
Белок к которому крепится вирус
Обновлено: 17.04.2024
Структурные белки делятся на 2 группы:
1) капсидные белки, образующие капсид, т. е. футляр для нуклеиновой кислоты вируса (от лат. сарза — вместилище), и входящие в состав капсида геномные белки, и ферменты;
2) суперкапсидные белки, входящие в состав суперкапсида, т. е. наружной вирусной оболочки.
Просто организованные вирусы, представляющие собой нуклеокапсид, содержат только капсидные белки. Сложно организованные вирусы содержат капсидные и суперкапсидные белки.
В составе капсида некоторых вирусов (пикорнавирусы, паповавирусы, аденовирусы) содержатся белки, ковалентно связанные с вирусным геномом (геномные белки). Эти белки являются терминальными, т. е. соединенными с концом вирусной нуклеиновой кислоты. Функции их неразрывно связаны с функциями генома и их регуляцией.
У ряда сложно организованных вирусов в составе капсида имеются ферменты, осуществляющие транскрипцию и репликацию вирусного генома — РНК и ДНК (РНК- и ДНК-полимеразы), а также ферменты, модифицирующие концы иРНК. Если ферменты и геномные белки представлены единичными молекулами, то капсидные белки представлены множественными молекулами. Эти белки и формируют капсидную оболочку, в которую у сложно организованных вирусов вставлены молекулы белков с другими функциями.
Основным принципом строения капсидной оболочки вирусов является принцип субьединичности, т. е. построение капсидной оболочки из субъединиц-капсомеров, образованных идентичными полипептидными цепями. Правильно построенные белковые субъединицы — капсомеры возникают благодаря способности вирусных капсидных белков к самосборке. Самосборка объясняется тем, что упорядоченная структура — капсид имеет наименьшую свободную энергию по сравнению с неупорядоченными белковыми молекулами. Сборка капсидной оболочки из субъединиц запрограммирована в первичной структуре белка и происходит самопроизвольно или при взаимодействии с нуклеиновой кислотой.
Суперкапсидные белки. Гликопротеиды. Суперкапсидные белки, или пепломеры, располагаются в липопротеидной оболочке (суперкапсиде или пеплосе) сложно устроенных вирусов. Они либо пронизывают насквозь липидный бислой как, например, гликопротеиды альфа-вирусов (вируса леса Семлики), либо не доходят до внутренней поверхности. Эти белки являются типичными внутримембранными белками и имеют много общего с клеточными мембранными белками. Как и последние, суперкапсидные белки обычно гликозилированы. Углеводные цепочки прикреплены к молекуле полипептида в определенных участках. Гликозилирование осуществляют клеточные ферменты, поэтому один и тот же вирус, продуцируемый разными видами клеток, может иметь разные углеводные остатки: может варьировать как состав углеводов, так и длина углеводной цепочки и место прикрепления ее к полипептидному остову[18].

Основной функцией гликопротеидов является взаимодействие со специфическими рецепторами клеточной поверхности. Благодаря этим белкам осуществляется распознавание специфических клеточных рецепторов и прикрепление к ним вирусной частицы, т. е. адсорбция вируса на клетке. Поэтому гликопротеиды, выполняющие эту функцию, называют вирусными прикрепительными белками.
Другой функцией гликопротеидов является участие в слиянии вирусной и клеточной мембран, т. е. в событии, ведущем к проникновению вирусных частиц в клетку. Вирусные белки слияния ответственны за такие процессы, как гемолиз и слияние плазматических мембран соседних клеток, приводящие к образованию гигантских клеток, синци-тиев и симпластов.
Неструктурные белки. Неструктурные белки изучены гораздо хуже, чем структурные, поскольку их выделяют не из очищенных препаратов вирусов, а из зараженных клеток, и возникают трудности в их идентификации и очистке от клеточных белков.
К неструктурным белкам относятся:
1) предшественники вирусных белков, которые отличаются от других неструктурных белков нестабильностью в зараженной клетке в результате быстрого нарезания на структурные белки;
2) ферменты синтеза РНК и ДНК (РНК- и ДНК-26 полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома;
4) ферменты, модифицирующие вирусные белки, например протеиназы и протеинкиназы.
Однако многие неструктурные белки при ряде вирусных инфекций еще не идентифицированы и функции их не определены. Типы структурных и неструктурных белков просто и сложно устроенных вирусов и их функции показаны на схеме (рис.6).
  Белки просто устроенных вирусов |

 Предшественники структурных | |
 | Капсидные |
 Структурные |  Полимеразы |
| Геномные | |
  Ферменты, модифицирующие белки | Неструктурные |
| Функции не идентифицированы |
  Белки сложно устроенных вирусов |
 | Капсидные |  Предшественники структурных | |
  В составе капсида | Геномные |  Полимеразы | |
 | |||
 Структурные | Полимеразы |   Регуляторы синтеза РНК, ДНК | Неструктурные |
| В составе суперкапсида |    | Белки слияния | Функции не идентифицированы |
| Ферменты, модифицир- ующие белки | |||
| Прикрепительные |
Рис. 6. Типы структурных и неструктурных белков просто и сложно устроенных вирусов.
Липиды вирусов
Липиды обнаружены у сложно организованных вирусов и в основном находятся в составе липопротеиднои оболочки (суперкапсида), формируя ее липидной бислой, в который вставлены суперкапсидные белки.
Все сложно организованные РНК-содержащие вирусы имеют в своем составе значительное количество липидов (от 15 до 35% от сухого веса). Из ДНК-содержащих вирусов липиды содержат вирусы оспы, герпеса и гепатита В. Примерно 50—60% липидов в составе вирусов представлено фосфолипидами, 20—30% составляет холестерин.
Липидный компонент стабилизирует структуру вирусной частицы. Экстракция липидов органическими растворителями, обработка вирусной частицы детергентами или липазами приводит к деградации вирусной частицы и потере инфекционной активности[19].
Вирусы, содержащие липопротеидную мембрану, формируются путем почкования на плазматической мембране клеток (или на мембранах эндоплазматической сети с выходом во внутриклеточные вакуоли). Поэтому липопротеидная оболочка этих вирусов представляет собой мембрану клетки-хозяина, модифицированную за счет наличия на ее наружной поверхности вирусных супер-капсидных белков. Из этого следует, что состав липидов почкующихся вирусов близок к составу липидов клетки-хозяина. К почкующимся вирусам относятся крупные
РНК-содержащие вирусы: ортомиксовирусы, парамиксо
вирусы, рабдовирусы, тогавирусы, ретровирусы, бунья
вирусы, аренавирусы, коронавирусы.
В связи с клеточным происхождением липидов общий состав липидной фракции и содержание ее отдельных компонентов у одного и того же вируса могут существенно различаться в зависимости от клетки-хозяина, где происходила репродукция вируса. Наоборот, если разные почкующиеся вирусы репродуцировались в одних и тех же клетках, их липиды оказываются более или менее сходными[20].
У вирусов оспы и гепатита В липиды имеют иное происхождение, так как эти вирусы не почкуются через плазматическую мембрану. У вирусов оспы липиды не образуют дифференцированной оболочки. Обработка вируса осповакцины эфиром не приводит к потере инфекционной активности или каким-либо структурным изменениям вириона. Липиды вируса гепатита В и его НВз-антигена образуются путем инвагинации мембран эндоплазматической сети. Вирус герпеса формируется путем почкования через ядерную оболочку, поэтому в его составе есть липиды ядерной оболочки.
Углеводы вирусов
Углеводный компонент вирусов находится в составе гликопротеидов. Количество Сахаров в составе гликопротеидов может быть достаточно большим, достигая 10—13% от массы вириона. Химическая специфичность их полностью определяется клеточными ферментами, обеспечивающими перенос и присоединение соответствующих сахарных остатков. Обычными сахарными остатками, обнаруживаемыми в вирусных белках, являются фруктоза, сахароза, манноза, галактоза, нейраминовая кислота, глюкозамин. Таким образом, подобно липидам, углеводный компонент определяется клеткой-хозяином, благодаря чему один и тот же вирус, Взращенный в клетках разных видов, может значительно различаться по составу Сахаров в зависимости от специфичности клеточных гликозилтрансфераз.
Углеводный компонент гликопротеидов играет существенную роль в структуре и функции белка. Он является каркасом для локальных участков гликопротеида, обеспечивая сохранение конформации белковой молекулы, и обусловливает защиту молекулы от протеаз. Возможны и другие функции углеводов, пока достоверно не установленные[21].
Заключение
В итоге работы можем сделать следующие выводы.
Вирусы — это неклеточные формы жизни, различимые только под электронным микроскопом. Это внутриклеточные паразиты. За пределами клетки они не проявляют своих свойств и имеют кристаллическую форму.
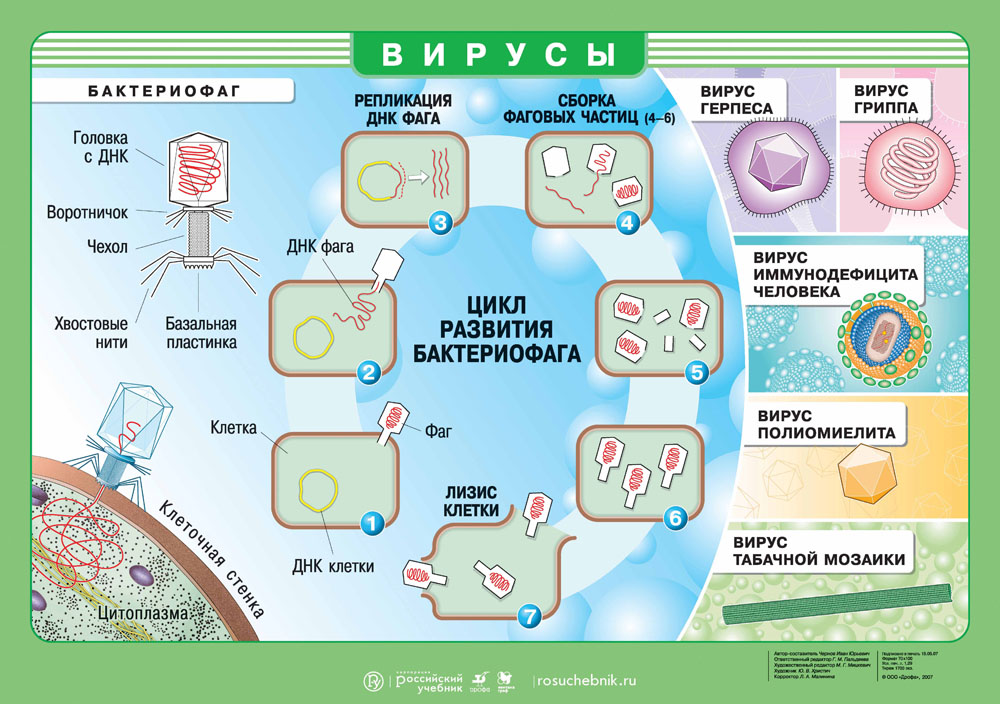
Наиболее просто организованные вирусы представляют собой нуклеопротеид, состоящий из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки (капсида). Сложные вирусы могут иметь дополнительную оболочку из липопротеина. Некоторые вирусы (бактериофаги) имеют аппарат транспортировки своего генома в бактерии, после проникновения в клетку капсид остается за пределами клетки. Тело бактериофага имеет сложное строение, оно содержит головку, хвостик (трубку, через которую геном проталкивается в клетку) и хвостовые отростки.
В клетку вирусы могут попасть вместе с пиноцитозными пузырьками или путем погружения части оболочки клетки с приклеившимся к ней вирусом в цитоплазму, а также путем растворения оболочки клетки.
Вирусы вносят в клетку свою генетическую информацию, и клетка начинает производить подобные вирусы.
Внутри клетки начинает синтезироваться ДНК или РНК вируса и образуется множество вирусов. В результате клетка гибнет, и вирусы выходят наружу, заражая новые клетки. Встроенный в геном клетки геном вируса может существовать в таком виде долгое время.
Вирусы вызывают табачную мозаику у растений, оспу, грипп, полиомиелит, гепатит, СПИД у человека. Наибольшую опасность в наше время представляет вирус СПИДа. Он попадает в организм человека при переливании крови, при половых контактах. Этот вирус поражает клетки организма, отвечающие за иммунитет. В результате человек оказывается беззащитным перед инфекционными болезнями и быстро погибает.
Вирусы, благодаря мутированию и способности быстро размножаться внутри клеток, становятся устойчивыми к действию лекарств, и это обстоятельство затрудняет лечение таких вирусных заболеваний, как грипп, гепатит и др.
Список литературы
1. Агол В.И. Биосинтез вирусных нуклеиновых кислот // Молекулярная биология: Структура и биосинтез нуклеиновых кислот / Под ред. А.С.Спирина. М.: 1990.
3. Борисов Л.Б. Микробиология, иммунология, вирусология. М.: МИА, 2005. – 736с.
4. Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 17-19
5. Вилляреал Л. Вирус: существо или вещество? // В мире науки. 2005. - №3.
6. Вирусология. Под редакцией Филдса Б., Найта Д., тт. 1–3, М.: Медицина, 1989- 492с.
7. Дяченко С. С., Синяк К. М., Дяченко Н. С. Патогенные вирусы человека. – Киев: Здоров'я, 1980. – 448 с.
9. Микробиология и иммунология. / Под ред. А.А. Воробьева. – М.: Медицина, 1999. – 464с.
10. Николаев А.Я. Биологическая химия. – М.: МИА, 2004. – 566 с.
11. Поздеев О. К. Медицинская микробиология. – Москва: ГЭОТАР – Мед., 2001.
12. Прозоркина Н. В., Рубашкина Л. А. П 78 Основы микробиологии, вирусологии и иммунологии: Учебное пособие для средних специальных медицинских учебных заведений. — Ростов нД: Феникс, 2002. -416с.
13. Фролов А. Ф., Шевченко Л. Ф., Широбоков В. П. Практическая вирусология. – Киев: Здоровье, 1989.
[1] Прозоркина Н. В., Рубашкина Л. А. П 78 Основы микробиологии, вирусологии и иммунологии: Учебное пособие для средних специальных медицинских учебных заведений. — Ростов нД: Феникс, 2002. -416с.
[2] Вилляреал Л. Вирус: существо или вещество? // В мире науки. 2005. - №3.
[4] Агол В.И. Биосинтез вирусных нуклеиновых кислот // Молекулярная биология: Структура и биосинтез нуклеиновых кислот / Под ред. А.С.Спирина. М.: 1990. С. 260-333.
[5] Микробиология и иммунология. / Под ред. А.А. Воробьева. – М.: Медицина, 1999. – 464с. С. 44-45.
[7] Николаев А.Я. Биологическая химия. – М.: МИА, 2004. – 566 с.
[8] Фролов А. Ф., Шевченко Л. Ф., Широбоков В. П. Практическая вирусология. – Киев: Здоровье, 1989. с. 217 – 225.
[9] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 17-19
[10] Вирусология. Под редакцией Филдса Б., Найта Д., тт. 1–3, М.: Медицина, 1989- 492с.
[11] Поздеев О. К. Медицинская микробиология. – Москва: ГЭОТАР – Мед., 2001. С. 459 – 465.
[12] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 17-19.
[13] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 19-22.
[14] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 19-22.
[15] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 19-22.
[16] Дяченко С. С., Синяк К. М., Дяченко Н. С. Патогенные вирусы человека. – Киев: Здоров'я, 1980. – 448 с.
[17] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 22-27
[18] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 22-27.
[19] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 28
[20] Борисов Л.Б. Микробиология, иммунология, вирусология. М.: МИА, 2005. – 736с., С. 603.
[21] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 29-30.
Мы кратко рассмотрели структуру вирионов гриппа и процесс кодирования вирусными РНК одного и более белков. Теперь обратимся к размножению вирусов гриппа.
Вирусы – это исключительно внутриклеточные паразиты, они не могут воспроизводиться вне клетки. Появление новых инфекционных частиц должно протекать внутри клетки. Войдя в клетки, вирусы начинают паразитировать в структуре хозяина для воспроизводства вирусного потомства. Все события, происходящие в инфицированной вирусом клетке, называются инфекционным циклом, или вирусной репликацией. Вирусологи искусственно разделяют инфекционный цикл на несколько стадий, чтобы проще было его изучить. Эти стадии включают прикрепление и вхождение вириона, трансляцию мРНК в белок, репликацию генома (создание большего количества РНК или ДНК), скопление новых частиц и выход частиц из клетки. Мы рассмотрим каждую из названных стадий, а затем обсудим, как вирус гриппа инфицирует нас и вызывает болезнь.
Остановимся на первой стадии – прикреплении вириона к клетке. Возьмем обычную клетку, с которой, я уверен, знаком каждый. Но не будет лишним повториться.
Рисунок 10.
Слева показан клеточный белок, прикрепленный к цитоплазматической мембране. Снизу изображена внутренняя часть клетки – цитоплазма. Часть белка пересекает мембрану, и также видны части цитоплазматической и внеклеточной сторон. Сферы – это сахара, прикрепленные ко множеству белков (белок + сахар = гликопротеин). Сиаловая кислота всегда является последним сахаром в цепи, которая прикрепляется к белку. Справа показана химическая структура сиаловой кислоты; следующий сахар (справа) – галактоза. Вирионы гриппа прикрепляются к клеткам, когда HA захватывает очень маленькую сиаловую кислоту.
Сахар в сравнении с HA в действительности очень мал – он подходит к небольшому кармашку на острие шипа. На рис. 11 изображена молекулярная модель, демонстрирующая, как HA прикрепляется к аналогу сиаловой кислоты. Шаровидный конец HA показан в верхней части рис. 11. Маленькие красные и белые сферы показывают места будущего прикрепления сиаловой кислоты – кармашки на верхнем конце HA.
Рисунок 13.
Процесс вхождения вируса гриппа в клетки – наиболее понятный из всех известных механизмов проникновения вирусов. После прикрепления вириона к сиаловой кислоте, имеющей рецепторы на поверхности клетки, вирус-рецепторный комплекс входит в клетки посредством эндоцитоза – процесса, в ходе которого клетки обычно берут молекулы из внеклеточной жидкости. Пока эндосомальные везикулы, содержащие частицы вируса, движутся к ядру клетки, их pH понижается. Это изменение в pH завершает клеточный канал, закачивающий протоны (H+) в везикулу. Когда эндосомальная pH достигает 5,0, вирусный белок HA подвергается конформационной перестройке. Это изменение подвергает пептид слияния на HA короткой гидрофобной последовательности, вставляющей его в эндосомальную мембрану, которая от этого сливается с вирусной оболочкой. Когда это происходит, вирусные РНК проникают в цитоплазму. Затем они транспортируются в ядро клетки, где начинается размножение.
В вирионе гриппа вирусные РНК не оголены, поскольку к ним прикреплены вирусные белки, включая белок M1. Этот белок образует оболочку, лежащую в основе липидной мембраны вириона. К сожалению, если вирусные РНК прикрепляются к белку M1, когда выходят из вириона, они не могут достичь ядра. Для решения этой проблемы вирион гриппа имеет в своей мембране несколько копий белка, называемого M2. Этот вирусный белок образует канал в мембране, который активно закачивает протоны из эндосомы во внутреннюю часть вириона. Данные протоны понижают pH внутри вириона, освобождая вирусные РНК от M1. В этом случае РНК могут войти в ядро.
Ионный канал M2, являющийся мишенью для противовирусных адамантанов, показан на рис. 14. Эти образования забивают канал и не позволяют протонам закачиваться в вирион. В присутствии адамантанов вирусным РНК нужно прикрепляться к M1, отчего они не могут достичь ядра. Поэтому вирусная репликация подавляется. Устойчивость к адамантанам возможна при изменении в аминокислотах, прокладывающих канал M2. Такие изменения не дают лекарству забить канал.
Рисунок 15.
На рис. 15 сферический конец белка HA, который прикрепляется к рецепторам клетки, изображен сверху, а вирусная мембрана – снизу. Для наглядности помечен только сайт расщепления HA. Нерасщепленная форма белка называется HA0; после расщепления клеточным ферментом образуются два белка, называемые HA1 (синий) и HA2 (красный). Две субъединицы остаются вместе на поверхности вирусной частицы. Новая амино(N)-концевая область HA2, образовавшегося при расщеплении, содержит последовательность гидрофобных аминокислот, называемых пептидом слияния. Во время вхождения вируса гриппа в клетки пептид слияния вставляется в эндосомальную мембрану и вызывает слияние вирусной и клеточной оболочек. Следовательно, вирусные РНК гриппа могут войти в цитоплазму. Процесс слияния описан в предыдущем параграфе.
Если белок HA не расщеплен, чтобы образовать HA1 и HA2, слияния не происходит. Поэтому вирусы гриппа с нерасщепленным HA не заразны. Расщепление вирусного HA происходит после того, как вновь синтезированные вирионы выходят из клетки. Вирусы гриппа эффективно размножаются в яйцах из-за присутствия протеазы в аллантоисной жидкости, способной расщеплять HA. Однако репликация многих штаммов вируса гриппа в культурах клеток требует добавления в среду соответствующей протеазы (как привило, трипсина).
У людей репликация вируса гриппа ограничена дыхательными путями, потому что это единственный участок, где производится протеаза, расщепляющая HA. Однако белок HA высокопатогенных штаммов H5 и H7 вируса птичьего гриппа может расщепляться протеазами, производимыми во многих различных тканях. В результате эти вирусы способны размножаться во многих органах птиц, включая селезенку, печень, легкие, почки и мозг. Это свойство объясняет способность штаммов H5N1 вируса птичьего гриппа размножаться вне дыхательных путей человека.
Подобно белкам HA высокопатогенных вирусов H5 и H7, HA штамма вируса гриппа 1918 г. может также расщепляться повсеместно присутствующими клеточными протеазами. Следовательно, вирус может размножаться в культурах клеток без добавления трипсина.
Белки HA H5 и H7 имеют множество основных аминокислотных остатков на сайте расщепления HA1-HA2, которые делают возможным расщепление посредством широко представленных протеаз. Но HA вируса гриппа H1 1918 г. не обладал этим свойством. Также N1 1918 г. не был в состоянии привлечь протеазы, которые расщепляли бы HA, т. е. не было механизма, позволяющего штамму A/WSN/33 вируса гриппа размножаться в клетках без трипсина. Понимание того, как белок HA вируса H1 1918 г. мог быть расщеплен протеазами – основной в понимании высокой патогенности данного штамма.
Рекомендуемая литература.
Chaipan, C., Kobasa, D., Bertram, S., Glowacka, I., Steffen, I., Solomon Tsegaye, T., Takeda, M., Bugge, T., Kim, S., Park, Y., Marzi, A., &Pohlmann, S. (2009). Proteolytic Activation of the 1918 Influenza Virus Hemagglutinin Journal of Virology, 83 (7), 3200-3211 DOI:10.1128/JVI.02205-08.
Главная задача биологии — это развитие представлений у человека о живых организмах, о многообразии видов, обо всех закономерностях развития живых существ, а также об их взаимодействии с окружающей природой. Предмет основы безопасности жизнедеятельности (ОБЖ) позволяет получить знания и умения, которые помогут сохранить жизнь и здоровье в опасных ситуациях. Эти ситуации всегда возникают неожиданно, но, тем не менее, большинство из них предсказуемы и к ним можно подготовиться заранее. ОБЖ учит нас предвидеть возможные опасности и минимизировать потери от той или иной ситуации. Сегодня мы сталкиваемся с новым видом вирусной опасности COVID-19,о котором поговорим с точки зрения биологии и ОБЖ.
Что такое вирус?
Вирус — это неклеточный инфекционный агент. Сегодня нам известно около 6 тысяч различных вирусов, но их существует несколько миллионов. Вирусы не похожи друг на друга и могут иметь как форму сферы, спирали, так и форму сложного асимметричного сплетения. Размеры вирусов варьируются от 20 нм до 300 нм.
Как устроен вирус?
В центре агента находится генетический материал РНК или ДНК, вокруг которого располагается белковая структура — капсид.
Капсид служит для защиты вируса и помогает при захвате клетки. Некоторые вирусы дополнительно покрыты липидной оболочкой, т.е. жировой структурой, которая защищает их от изменений окружающей среды.
Вирусолог Дэвид Балтимор объединил все вирусы в 8 групп, из которых некоторые группы вирусов содержат 1-2 цепочки ДНК. Другие же содержат 1 цепочку РНК, которая может удваиваться или достраивать на своей матрице ДНК. При этом каждая группа вирусов производит себя в различных органеллах зараженной клетки.
Вирусы имеют определенный диапазон хозяев, т.е. он может быть опасен для одних видов и абсолютно безвреден для других. Например, оспой болеет только человек, а чумкой только некоторые виды плотоядных. Вирус не способен выжить сам по себе, поэтому активируется только в хозяйской клетке, используя ее ресурсы и питательные вещества. Цель вируса — создание множества копий себя, чтобы инфицировать другие клетки!
Как вирус попадает в организм?
- через физические повреждения (например, порезы на коже)
- путём направленного впрыскивания (к примеру, укус комара)
- направленного поражения отдельной поверхности (например, при вдыхании вируса через трахею)
- к эпителию слизистых оболочек (это например вирус гриппа)
- к нервной ткани (вирус простого герпеса)
- к иммунным клеткам (вирус иммунодефицита человека)

Геном вируса встраивается в одну из органелл или цитоплазму и превращает клетку в настоящий вирусный завод. Естественные процессы в клетке нарушаются, и она начинает заниматься производством и сбором белка вируса. Этот процесс называется репликацией. И его основная цель — это захват территории. Во время репликации генетический материал вируса смешивается с генами клетки хозяина — это приводит к активной мутации самого вируса, а также повышает его выживаемость. Когда процесс репликации налажен, вирусная частица отпочковывается и заражает уже новые клетки, в то время как инфицированная ранее клетка продолжает производство.
Выход вируса
Вирус создал множество собственных копий, клетка оказывается изнуренной из-за использования ее ресурсов. Больше вирусу клетка не нужна, поэтому клетка часто погибает и новорожденным вирусам приходится искать нового хозяина. Это и есть заключительная стадию жизненного цикла вируса.
Скорость распространения вирусной инфекции
Размножение вирусов протекает с исключительно высокой скоростью: при попадании в верхние дыхательные пути одной вирусной частицы уже через 8 часов количество инфекционного потомства достигает 10³, а концу первых суток − 10²³.
Вирусная латентность
Как вирус распространяется?
- воздушно-капельный (кашель, чихание)
- с кожи на кожу (при прикосновениях и рукопожатиях)
- с кожи на продукты (при прикосновениях к пище грязными руками вирусы могут попасть в пищеварительную и дыхательную системы)
- через жидкие среды организма (кровь, слюну и другие)
Почему с вирусами так тяжело бороться?
Сегодня людям уже удалось победить некоторые вирусы, а некоторые взять под жесткий контроль. Например, Оспа (она же черная оспа). Болезнь вызывается вирусом натуральной оспы, передается от человека к человеку воздушно-капельным путем. Больные покрываются сыпью, переходящей в язвы, как на коже, так и на слизистых внутренних органов. Смертность, в зависимости от штамма вируса, составляет от 10 до 40 (иногда даже 70%), На сегодняшний день вирус полностью истреблен человечеством.
Кроме того, взяты под контроль такие заболевания, как бешенство, корь и полиомиелит. Но помимо этих вирусов существует масса других, которые требуют разработок или открытия новых вакцин.
Коронавирус
Виновником эпидемии, распространяющейся сегодня по миру, стал коронавирус, вирусная частица в 0,1 микрона. Свое название он получил благодаря наростам на своей структуре, своеобразным шипам. Внутри вируса спрятан яд, с помощью которого он подчиняет себе зараженный организм. Этот вирус воздействует не только на человека, но и на птиц, свиней, собак и летучих мышей. В настоящий момент выделяют от 30 до 39 разновидностей коронавирусной инфекции. Но для человека патогенно всего 6. И как любой другой вирус COVID-19 мутирует.

К наиболее распространенным симптомам COVID-19 относятся повышение температуры тела, сухой кашель и утомляемость. К более редким симптомам относятся боли в суставах и мышцах, заложенность носа, головная боль, конъюнктивит, боль в горле, диарея, потеря вкусовых ощущений или обоняния, сыпь и изменение цвета кожи на пальцах рук и ног. Как правило, эти симптомы развиваются постепенно и носят слабо выраженный характер. У некоторых инфицированных лиц болезнь сопровождается очень легкими симптомами.
Сколько же может жить этот вирус вне организма? Все зависит от типа вируса и от той поверхности, на которую вирусы попали. В качестве примера было рассмотрено 3 вируса, по которым велись исследования. Изучали время, на которое может задерживаться вирус на различных поверхностях. Данные приведены в таблице.

Поскольку пока не изобретено вакцины от COVID-19, в целях защиты от инфекции самым важным для нас является соблюдение гигиены.
Гигиена — раздел медицины, изучающий влияние жизни и труда на здоровье человека и разрабатывающая меры (санитарные нормы и правила), направленные на предупреждение заболеваний, обеспечение оптимальных условий существования, укрепление здоровья и продление жизни.
Сегодня следует соблюдать определенные правила гигиены:
- Соблюдение режима труда и отдыха, не допускающего развития утомления и переутомления.
- Выполнение условий, обеспечивающих здоровый и полноценный сон (свежий воздух, отсутствие шума, удобная постель, оптимальная продолжительность).
- Правильное здоровое питание в соответствии с потребностями организма.
- Комфортный микроклимат в жилище (температура, влажность и подвижность воздуха, естественная и искусственная освещенность помещений).
- Содержание в чистоте тела и тщательный уход за зубами.
- Спокойное и корректное поведение в конфликтных ситуациях.

Внутри вириона гриппа А находятся восемь сегментов вирусной РНК. Эти молекулы несут всю необходимую информацию для образования новых частиц вируса гриппа. Эти восемь РНК схематично показаны желто-зелеными линиями в верхней части рис. 2. РНК представляют собой цепи из четырех различных нуклеотидов – A, C, G и U. В случае с вирусом гриппа восемь РНК имеют длину порядка 14000 нуклеотидов. Нуклеотиды составляют генетический код, который считывается преобразующим механизмом клетки в триплеты, определяющие аминокислоту.
Необходимо рассмотреть два важных аспекта этих вирусных РНК. Во-первых, вы можете видеть, что концы вирусных РНК помечены как 3′ и 5′. Нуклеиновые кислоты обладают полярностью, так что один конец цепи имеет отличные химические свойства от другого. Эта полярность представлена как 3′ и 5′. Во-вторых, во время копирования, или удвоения, нуклеиновой кислоты ферментами, которые носят название полимераз, появляется нить с комплементарной полярностью. Вирусные РНК гриппа называются (-), или РНК с отрицательной нитью, поскольку они представляют собой отрицательную полярность РНК с трансляцией в белок. Молекулы РНК, являющиеся образцами для синтеза белков, называются (+), или положительной полярностью. После вхождения в клетку отрицательная нить (-) вирусной РНК гриппа должна скопироваться в комплементарные (+) нити, так что они могут служить образцом для белков. Вирусные РНК копируются ферментом (РНК-полимеразой), который привносится в клетку вместе с вирусом.
На рис. 2 желто-зелеными линиями обозначены обнаруженные в вирионе гриппа РНК с отрицательными нитями. Как только вирион входит в клетку, эти восемь РНК копируются в положительную нить мРНК. Наконец, мРНК может служить образцом для синтеза белков. Специфические вирусные белки, производимые каждой вирусной мРНК, показаны в нижней части рис. 2. Видно, что, например, сегмент 4 РНК несет информацию о вирусном белке HA, а сегмент 6 – о вирусном белке NA. Заметьте, что некоторые сегменты РНК несут информацию более чем об одном белке. Вирусы гриппа субтипов А и B имеют восемь сегментов РНК, тогда как субтипа C – только семь.
Вирусы гриппа называются РНК-вирусами с отрицательной нитью из-за полярности РНК, привносимой в вирион. Другие РНК-вирусы, такие как полиовирус, являются РНК-вирусами c положительной нитью, так как их геномная РНК может преобразовываться в белок сразу после вхождения в клетку.
1.3. Субтипы A, B и C вируса гриппа
рисунок). Оболочечные вирионы с шестиугольной структурой на поверхности формируют длинную (500 микрон) напоминающую шнур структуру при отпочковании от клетки (рис. 4). Как и в случае с вирусами гриппа A и B, ядро вируса гриппа C состоит из рибонуклеопротеина, созданного из вирусной РНК и 4 белков. Белок M1 расположен под мембраной, как и в вирионах гриппа A и B. Второстепенный вирусный оболочечный белок CM2 функционирует как ионный канал. Основной оболочечный гликопротеин вируса гриппа C называется HEF (слияние гемагглютинина и эстеразы – hemagglutinin-esterase-fusion), поскольку он обладает функциями и HA, и NA. Поэтому вирион гриппа содержит 7 сегментов РНК, а не 8, как вирусы гриппа субтипов A и B.
Практически все взрослые когда-либо заражались вирусом гриппа C, вызывающим мягкое течение болезни в верхних дыхательных путях. Осложнения с переходом на нижние дыхательные пути – редкость. Против вируса гриппа C не существует вакцины.
Мне достаточно хорошо знакомы вирусы гриппа B и C – я получил степень доктора наук за их изучение. В моей работе говорится, что геном вируса гриппа C состоит из 7 сегментов РНК, и показана рекомбинация среди различных штаммов вируса гриппа C.
Рисунок 4.
Рекомендуемая литература.
- Hatta, M., & Kawaoka, Y. (2003). The NB Protein of Influenza B Virus Is Not Necessary for Virus Replication In Vitro Journal of Virology, 77 (10), 6050-6054 DOI:10.1128/JVI.77.10.6050-6054.2003.
- RacanielloVR, & Palese P (1979). Isolation of influenza C virus recombinants. Journal of Virology, 32 (3), 1006-14 PMID: 513198.
1.4. РНК вируса гриппа: трансляция в белок
Рисунок 6.
Рисунок 7.
Последовательности, взятые нами для образца, принадлежат штамму 1918 H1N1 вируса гриппа. Обратите внимание на аминокислоту PB1-F2, выделенную синим цветом. Эта аминокислота играет важную роль в биологическом функционировании белка, которую мы еще рассмотрим впоследствии.
В журнале Nature опубликован очень подробный обзор, характеризующих особенности строения и жизненный цикл SARS-CoV2, механизмы инфицирования и избегания своевременного иммунного ответа, обозначены потенциальные мишени для создания противовирусных препаратов. Также представлены результаты компьютерного моделирования строения вируса и механизмов проникновения в клетку.
Для того, чтобы вирус проник в клетку, помимо связи с рецептором АСЕ2, необходимо еще одно взаимодействие с трансмембранной протеазой, которая частично расщепила бы фрагмент спайк-протеина с высвобождением гидрофобных аминокислот, которые стремились бы с скорейшему слиянию с рядом расположенной мембраной клетки-хозяина. SARS-CoV2 использует с этой целью две протеазы - TMPRSS2 (преимущественно) и катепсин. Преимущественное использование TMPRSS2 и прямое проникновение вируса в клетку минуя механизм образования эндосом с участием катепсина и является объяснением неэффективности гидроксихлорохина, который воздействует исключительно на катепсин-зависимый механизм проникновения вируса.
Попав в клетку, вирус подавляет экспрессию генов клетки-хозяина, в том числе – образование интерферонов, сигнализирующих о вирусной инфекции. Для ковидной инфекции типична очень низкая концентрация интерферонов в крови.
В дальнейшем перестраивается эндоплазматический ретикулум с формированием шарообразных двухмембранных структур, в которых происходит активный синтез белков вируса. Выход вирионов наружу осуществляется в лизосомах, формирующихся в комлексе Гольджи, путем экзоцитоза. Считается, что молекулярные механизмы, ответственные за описанные процессы, могут быть мишенями для противовирусной терапии.
В месте соединения S1 и S2 субъединиц спайк-протеина есть зона, которая способна связывать и быть расщепленной фурином – протеазой клетки-хозяина, содержащейся в лизосомах. Такое предварительное расщепление значительно упрощает в дальнейшем взаимодействие с трансмембранной протеазой TMPRSS2 и значительно увеличивает проникновение вирусной частицы в следующую клетку. Для вируса SARS показано, что порядка 10% вирусных частиц на выходе из клетки – хозяина связано с фурином. У вируса SARS-CoV2 в соответствующем локусе, отвечающем за связь с фурином, идентифицированы мутации. Как следствие, до 50% вирионов альфа-разновидности SARS-CoV2, выходящих из клетки-хозяина, связаны с фурином, а в случае дельта-вируса – до 90%, что, по-видимому, и объясняет более агрессивное распространение этого варианта вируса. Есть данные, что у пациентов с дельта-формой в тканях легких и носоглотки содержание вирионов достоверно выше, чем у носителей альфа-формы.

По материалам: Megan Scudellari. How the coronavirus infects cells — and why Delta is so dangerous. Nature 595, 640-644 (2021). doi:
Читайте также: