью к этому сорту. Клоны, приобретшие способность заражать ранее устойчивый сорт, называют вирулентными.
Как видно из таблицы, сорт, не имеющий генов устойчивости, поражается любым клоном паразита, даже не имеющим генов вирулентности. Такой сорт называется универсально-восприимчивым.
Как же возникают и накапливаются гены вирулентности? Все организмы существуют в природе в виде популяций, т. е. групп особей одного вида, занимающих определенную территорию и способных скрещиваться друг с другом. В процессе размножения клона в отдельных клетках происходят генные мутации — случайные изменения в молекулах нуклеиновой кислоты, приводящие к изменению в строении белка, образование которого управляется измененным участком нуклеиновой кислоты (геном). Частота таких мутаций очень мала — по большинству генов не выше одной на десять миллионов ядер, однако вследствие огромного размера клонов микроорганизмов подобные мутации всегда в них присутствуют. Потомство мутантной клетки будет отличаться от исходного клона, т. е. образует новый клон. Вследствие этого популяция всегда неоднородна по многим генам. Такая неоднородность еще больше усиливается в популяциях организмов, имеющих половой процесс, ибо если скрещиваются клоны, имеющие мутации в разных генах, то в потомстве могут возникать формы, как сочетающие обе мутации, так и не имеющие ни одной.

Кроме мутаций, происходящих внутри популяции, вирулентные клоны могут попадать в нее из других популяций паразита.
Таким образом, на скорость возникновения вирулентных клонов влияют следующие факторы:
1) частота мутирования генов вирулентности;
2) плотность популяции;
3) наличие процессов комбинации генов (рекомбинации);
4) возможность миграции из других популяций. Рассмотрим эти факторы.
Частота мутирования, как отмечалось выше, невелика, однако она может быть значительно повышена воздействиями, которые повреждают нуклеиновые кислоты. Мутагенным действием обладают излучения, длина волны которых ниже длины волн видимого света (ультрафиолетовое, рентгеновское, гамма-излучения и др.), а также различные химические вещества, в том числе некоторые пестициды, широко используемые в сельском хозяйстве.
Плотность популяции имеет огромное значение в изменчивости, ибо вероятность возникновения редких мутаций тем выше, чем больше особей в популяции. Объем популяции паразитов тем выше, чем больше площадь, занимаемая восприимчивым сортом, чем меньше полей в севообороте и чем более благоприятны для развития болезни погодные условия.
Наличие рекомбинации генов резко повышает приспособительные возможности популяции. Если, например, сорт имеет два гена устойчивости, то согласно правилу Флора, чтобы поразить этот сорт, паразит должен иметь два гена вирулентности. Частота возникновения мутаций одновременно по двум генам равна произведению частот мутаций по каждому гену. Если частота мутирования по каждому гену составляет одно ядро на миллион, то частота одновременного мутирования по двум генам будет равна одному ядру на триллион, т. е. такое событие маловероятно даже в очень больших популяциях. Следовательно, вторая мутация может возникнуть лишь в потомстве клона, имеющего первую. Однако, поскольку концентрация мутантных клонов низкая, вероятность возникновения в них второй мутации также очень мала. Если же в популяции паразита протекает половой процесс, то становится возможным возникновение гибридов между двумя особями, несущими различные мутации вирулентности, и объединение этих мутаций в одном гибриде. Поэтому все мероприятия, снижающие способность паразита к гибридизации, замедляют процессы его изменчивости. Эффективным мероприятием является уничтожение промежуточных хозяев ржавчинных грибов — барбариса, крушины и др.
Кроме полового процесса, известны и другие способы обмена генетического материала между разными особями микроорганизмов. У многих грибов возможно слияние гиф растущих рядом особей (анастомозы). Вследствие этого ядра из одного мицелия через поры в перегородках клеток могут мигрировать в другой. Образуются клетки, содержащие ядра разных типов и объединяющие поэтому свойства обоих мицелиев (гетерокарионы). Гетерокариоз, или разноядерность, широко распространен среди грибов. У некоторых грибов гетерокариоз возникает вскоре после освобождения спор. Конидии возбудителя спорыньи выделяются с каплей нектара, в которой и прорастают, образуя при этом анастомозы и гетерокариотичные клетки. У некоторых грибов анастомозы возникают перед внедрением ростков спор в ткань растения-хозяина.
Возможности миграции вирулентных клонов также играют большую роль в потере сортами устойчивости. В качестве примера укажем, что вирулентные клоны часто возникают у возбудителей фитофтороза и рака картофеля, однако в первом случае они широко распространяются, а во втором сохраняются в виде локальных очагов. Это объясняется тем, что споры возбудителя фитофтороза распространяются по воздуху (ветром и дождевыми брызгами), а возбудителя рака — через почву; кроме того, клубни картофеля, зараженные фитофторозом, свободно перевозятся из одного места в другое, тогда как клубни из полей, зараженных раком, запрещается перевозить в другие места, так как рак — карантинный объект.
Методы изучения вирулентности. Вирулентность фитопатогенных организмов изучают с помощью заражения определенного набора сортов сельскохозяйственных растений, называемых растениями-дифференциаторами. Чаще всего используют два типа наборов дифференциаторов.
Эмпирический набор включает набор сортов, созданный на основе полевых наблюдений. Качество такого набора определяется тем, насколько хорошо с его помощью можно разделить (дифференцировать) популяцию паразита на клоны с различной вирулентностью. Например, универсально восприимчивый сорт не способен дифференцировать популяцию, поскольку он поражается всеми клонами. Очень устойчивый сорт также не будет дифференцировать популяцию вследствие устойчивости ко всем клонам. Дифференцируют популяцию паразита сорта, для которых часть клонов патогена вирулентна, а часть — авирулентна.

Например, в стандартный набор дифференциаторов возбудителя стеблевой ржавчины пшеницы входит 12 сортов. Среди них сорт Литтл Клаб поражается почти всеми клонами, собранными из многих стран, а сорт Капли устойчив почти ко всем клонам. Эти сорта служат в качестве контроля. Остальные 10 сортов поражаются одними клонами и устойчивы к другим, следовательно, способны дифференцировать популяцию. Клоны, обладающие одинаковой вирулентностью по отношению к набору сортов-дифференциаторов, объединяются в физиологические, расы. Если два клона не отличаются по вирулентности на наборе дифференциаторов, но имеют различную вирулентность по отношению к каким-либо сортам, не входящим в стандартный набор, их обозначают как биотипы внутри одной расы.
Различным расам дают цифровые обозначения. Наиболее простой способ обозначения — присвоение порядкового номера по мере обнаружения. Например, первый клон возбудителя стеблевой ржавчины, обнаруженный с помощью стандартного набора дифференциаторов, оказался вирулентным к шести сортам из 12, а второй — к трем. Все клоны, ведущие себя, как первый, получили название расы 1, а клоны, вирулентные, как второй,— расы 2. Сейчас на этом наборе описано свыше 300 рас, многие из которых разделены на ряд биотипов. Данный метод неудобен тем, что название расы ничего не говорит о степени ее вирулентности. Необходимо в каждом случае смотреть расшифровку номеров, опубликованную в специальных руководствах.
Гораздо больше информации можно получить, используя систему классификации рас, предложенную американским фитопатологом Хабгутом. Растения-дифференциаторы, обозначенные порядковыми номерами, записывают на бумаге линейно справа налево. Под ними выписывают реакцию на заражение данным клоном по двоичной системе (0 — устойчивость, 1 — восприимчивость). Значение каждой реакции умножают на 2n, где n — номер дифференциатора. Сумма произведений составляет номер расы. Например:

Чтобы по номеру расы определить, какие дифференциаторы восприимчивы к ней, надо 2 возводить в максимальную степень и полученное число вычесть из номера расы.
Остаток также возводится в максимальную степень и т. д. Например, раса 22 не поражает дифференциаторы №№ 6 и 5, ибо 25 больше 22, но поражает дифференциатор № 4, так как 24=16. Разность между 22 и 16 равна 6, следовательно, второй восприимчивый дифференциатор может быть № 2 (22=4), но не № 3, ибо 23 больше 6. Разность между шестью и четырьмя равна 2 (21), следовательно, эта раса поражает также дифференциатор № 1. Поскольку разница равна 0, дифференциатор № 0 устойчив.
Таким способом М. М. Левитин и О. С. Афанасенко впервые обозначили расы возбудителя сетчатого гельминтоспориоза ячменя, встречающиеся на территории СССР.
Б.М. Миркин, Л.Г. Наумова
Основы общей экологии
Учебное пособие. М.: Университетская книга, 2005.
Паразиты – это организмы, которые питаются за счет организма‑хозяина. Это очень разнообразная группа организмов (животные, растения, грибы, бактерии), которую изучает специальная наука – паразитология.
Различаются следующие группы паразитов.
Биотрофы и некротрофы. Биотрофы всю жизнь питаются за счет живого хозяина, а некротрофы губят его (или часть его тела, например лист растения) и завершают свою биографию как рассматриваемые далее детритофаги.
Микропаразиты и макропаразиты. Различаются по размерам. К микропаразитам относятся вирусы, бактерии, микроскопические грибы и простейшие, к макропаразитам – все прочие.
Истинные паразиты и паразитоиды. Истинные паразиты всю свою жизнь питаются за счет организма‑хозяина (или нескольких хозяев, если в течение жизненного цикла переселяются из организма одного вида в другой). Паразитоиды (как правило, насекомые) на определенных стадиях жизненного цикла ведут свободный образ жизни (питаются как фитофаги или зоофаги). После этого они откладывают яйца в тело организма‑хозяина, в котором паразитируют личинки. Паразитоиды представляют переход к хищникам. Насекомые‑паразитоиды используются для биологического метода контроля насекомых‑вредителей в сельском хозяйстве (трихограмма, теленомус‑наездник и др.).
Под микроскопом он открыл, что на блохе
Живет блоху кусающая блошка;
На блошке той – блошинка‑крошка,
В блошинку же вонзает зуб сердито
Блошиночка… и так ad infinitum.
В этом случае из паразитов формируется пищевая цепь.
Различаются группы паразитов и по сложности жизненного цикла. Одни виды паразитов передаются при непосредственном контакте особей хозяина (например вирусы и бактерии, вызывающие болезни человека). Другие паразиты перед заражением основного хозяина, в теле которого они образуют потомство, проходят через один или несколько видов промежуточных хозяев (например широкий лентец – паразит человека, но в течение жизненного цикла он проходит через стадии жизни в рачках‑циклопах и рыбах).
Среди полупаразитов есть виды, наносящие вред хозяйству человека. Например погремки (Rhinanthus) при доминировании в луговых сообществах становятся основными продуцентами органического вещества. В этом случае за счет потери энергии при переходе с одного трофического уровня на другой (см. 10.4) биологическая продукция травостоя снижается в 2‑3 раза. Причем погремок не поедается сельскохозяйственными животными ни на пастбище, ни в сене.
В процессе длительной коэволюции (взаимоприспособления) паразитов и хозяев вырабатываются специальные механизмы, которые позволяют им устойчиво сосуществовать.
Защитные реакции хозяев могут быть следующими:
– иммунный ответ организма, т.е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов;
– сбрасывание зараженных частей (это особенно характерно для растений‑хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги;
– выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при паразитировании тли);
– уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой добычей хищников, которые таким образом снижают долю зараженных особей в популяции;
– формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям.
Ситуация изменяется в антропогенных экосистемах, особенно в сельскохозяйственных, где заражение скота паразитами может привести к гибели многих животных. Представляют опасность взаимоотношения паразитов и человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом (при заражении простейшим – лямблией), болезнями бактериальной и вирусной природы.
1. Чем паразиты отличаются от хищников?
2. Расскажите о разнообразии паразитов.
3. Какие защитные реакции против паразитов вырабатываются у хозяев?
Отношения типа хищник-жертва, паразит-хозяин – это прямые пищевые связи, которые для одного из партнеров имеют отрицательные, а для другого – положительные последствия. По существу, к этому типу экологических взаимодействий можно отнести все варианты пищевых связей. Хотя пасущуюся на лугу корову и дятла, добывающего личинок из-под коры дерева, обычно не называют хищниками, тот тип взаимоотношений, в которые они вступают с организмами, служащими им пищей, имеет много общего с отношениями хищника и его жертв. Поэтому в западной литературе связь хищник– жертва понимается обычно в широком смысле, включая все формы добывания пищи. Они, в свою очередь, подразделяются на несколько категорий: 1) истинное хищничество, или хищничество в узком смысле слова; 2) паразитизм; 3) собирательство и 4) пастьба.
Хищниками обычно называют животных, питающихся другими животными, которых они ловят и умерщвляют. Для хищников характерно специальное охотничье поведение.
Добыча жертвы требует от них значительных затрат энергии на поиск, погоню, захват, преодоление сопротивления жертв.
Если размеры жертв намного меньше размеров питающихся ими животных, численность объектов питания высока и сами они легкодоступны – в этом случае деятельность плотоядного вида превращается в поиск и простой сбор добычи и называется собирательство.
К собирательству можно отнести фильтрационное питание водных животных, седиментацию, или осаждение водной взвеси, сбор пищи илоедами или дождевыми червями. К нему же примыкает так называемое хищничество растений. У многих растений при недостатке азота в питании развиты способы улавливания и фиксации прилетающих к ним насекомых и переваривание белков их тел протеолитическими ферментами (пузырчатки, росянки, непентесы, венерина мухоловка и др.).
По способу овладения пищевыми объектами собирательство приближается к типичной пастьбе фитофагов. Специфика пастьбы заключается в поедании неподвижного корма, находящегося в относительном изобилии, на поиски которого не приходится тратить много усилий. С экологической точки зрения такой способ питания характерен как для стада копытных на лугу, так и для листогрызущих гусениц в кроне дерева или личинок божьих коровок в колониях тлей.
Таким образом, несмотря на экологическую специфичность разных способов питания, они связаны между собой всевозможными переходами, а иногда встречаются вместе у одного и того же вида. Такие же переходы можно встретить в природе между типичным хищничеством и паразитизмом.
Паразитизм такая форма связей между видами, при которой организм-потребитель использует живого хозяина не только как источник пищи, но и как место постоянного или временного обитания (рис. 85). По существу, типичный паразитический характер имеют связи насекомых-вредителей с растениями. Паразиты обычно намного мельче своего хозяина.
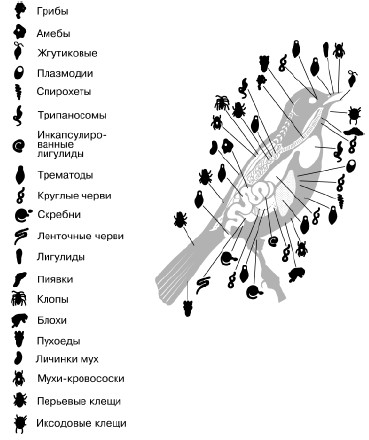
Рис. 85. Паразиты певчих птиц
Во взаимоотношениях хищник-жертва, паразит-хозяин наиболее ярко проявляется эволюционная и экологическая роль пищевых связей организмов. Хищничество, связанное с активным поиском и энергичными способами овладения сопротивляющейся и убегающей добычей, ведет к выработке разнообразных экологических адаптаций как у жертв, так и у их потребителей. При активном способе защиты от врагов естественный отбор способствует развитию у жертв органов чувств, быстроты реакции, скорости бега, инстинктов обманного поведения, что сопровождается совершенствованием нервной системы и ведет к прогрессивной эволюции группы.
При пассивном способе защиты развиваются покровительственная окраска, твердые панцири, шипы, иглы, инстинкты затаивания, использования недоступных хищникам убежищ и т. п. Некоторые из этих способов защиты характерны не только для малоподвижных или сидячих видов, но и для активно спасающихся от врагов животных.
В свою очередь, трудность обнаружения и поимки жертв способствует у хищников отбору на лучшее развитие органов чувств (зоркость, тонкий слух, чутье и т. д.), на более быструю реакцию на добычу, выносливость при преследовании и т. п. Таким образом, экологические связи хищников и жертв направляют ход эволюции сопряженных видов.
Хищники обладают обычно широким спектром питания. Добыча жертв требует много сил и энергии. Специализация ставила бы хищников в сильную зависимость от численности определенного вида жертв. Поэтому большинство видов, ведущих хищный образ жизни, способно переключаться с одной добычи на другую, особенно на ту, которая в данный период более доступна и многочисленна. Правда, у многих хищников есть предпочитаемые виды жертв, которых они добывают чаще, чем других. Эта избирательность может быть обусловлена разными причинами. Во-первых, хищник активно выбирает наиболее полноценную в кормовом отношении пищу. Например, нырковые утки и сиговые рыбы в северных водоемах выбирают среди водных беспозвоночных преимущественно личинок комаров-хирономид (мотыль), и их желудки иногда бывают наполнены мотылем, несмотря на наличие в водоеме и другого корма.
Характер пищи может быть обусловлен также пассивной избирательностью: хищник в первую очередь поедает такие корма, к добыче которых он наиболее приспособлен. Так, многие воробьиные питаются всеми насекомыми, обитающими открыто на поверхности почвы, на траве, листьях и т. п., но не поедают почвенных беспозвоночных, для добычи которых нужны специальные приспособления. Наконец, третьей причиной пищевой избирательности хищников может быть активное переключение на наиболее массовую добычу, появление которой стимулирует охотничье поведение. При высокой численности леммингов даже соколы-сапсаны, основной способ охоты которых – добывание птиц в воздухе, начинают охотиться на леммингов, схватывая их с земли. Способность переключения с одного вида добычи на другой – одно из необходимых экологических приспособлений в жизни хищников.
Паразитизм, в отличие от хищничества, характеризуется более узкой специализацией видов. Поскольку хозяин обеспечивает паразиту не только пищу, но и микроклимат, защиту и т. п., то чем лучше приспособленность паразита к особенностям организма хозяина, тем вероятнее его успех в размножении и оставлении потомства.
Катастрофический вред от паразитов выражен преимущественно лишь в тех связях, которые еще не стабилизированы длительным ходом естественного отбора. Поэтому случайно завезенные вредители поражают сельскохозяйственные растения или животных часто намного сильнее, чем местные.
Среди многообразных форм паразитических отношений есть и такие, при которых гибель хозяина – обязательное следствие пребывания в нем паразита. Этот тип связей особенно распространен у насекомых, откладывающих свои яйца в яйца или личинки других. Такие насекомые получили название паразитоидов. Гибель хозяина обусловлена малым запасом в нем пищи, которой едва хватает на развитие одной или немногих личинок вида-потребителя.
Паразитизм, таким образом, связан всевозможными переходами с другими типами взаимоотношений.
Основная экологическая роль хищничества, паразитизма и других вариантов пищевых связей в сообществах заключается в том, что, последовательно питаясь друг другом, живые организмы создают условия для круговорота веществ, без которого невозможна жизнь. Вторая не менее важная роль этих отношений – взаимная регуляция численности видов.
В естественных экосистемах растения и поедающие их фитофаги приспосабливаются друг к другу таким образом, чтобы животные не могли быстро съесть все растения и остаться без пищи. Для замедления поедания животными у растений вырабатываются специальные адаптации (рис. 39). Они защищаются:
– вырабатывая горькие вещества, ухудшающие их вкус;
– выделяя резкие запахи, отпугивающие фитофагов;
– образуя колючки и низкие формы с прижатыми к земле листьями, малодоступными для пасущихся животных.
Однако растения защищаются от фитофагов весьма умеренно и лишь настолько, чтобы их съели не слишком быстро (но все-таки съели!). Полная защищенность растений от поедания может нанести вред не только фитофагам, но и самим растениям:
– если растения не будут съедены, то содержащиеся в них питательные вещества не возвратятся в почву, и это осложнит условия для роста других растений;
Однако, несмотря на эти защитные приспособления растений, при периодических вспышках плотности популяций фитофагов возможно чрезмерное выедание популяций растений. Результатом такого чрезмерного выедания растений становится подрыв кормовой базы фитофагов и снижение плотности их популяций.
Равновесие между хищниками и их жертвами у животных также поддерживается специальными механизмами, исключающими полное истребление жертв. Так, жертвы могут:
– убегать от хищника (в этом случае в результате соревнования повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей);
– распространяться группами, что делает их поиск и промысел для хищника более энергоемким;
– прятаться в укрытия;
Полной защиты от хищников всех особей в популяциях животных-жертв не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге – к катастрофе популяций жертв. В то же время, при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные), и ввиду резкого увеличения их численности подрывается кормовая база.
Защитные реакции хозяев могут быть следующими:
– повышение устойчивости к заражению паразитами (устойчивость культурных растений к паразитным грибам может быть повышена селекцией);
– сбрасывание зараженных частей (это особенно характерно для растений-хозяев, которые сбрасывают сильно зараженные листья);
– быстрый рост здоровых тканей взамен пораженных (это имеет место при поедании тканей растений тлями; некоторые сорта ржи способны за счет образования новых побегов уменьшить вред от паразитов, которые заселили часть побегов);
– уменьшение плотностипопуляций хозяев, что снижает вероятность распространения паразита и заражения им. Этому помогают хищники: зараженные животные менее подвижны и становятся для них более легкой добычей.
В естественных экосистемах экологическое равновесие между популяциями паразитов и их хозяев – нормальное явление. Ситуация изменяется в экосистемах, создаваемых человеком, особенно в сельскохозяйственных, где заражение паразитами может привести к существенному падежу скота. Паразиты представляют опасность и для человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом (при заражении простейшим – лямблией), болезнями бактериальной и вирусной природы. Ущерб человеку могут нанести и насекомые-паразиты (вши, клопы, клещи и др.).
1. Какие приспособления растений замедляют их потребление фитофагами?
2. За счет каких приспособлений регулируются взаимоотношения жертв и хищников?
Читайте также: