
Белок м2 вируса гриппа
Обновлено: 22.04.2024
Штаммы вируса гриппа. Антигены вируса гриппа. Строение вируса гриппа.
Вирулентные штаммы вируса гриппа, в отличие от невирулентных, обязательно активизируются за счет расщепления протеазами. Гемагглютинины вируса гриппа расщепляются внутриклеточно и потому способны инфицировать клетки различных тканей и вызывать системную инфекцию. Ортомиксовирусы с нерасщепленным НА имеют низкую инфекционную активность. Инфекционная форма вируса с расщепленным НА в организме млекопитающих образуется только в клетках, выстилающих дыхательный тракт. При отсутствии трипсина в культуре клеток расщепляется НА только вирулентных штаммов вируса. Для вируса гриппа птиц установлена строгая корреляция между расщепляемостью НА, способностью размножаться в культуре клеток и патогенностью для кур. Расщепление НА на субъединицы не является обязательным для сборки и выхода вирионов из клетки и проявления гемагглютинирующей активности. Полагают, что фактором, детерминирующим патогенность вируса гриппа птиц, является чувствительность НА к протеолитическому расщеплению.
В сайте расщепления НА у вирулентных штаммов вируса гриппа содержится несколько основных аминокислот, а у невирулентных — только один остаток аргинина. Таким образом, только участок из нескольких основных аминокислот у С-конца НА образует сайт узнавания для ферментов, ответственных за расщепление.

Основные функции НА: гемагглютинирующая активность вируса; прикрепление вирионов к клеткам-мишеням путем связывания с сиалосодержащими рецепторами; слияние вирусной и клеточной мембран. НА является основным специфическим антигеном вируса, определяющим (наряду с NA) подтип и вызывающим образование антител, нейтрализующих инфекционность вируса и его ГА-активность. НА играет главную роль в индукции протективного иммунитета при гриппе. В молекуле НА имеются 3-4 антигенных домена, изменения в которых определяют антигенный дрейф. NA является ферментом, катализирующим отщепление сиаловой кислоты от субстрата. Удаление сиаловой кислоты с НА облегчает его расщепление клеточными протеазами. Антитела к белку NA имеют вспомогательное значение в защите и нейтрализуют вирус лишь при высоком титре. Матриксный белок М - самый низкомолекулярный структурный белок. Он принимает участие в морфогенезе вириона и стабилизации его структуры. Нуклеопротеид (NP) - основной внутренний белок, формирующий субъединицы капсида. Белки NP и М являются типоспецифическими антигенами, общими для всех вирусов гриппа одного типа, и в этом отношении резко отличаются от высокодивергентных поверхностных белков. Они не вызывают образования протективных антител. Однако нуклеопротеин вируса гриппа — основной антиген, узнаваемый цитотоксическими Т-лимфоцитами. Аминокислотные последовательности 260—283 нуклеопротеина вируса гриппа А являются индукторами Т-клеточного ответа. Изменение антигенных свойств вируса гриппа — результат двух генетических процессов: антигенных дрейфа и шифта. Антигенный дрейф происходит в основном через накопление аминокислотных замен (точечных мутаций) в НА1. Замена одного аминокислотного остатка в эпитопе нарушает его связывание с соответствующими МАТ. Основные механизмы антигенного шифта — реассортация отдельных генов, возвращение в популяцию старых генов, прямые мутации, изменяющие специфичность к хозяину. NA может измениться независимо от НА. Вариабельность НА вируса гриппа А значительно выше вариабельности НА вируса гриппа В.
В процессе адаптации вируса гриппа к различным системам наблюдают изменение антигенной структуры НА, в основе которой лежит селекция мутантов с измененной рецепторсвязывающей специфичностью. Вирусы гриппа А и В человека, размноженные в КЭ, могут изменяться антигенно и претерпевать мутации в ГА (замена трех аминокислот в ГА). Вирус, размноженный в КЭ или в культуре клеток, защищает против культурального вируса более эффективно, чем против полевого вируса, выделенного от человека.
Один и тот же клинический изолят вируса гриппа при размножении в культуре клеток животных (МДСК) или в КЭ имеет разные антигенные свойства, что связано с заменой одной-двух аминокислот в разных участках молекулы НА. Особенно заметные изменения антигенности в молекуле НА наблюдали при замене аминокислотных остатков 187 и 189 в процессе адаптации вируса к КЭ. Изменение сайта гликозилирования может сопровождаться изменением антигенности и вирулентности вируса. Состав углеводов гемагглютинина вирусов и гриппа птиц может изменяться в различных хозяйских клетках даже в течение одного пассажа. У вирусов гриппа птиц типа А идентифицировано девять нейраминидазных N-антигенов, обозначенных 1-9, и 13 гемагглютинирующих антигенов Н, обозначенных 1-13. Состав Н- и N-антигенов полевых изолятов зависит от вида птиц, места и времени вспышек гриппа. Свиньи восприимчивы к различным подтипам вируса гриппа А и, возможно, вместе с водоплавающей птицей являются главным резервуаром вируса гриппа в природе.
В пермиссивных клетках вирусный НА активируется путем расщепления на две части НА1 и НА2, которые остаются связанными дисульфидными связями. Вирионы прикрепляются к клеткам, активированным НА, соединяются с рецепторами сиаловой кислоты плазматической мембраны и входят в клетку эндоцитозом. После сплавления оболочки вируса и эндоплазматической мембраны транскрипционный комплекс освобождается и транспортируется в ядро, где происходит транскрипция и репликация РНК.
Так же, как у всех других вирусов с негативно-полярным РНК-геномом, геном ортомиксовирусов выполняет две функции: 1) матричную для синтеза мРНК и 2) матричную для синтеза позитивно-полярной промежуточной формы РНК, которая служит матрицей для синтеза потомства геномной РНК. Из 8 первичных транскриптов, синтезированных на 8 генных сегментах вирусов гриппа А и В, 6 являются моноцисторными и транслируются прямо в белки. Два других — подвергаются сплайсингу с образованием двух мРНК, которые транслируются с различных рамок считывания с образованием двух белков. Вирусные белки синтезируются, используя клеточный механизм трансляции. Ортомиксовирусы используют несколько механизмов для увеличения кодирующей способности: сплайсинг мРНК, спаренную стоп-старт-трансляцию тандемных генов и сдвиг рамки считывания.
Репликация геномных РНК сегментов требует синтеза полноразмерных, положительной полярности РНК посредников, которые, в отличие от соответствующих мРНК транскриптов, не имеют КЭП-структуры на 5'-конце и поли (А) последовательности на З'-конце. Вновь синтезированный нуклеопротеин присоединяется к этим РНК, облегчая их использование в качестве матрицы для синтеза геномной РНК. Вирионы формируются почкованием, включая М белок и нуклеокапсид, которые встроены на плазматической мембране, в которую включены НА и NA. Механизмы копирования каждого РНК сегмента и включение их в каждый вирион неизвестны.
Хотя в результате реассортации генов теоретически могут возникать вирусы с любой комбинацией Н и N генов, однако только ограниченное число вирусов с определенной комбинацией Н и N генов оказались важными патогенами, имеющими право на существование. В том числе вызывающие респираторную патологию у людей: H1N1, H2N2, H3N2, H5N1 и, возможно, H3N8; у лошадей H7N7 и H3N8; у норок H10N4; у котиков H7N7 и H4N5; вирусы, часто выделяемые от свиней - H1N1 и H3N2. Главными возбудителями гриппа птиц являются вирусы с H5N2 и H7N1, хотя встречаются и другие варианты вируса гриппа А. Так как установлена резистентность диких видов птиц к вирусу гриппа А, им отводили роль резервуара вируса, прежде всего для домашней птицы. При экспериментальном заражении вирусом H1N5 дикие утки, как правило, не проявляли клинических признаков болезни, хотя вирус в их организме размножался и выделялся во внешнюю среду. Латентное инфицирование вирусом гриппа может иметь место у других домашних и диких животных.
Грипп - острое инфекционное заболевание, вызванное вирусом гриппа (семейство Orthomyxoviridae), которое в холодное время года поражает до 15% населения земного шара. Актуальность данного заболевания отражена в статистических данных: ежегодно в мире до 500 млн человек заболевают гриппом, из них 2 млн случаев заканчиваются смертельным исходом. Если рассматривать ситуацию в рамках российской действительности, то ежегодно регистрируют от 27,3 до 41,2 млн человек, заболевших гриппом и ОРВИ. Каждый 6-7 россиянин вовлекается в эпидемический процесс.
ГРИПП - МЕДИКО-СОЦИАЛЬНАЯ И ЭПИДЕМИОЛОГИЧЕСКАЯ ПРОБЛЕМА

При попадании в верхние дыхательные пути одной вирусной частицы уже через 8 ч количество инфекционного потомства достигает 10 3 , а к концу первых суток - 10 23 . Высокая скорость размножения объясняет короткий инкубационный период -от одних до двух суток. Вирус, обладая выраженным цитолитическим эффектом, повреждает клетки трахеи и бронхов с последующей дистрофией, некрозом и десквамацией эпителия.
Вирус попадает в кровь, разносится по всему организму, активирует систему протеолиза, повреждает эндотелий капилляров, угнетает кроветворение.
ВИРУС ГРИППА ПЫТАЕТСЯ ВНЕДРИТЬСЯ В ЭПИТЕЛИЙ ВЕРХНЕГО ОТДЕЛА РЕСПИРАТОРНОГО ТРАКТА
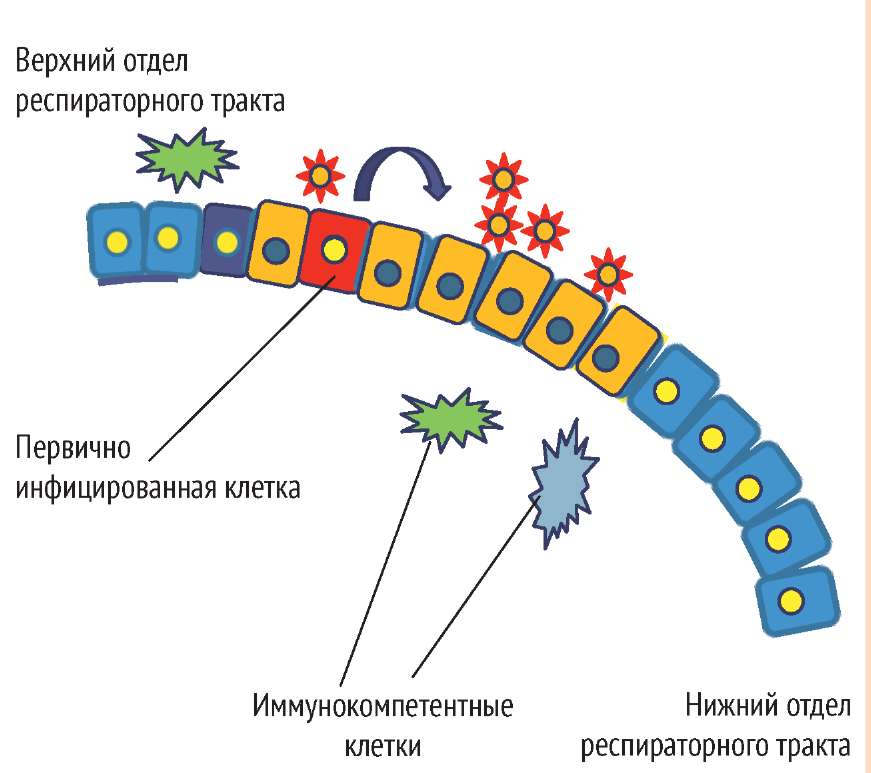
Инфекционная доза вируса гриппа 10-100 вирусных частиц
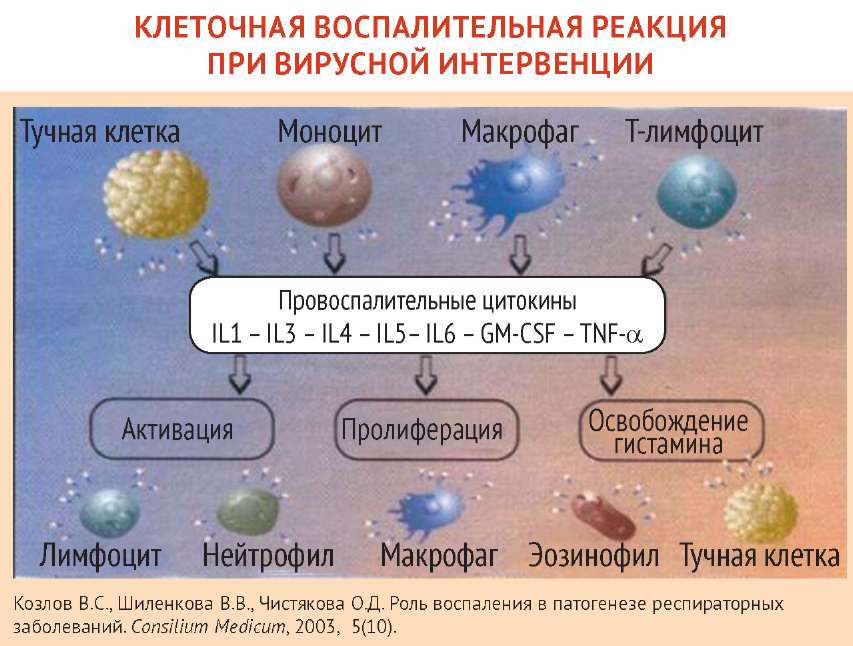
Патогенез гриппозной инфекции
Возбудителем гриппозной инфекции является вирус гриппа, который относится к оболочечным РНК-содержа-щим вирусам с негативным геномом. Внутри вириона находятся 8 сегментов РНК, связанных с белками поли-меразного комплекса и белком нукле-окапсида. Внутреннюю сторону липидной мембраны вируса выстилает основной структурный мембранный белок М1, в нее же погружен формирующий ионные каналы М2-белок. Снаружи в липидную мембрану вируса погружены два поверхностных гликопротеида вируса гриппа - гемагглютинин (НА) и вирусный фермент нейраминидаза (NA), которые обуславливают необычайную исключительную изменчивость вируса гриппа [2]. Патогенность вирусов гриппа зависит от всех генов и кодирующихся ими вирусоспецифических белков. Рецептор-связывающий белок -гемагглютинин (HA) взаимодействует со специфическим рецептором клетки - сиаловой кислотой, поэтому НА называют рецептор-связывающий белок. Кроме того, он стимулирует экспорт вирусного генома к месту сборки вируса. НА высокопатогенных вирусов расщепляется протеазами респираторного тракта и фуриноподобными протеазами, которые синтезируются в различных органах и системах, что придает патогенным вирусам способность поражать различные органы и ткани. НА определяет способность вируса к преодолению межвидовых барьеров. Изменения структуры гемагглютинина составляют основу ускользания вирусов гриппа от специфического иммунного ответа.
В ПАТОГЕННОСТИ ВИРУСОВ ГРИППА ИГРАЮТ РОЛЬ ВСЕ ГЕНЫ И КОДИРУЮЩИЕСЯ ИМИ ВИРУСОСПЕЦИФИЧЕСКИЕ БЕЛКИ
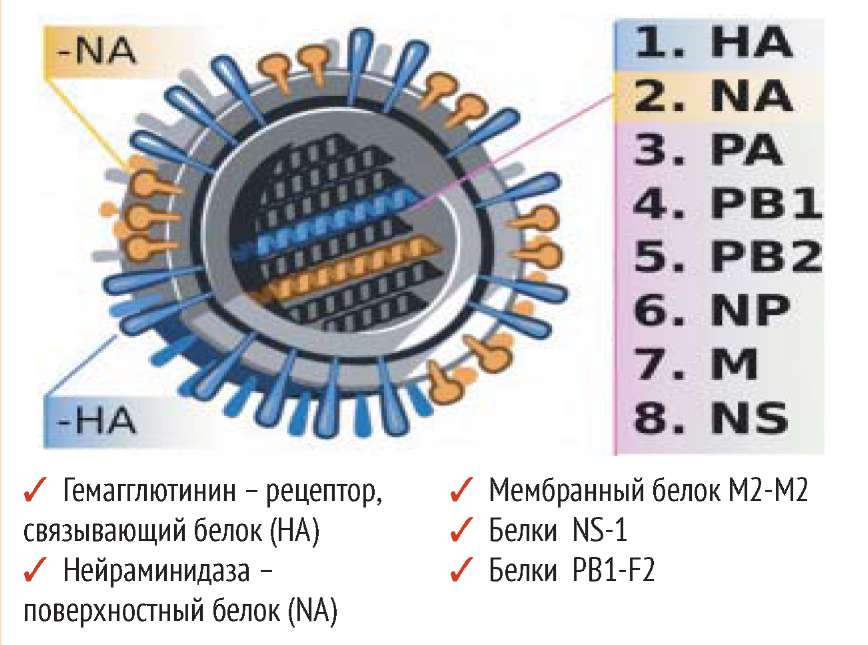
Нейраминидаза (NA) удаляет сиаловую кислоту от НА, позволяя ему освободиться от поверхности клетки и участвовать в репродукции вируса. Нейраминидаза участвует в процессе расщепления вновь синтезированной молекулы НА. НА и NA - протеолиз, активация плазминогена и мембранных протеаз, индукция цитокинового шторма.
Острое начало заболевания характеризуется резким повышением температуры тела до 38 0 C и выше, выраженной интоксикацией (озноб, головная боль, ломота в мышцах, суставах, боль при движении глазными яблоками или надавливании на них, светобоязнь) с последующим присоединением катарально-респираторных симптомов (сухость слизистых верхних дыхательных путей, першение в горле, першение или боли за грудиной, сухой кашель, одышка) [1]. Главная особенность течения заболевания, вызванного вирусом гриппа, в том, что симптомы интоксикации опережают развитие катарального синдрома, который выражен поражением респираторного тракта: трахеитом (кашель, саднение и боли за грудиной), часто с при соединением геморрагического синдрома (носовые кровотечения, мокрота с прожилками крови).
КЛИНИЧЕСКИЕ СИНДРОМЫ И СИМПТОМЫ ГРИППА
? Острое начало с синдрома интоксикации
? Симптомы интоксикации опережают развитие катарального синдрома
? Наличие лихорадки (фебрильная, постоянного типа)
? Ведущий синдром поражения респираторного тракта - трахеит (кашель, саднение и боли за грудиной)
? Наличие геморрагического синдрома (носовые кровотечения, мокрота с прожилками крови)
Тактика ведения больного гриппом
В лечении больных гриппом необходимо придерживаться комплексного лечения этиопатогенетической направленности. Комплексность и поэтапность терапии включают назначение противовирусных лекарственных препаратов (желательно в первые 48 ч от появления симптомов заболевания) при регистрации врачом гриппоподобного заболевания либо лабораторно подтвержденного диагноза грипп. В острый период заболевания больному показана противовоспалительная и жаропонижающая терапия: ибупрофен, целекоксиб, напроксен, парацетамол, анальгин, литическая смесь, а также дезинтоксикационная и антиоксидантная терапия с включением ингибиторов протеолиза, назначением флавоноидов и витаминов с адекватной респираторной поддержкой и использованием отхаркивающих, сосудосуживающих и антиконгестивных препаратов [1].

ПРОТИВОВИРУСНАЯ ТЕРАПИЯ: ПРЕПАРАТЫ С ПРЯМЫМ ПРОТИВОВИРУСНЫМ ЭФФЕКТОМ (РЕКОМЕНДАЦИИ ВОЗ)
Дни заболевания
Патогенетические механизмы
Терапия
Противовирусная терапия: осельтамивир, занамивир, умифеновир
Антигистаминные препараты: хлоропирамин, дезлоратадин, дифенгидрамин
Жаропонижающая терапия: парацетамол, анальгин, литическая смесь
системной
воспалительной
реакции с активацией
специфического
протеолиза
Противовоспалительная терапия Блокаторы ЦОГ: целекоксиб, напроксен, диклофенак
Антиоксиданты: поливитамины, витамин С, витамин Е, дигидрокверцетин, рутозид, ацетилцистеин
Ингибиторы протеолиза: апротинин (Контрикал, Аэрус), аминометилбензойная кислота, аминокапроновая кислота
Сосудосуживающие препараты: ксилометазолин, оксиметазолин, фенилэфрин, нафазолин Отхаркивающие средства: ацетилцистеин, гвайфенезин, амброксол
Период ранней реконвалесценции с возможным развитием осложнений
Контроль за состоянием сердечнососудистой, дыхательной, нервно-эндокринной системы пациентов Продолжение симптоматической терапии и назначение растительных и синтетических иммуномоделирующих препаратов
Основные классы противовирусных препаратов
Согласно анатомо-терапевтически-химической (АТХ) классификации все лекарства делятся на группы согласно их основному терапевтическому применению. Появление противовирусных препаратов прямого действия (ПППД, direct-acting antiviral, DAA) является по-настоящему революционным. Опыт лечения с применением ПППД говорит о том, что использование этих препаратов значительно повышает частоту выздоровления больных гриппом. По определению экспертов Food and Drug Administration (США), противовирусными могут быть названы только препараты, оказывающие непосредственное прямое воздействие на репликацию вируса, т. е. действие этих препаратов направлено на определенную вирусоспецифическую мишень в цикле размножения вируса. Существует несколько классов препаратов прямого противовирусного действия. Мишенью действия противогриппозных препаратов адамантанового ряда (амантадин и римантадин) является М2-белок вируса гриппа. Функцию вирусного фермента нейраминидазы блокируют противогриппозные препараты осельтамивир и занамивир. Вирусоспецифической мишенью действия умифеновира (Арбидол) в цикле вирусной репродукции является поверхностный белок вируса гриппа гемагглютинин (НА).
ПРОТИВОВИРУСНАЯ ТЕРАПИЯ: ПРЕПАРАТЫ С ПРЯМЫМ ПРОТИВОВИРУСНЫМ ЭФФЕКТОМ (РЕКОМЕНДАЦИИ ВОЗ)
Класс
Мишень
Препараты
Ингибитор слияния (фузии)
Осельтамивир (Тамифлю™) Занамивир (Реленза™)
Ионные каналы М2
Амантадин Ремантадин Орвирем
* Не рекомендован ВОЗ в связи с отсутствием к нему чувствительности большинства сезонных штаммов гриппа (Guidelines for Pharmacological Management of Pandemic Influenza A(H1N1) 2009 and other Influenza Viruses. Revised February 2010. Part I: Recommendations).
Механизм действия противовирусного препарата умифеновир (Арбидол)
Арбидол (умифеновир) - препарат с прямым противовирусным эффектом, код АТХ: [J05AX], включен в АТХ-классификацию как препарат с прямым противовирусным действием.
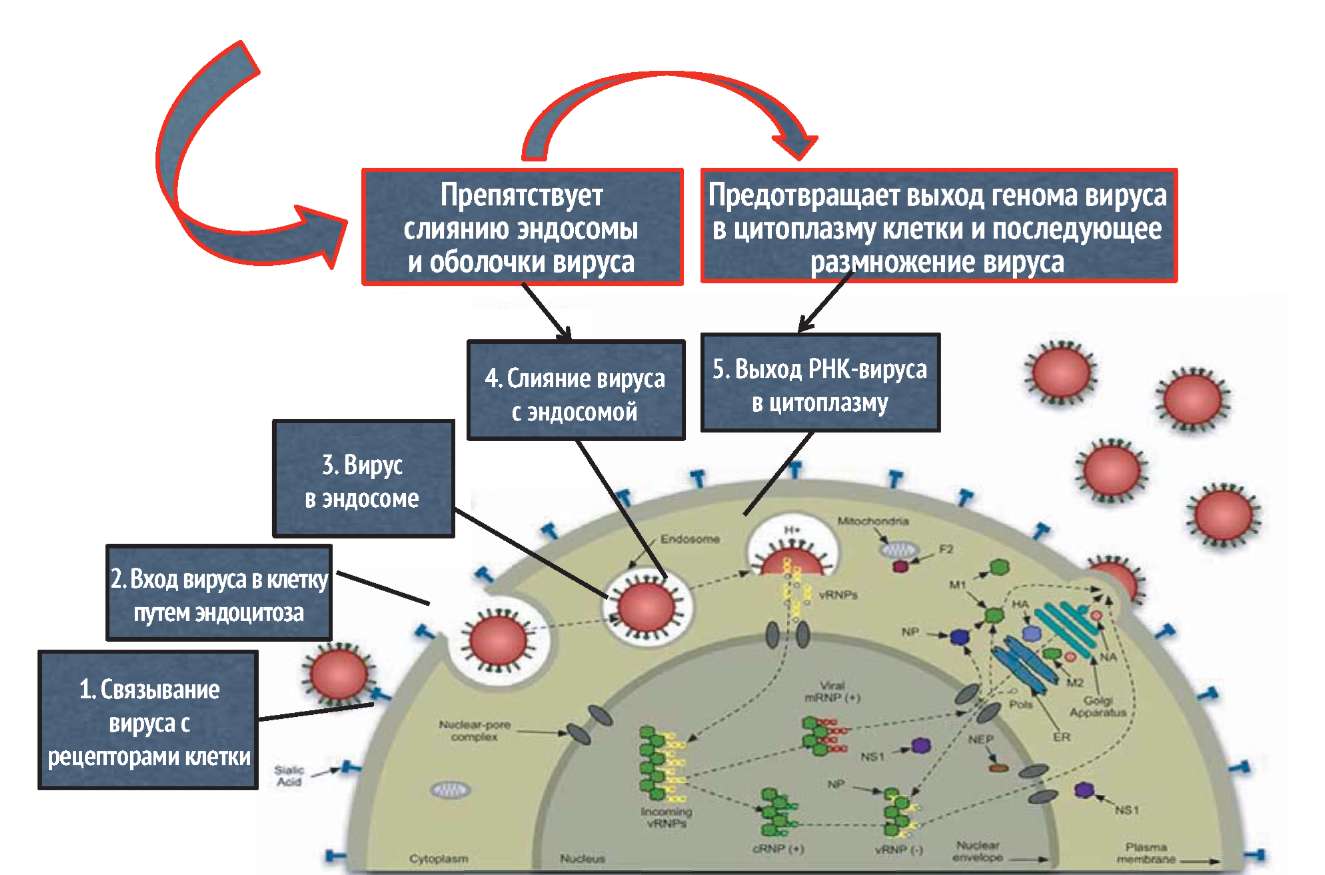
Важнейшим показателем прямого противовирусного действия препарата является его способность или способность его метаболитов подавлять репродукцию вируса в клетках, которая численно выражается в значении ингибиторной концентрации 50 (ИК50 - концентрация препарата, ингибирующая размножение вируса на 50%). Анализ данных, полученных в различных отечественных и зарубежных исследовательских центрах, позволяет сделать однозначный вывод о том, что Арбидол отвечает всем стандартным международным требованиям, предъявляемым к противовирусным препаратам. Арбидол (умифеновир) взаимодействует с гемагглютинином (НА), препятствует его конформации, необходимой для слияния НА с мембранами эндосом. Предотвращает выход генома вируса в цитоплазму клетки и последующее размножение вируса гриппа.
АРБИДОЛ (УМИФЕНОВИР) ВЗАИМОДЕЙСТВУЕТ С ГЕМАГГЛЮТИНИНОМ, ПРЕПЯТСТВУЕТ ЕГО КОНФОРМАЦИИ (РАСПОЛОЖЕНИЕ АТОМОВ В МОЛЕКУЛЕ), НЕОБХОДИМОЙ ДЛЯ СЛИЯНИЯ С МЕМБРАНАМИ ЭНДОСОМ
МЕХАНИЗМ ДЕЙСТВИЯ АРБИДОЛА
Boriskin Y.S., Leneva I.A., Pecheur E.I., PoLyak SJ. ArbidoL: a broad-spectrum antiviral compound that blocks viral fusion. Curr Med Chem., 2008, 15(10): 997-1005.
Схема назначения Арбидола
Арбидол (умифеновир) может назначаться взрослым и детям с 3-летнего возраста. Обязательным условием является курсовое назначение препарата и раннее его применение -при первых симптомах заболевания (в первые 48 ч).
СХЕМА НАЗНАЧЕНИЙ ПРЕПАРАТА АРБИДОЛ ВЗРОСЛЫМ И ДЕТЯМ СТАРШЕ 12 ЛЕТ
Лечение гриппа и ОРВИ
200 мг 4 раза в день, курс 5 дней
Постконтактная профилактика гриппа и ОРВИ
200 мг 1 раз в сутки, курс 10-14 дней
Сезонная профилактика гриппа и ОРВИ
200 мг 2 раза в неделю, курс 3 недели
Результаты исследования терапевтической эффективности Арбидола
ИССЛЕДОВАНИЯ ПРЕПАРАТА АРБИДОЛ ПРОДОЛЖАЮТСЯ
Двойное слепое рандомизированное 1 плацебо-контролируемое многоцентровое клиническое 1 исследование по изучению эффективности 1 и безопасности препарата Арбидол® 1 при лечении и профилактике гриппа и других ОРВИ 1
850 пациентов ПЦР-диагностика 16 вирусных возбудителей ОРВИ, в т. ч. 4 видов вируса гриппа
Цель исследования
? Получение дополнительных данных о безопасности и терапевтической эффективности препарата Арбидол® у пациентов с диагнозом грипп или ОРВИ
? Получение дополнительных данных о безопасности и эффективности применения препарата Арбидол® для профилактики гриппа/ОРВИ среди контактных лиц заболевшего
? Оценка частоты возникновения резистентных к препарату Арбидол® штаммов вируса гриппа при терапевтическом и профилактическом применении препарата.
- снижает тяжесть течения инфекционного заболевания и длительность острого периода;
- абсолютное большинство пациентов к 4-му дню наблюдения демонстрировали элиминацию возбудителя из организма человека, что подтверждает высокую противовирусную активность Арбидола.
ПРИ ПРИЕМЕ ПРЕПАРАТА АРБИДОЛ У ПАЦИЕНТОВ С ГРИППОМ НАБЛЮДАЕТСЯ БОЛЕЕ ЛЕГКОЕ ТЕЧЕНИЕ ЗАБОЛЕВАНИЯ И СЛАБАЯ ВЫРАЖЕННОСТЬ ОСНОВНЫХ СИМПТОМОВ ИНТОКСИКАЦИИ ВЛИЯНИЕ АРБИДОЛА НА СРОКИ ЭЛИМИНАЦИИ ВИРУСА ГРИППА
Известный факт
1. Выраженность симптомов гриппа коррелирует со степенью вирусной нагрузки, поэтому применение противовирусного препарата снижает тяжесть течения инфекционного заболевания и длительность острого периода
2. Противовирусный препарат сокращает период элиминации вируса гриппа
1. Carrat F., Vergu E., Ferguson N., Lemaitre M., Cauchemez S., Leach S. & Valleron A. Time Lines of Infection and Disease in Human Influenza. A Review of Volunteer Challenge Studies American Journal of Epidemiology, 2008, 167(7): 775-785.
2. Sophia Ng et al. Effects of oseltamivir treatment on duration of clinical illness and viral shedding, and household transmission of influenza virus. Clin Infect Dis., 2010, 50(5): 707-714.
Ранее проведенные исследования доказали, что Арбидол сокращает длительность основных симптомов ОРВИ негриппозной этиологии на 1,5-2 сут., а общую продолжительность болезни - на 2, 4 сут.
Активность Арбидола в отношении других возбудителей ОРВИ
Арбидол активен в отношении широкого спектра вирусов, вызывающих респираторные инфекции у человека: респираторно-синцитиального вируса, аденовируса, вируса парагриппа, риновируса, коронавируса.
АРБИДОЛ СОКРАЩАЕТ ДЛИТЕЛЬНОСТЬ ОСНОВНЫХ СИМПТОМОВ ОРВИ НЕГРИППОЗНОЙ ЭТИОЛОГИИ НА 1,5-2 СУТ., ОБЩУЮ ПРОДОЛЖИТЕЛЬНОСТЬ БОЛЕЗНИ - НА 2, 4 СУТ.
Малеев В.В., Селькова Е.П., Простяков И.В., Осипова ЕА. Фармакоэпидемиологическое исследование течения гриппа и других ОРВИ в сезоне 2010-2011 гг. Инфекционные болезни, 2012, 10(3): 15-23.
АРБИДОЛ АКТИВЕН В ОТНОШЕНИИ ВИРУСОВ ГРИППА А И В, А ТАКЖЕ РЯДА ДРУГИХ ВОЗБУДИТЕЛЕЙ ОРВИ
Семейство
Возбудитель
Показатель активности Арбидола
Вирус гриппа А H1N1 A/PR/8/34 (сезонный) A/California/07/2009 (пандемич.)
Внутри вириона гриппа А находятся восемь сегментов вирусной РНК. Эти молекулы несут всю необходимую информацию для образования новых частиц вируса гриппа. Эти восемь РНК схематично показаны желто-зелеными линиями в верхней части рис. 2. РНК представляют собой цепи из четырех различных нуклеотидов – A, C, G и U. В случае с вирусом гриппа восемь РНК имеют длину порядка 14000 нуклеотидов. Нуклеотиды составляют генетический код, который считывается преобразующим механизмом клетки в триплеты, определяющие аминокислоту.
Необходимо рассмотреть два важных аспекта этих вирусных РНК. Во-первых, вы можете видеть, что концы вирусных РНК помечены как 3′ и 5′. Нуклеиновые кислоты обладают полярностью, так что один конец цепи имеет отличные химические свойства от другого. Эта полярность представлена как 3′ и 5′. Во-вторых, во время копирования, или удвоения, нуклеиновой кислоты ферментами, которые носят название полимераз, появляется нить с комплементарной полярностью. Вирусные РНК гриппа называются (-), или РНК с отрицательной нитью, поскольку они представляют собой отрицательную полярность РНК с трансляцией в белок. Молекулы РНК, являющиеся образцами для синтеза белков, называются (+), или положительной полярностью. После вхождения в клетку отрицательная нить (-) вирусной РНК гриппа должна скопироваться в комплементарные (+) нити, так что они могут служить образцом для белков. Вирусные РНК копируются ферментом (РНК-полимеразой), который привносится в клетку вместе с вирусом.
На рис. 2 желто-зелеными линиями обозначены обнаруженные в вирионе гриппа РНК с отрицательными нитями. Как только вирион входит в клетку, эти восемь РНК копируются в положительную нить мРНК. Наконец, мРНК может служить образцом для синтеза белков. Специфические вирусные белки, производимые каждой вирусной мРНК, показаны в нижней части рис. 2. Видно, что, например, сегмент 4 РНК несет информацию о вирусном белке HA, а сегмент 6 – о вирусном белке NA. Заметьте, что некоторые сегменты РНК несут информацию более чем об одном белке. Вирусы гриппа субтипов А и B имеют восемь сегментов РНК, тогда как субтипа C – только семь.
Вирусы гриппа называются РНК-вирусами с отрицательной нитью из-за полярности РНК, привносимой в вирион. Другие РНК-вирусы, такие как полиовирус, являются РНК-вирусами c положительной нитью, так как их геномная РНК может преобразовываться в белок сразу после вхождения в клетку.
1.3. Субтипы A, B и C вируса гриппа
рисунок). Оболочечные вирионы с шестиугольной структурой на поверхности формируют длинную (500 микрон) напоминающую шнур структуру при отпочковании от клетки (рис. 4). Как и в случае с вирусами гриппа A и B, ядро вируса гриппа C состоит из рибонуклеопротеина, созданного из вирусной РНК и 4 белков. Белок M1 расположен под мембраной, как и в вирионах гриппа A и B. Второстепенный вирусный оболочечный белок CM2 функционирует как ионный канал. Основной оболочечный гликопротеин вируса гриппа C называется HEF (слияние гемагглютинина и эстеразы – hemagglutinin-esterase-fusion), поскольку он обладает функциями и HA, и NA. Поэтому вирион гриппа содержит 7 сегментов РНК, а не 8, как вирусы гриппа субтипов A и B.
Практически все взрослые когда-либо заражались вирусом гриппа C, вызывающим мягкое течение болезни в верхних дыхательных путях. Осложнения с переходом на нижние дыхательные пути – редкость. Против вируса гриппа C не существует вакцины.
Мне достаточно хорошо знакомы вирусы гриппа B и C – я получил степень доктора наук за их изучение. В моей работе говорится, что геном вируса гриппа C состоит из 7 сегментов РНК, и показана рекомбинация среди различных штаммов вируса гриппа C.
Рисунок 4.
Рекомендуемая литература.
- Hatta, M., & Kawaoka, Y. (2003). The NB Protein of Influenza B Virus Is Not Necessary for Virus Replication In Vitro Journal of Virology, 77 (10), 6050-6054 DOI:10.1128/JVI.77.10.6050-6054.2003.
- RacanielloVR, & Palese P (1979). Isolation of influenza C virus recombinants. Journal of Virology, 32 (3), 1006-14 PMID: 513198.
1.4. РНК вируса гриппа: трансляция в белок
Рисунок 6.
Рисунок 7.
Последовательности, взятые нами для образца, принадлежат штамму 1918 H1N1 вируса гриппа. Обратите внимание на аминокислоту PB1-F2, выделенную синим цветом. Эта аминокислота играет важную роль в биологическом функционировании белка, которую мы еще рассмотрим впоследствии.
В. В. Зарубаев* кандидат биологических наук
А. А. Штро*, кандидат биологических наук
Е. Н. Свентицкий**, доктор технических наук, профессор
* ФГБУ НИИ гриппа МЗ РФ, Санкт-Петербург ** ГосНИИ ОЧБ ФМБА России, Санкт-Петербург
Резюме. Рассмотрены подходы к профилактике респираторных вирусных инфекций, включая применение средств инактивации внеклеточного вируса на поверхностях и в объеме помещений, что позволяет предотвратить инфицирование людей в местах их скоплений.
Ключевые слова: респираторные вирусные инфекции, грипп, инфекционная активность, санитарные меры, профилактика.
Abstract. Approaches to prevention of respiratory viral infections were considered, including application of inactivation drugs for extracellular virus on the surfaces and inside the facilities, which allows to prevent contamination in gathering of people. Keywords: respiratory viral infections, influenza, infectious activity, sanitary measures, prophylaxis.
Наиболее эффективным средством профилактики гриппа является вакцинация. Однако вследствие постоянной смены антигенных свойств возбудителя требуется постоянный мониторинг и разработка новых вакцинных штаммов, соответствующих циркулирующим в человеческой популяции в каждый конкретный эпидемический сезон.
В отношении группы производных адамантана можно отметить сравнительно высокую токсичность и узкий спектр действия (препараты активны против гриппа А, но не против гриппа В, не имеющего белка M2). Для ингибиторов нейраминидазы характерна высокая стоимость синтеза, что делает эти препараты менее доступными для широкого использования. Кроме того, к препаратам обеих групп вирус гриппа способен вырабатывать устойчивость [10, 11].
Однако для более надежного контроля заболеваемости наряду с вакцинопрофилактикой и химиотерапией важную роль играют средства инактивации внеклеточного вируса на поверхностях и в объеме помещений. Такие меры позволяют во многом предотвратить инфицирование людей в местах их скоплений, таких как общественные места, массовые мероприятия и т. п. Для обеззараживания поверхностей и объемов помещений используются ультрафиолетовые лампы различного спектрального диапазона и химические средства дезинфекции.
Преимуществом ультрафиолетового облучения является его комплексный механизм. Эффективность такого воздействия на патогены обусловлена как прямым воздействием излучения, так и реакционно-способными молекулами озона О3, генерируемыми при взаимодействии ультрафиолетовых квантов с кислородом воздуха. Следует, однако, отметить, что доза облучения падает пропорционально квадрату расстояния от источника и для инактивации вируса на поверхностях в помещениях потребуется время, также пропорциональное квадрату линейных размеров помещения. Кроме того, для эффективного обеззараживания поверхностей требуется прямая их экспозиция к облучению, что не всегда достижимо в помещениях сложного профиля и с установленным оборудованием и мебелью. В этом случае основной эффект дезинфекции достигается за счет диффузии молекул озона, имеющей меньшую эффективность.
В России в настоящее время для обеззараживания помещений из дезинфицирующих средств в форме аэрозоля применяют [12]:
- 37%-е растворы формальдегида;
- 20%-й раствор параформальдегида с добавлением 1% едкой щелочи;
- 24%-й раствор глутарового альдегида;
- 30%-й раствор алкамона;
- препарат надуксусной кислоты;
- 10%-й раствор перекиси водорода с добавлением 1% муравьиной кислоты.
Для дезинфекции воздуха и поверхностей помещений в присутствии животных в форме аэрозоля применяют молочную кислоту, йод, триэ-тиленгликоль и гипохлорит натрия. Следует указать на такие недостатки этих препаратов, как:
- высокая токсичность для человека;
- необходимость направленного распыления при обработке поверхностей;
- необходимость дополнительной очистки или нейтрализации после обработки деконтаминантом (например, 25% аммиак после обработки формальдегидом и глутаральдегидом, раствор железосинероди-стого калия после обработки алкамоном и пр.);
- высокие нормы расхода препарата дезинфектанта (15-90 мл/м 3 или 100-200 мл/м 2 при обработке поверхностей).
1. Химические окислители. Сюда входят такие соединения, как кислоты (фосфорная, соляная, лимонная и др.), соли хлорсодержа-щих кислот (хлориты, гипохлориты), перекись водорода, йод и пр. Такие вещества оказывают неспецифическое разрушающее действие на все материалы, и степень этого разрушения зависит от устойчивости материала к окисляющим агентам, при этом биологические соединения, составляющие бактерии и вирусы, оказываются более уязвимыми по сравнению с материалами самих помещений, мебели и оборудования, чем и обусловлен эффект дезинфекции. Окислители могут применяться как для протирания поверхностей, так и для обеззараживания воздуха посредством аэрозолирования. В любом случае, однако, лишь некоторые из них (перекись водорода) впоследствии распадаются при взаимодействии с компонентами воздуха. Многие другие дезинфектанты этой группы (кислоты, йод и пр.), напротив, сохраняют свою химическую активность и реакционную способность, чем создают потенциальную опасность для человека и животных, находящихся в дальнейшем в обработанных помещениях. В особенности актуален этот аспект при аэрозольной обработке и с учетом летучести активных соединений. Например, пары йода представляют опасность для слизистых оболочек ротовой полости, глаз и носоглотки и при попадании на них могут привести к побочным последствиям. Кроме того, соединения, действующие при помощи активного хлора (например, препараты Хлорокс (компания Clorox), Жавель солид (компания Жазол) и др.), имеют ограниченный срок действия вследствие малой стабильности хлора. Поэтому при проведении дезинфекционных мероприятий следует либо готовить рабочие растворы непосредственно перед дезинфекцией, либо контролировать концентрацию активного хлора, что дополнительно усложняет процедуру деконтаминации [15].
2. Поверхностно-активные вещества. Как правило, аммониевые производные жирных кислот (С12-С18). Их активность обусловлена воздействием на липидные оболочки и белковые капсиды вирусов, и их использование ограничивается в основном дезинфекцией поверхностей. Эти соединения не распадаются со временем, поэтому после обработки требуется дополнительная процедура промывки дезинфицированных поверхностей водой или другим дезинфектантом.
3. Фиксирующие и денатурирующие агенты. Сюда относятся этанол, фенол, формальдегид, глутаральдегид и пр. Активность соединений этого класса является следствием их способности к дегидратации (этанол), денатурации (фенол) или образованию перекрестных сшивок (формальдегид) в биологических молекулах. К характерным особенностям этих соединений относится их высокая летучесть, с одной стороны, обеспечивающая их высокую проникающую способность, а с другой — представляющая потенциальную опасность при контакте с органами дыхания.
К примеру, одним из основных средств деконтаминации против гриппа птиц за рубежом является препарат Виркон (Virkon®S) производства DupontAnimalHealthiSolutjons или KRKA, представляющий собой смесь соединений перекиси, поверхностно-активных веществ, органических кислот и неорганических буферных систем [13]. Главным компонентом состава является калия персульфат 50%. Препарат обладает антимикробной активностью в отношении грамотрицательных и грамположительных бактерий (кроме микобактерий туберкулеза), вирусов (включая ВИЧ, гепатит В, герпес, грипп, полиомиелит, ротавирус, энтеровирусы и др.), грибов рода Candida, дермато-фитов. В отношении спор бактерий препарат не проявляет активности.
С целью оценки дезинфицирующих свойств персонального обез-зараживателя воздуха нового типа (генератора диоксида хлора) как средства инактивации возбудителей воздушно-капельных вирусных инфекций было проведено исследование его вирулицидного действия в аэрозольной камере с распыленным вируссодержащим аэрозолем, содержащим вирус гриппа А. После активации средства (извлечения кассеты из герметичной алюминиевой упаковки) запускается медленное испарение активного компонента (диоксида хлора), который уничтожает болезнетворные вирусы и бактерии в окружающем воздухе.
Материалы и методы исследования
Подготовка обеззараживателя воздуха для тестирования
Обеззараживатель воздуха индивидуальный (генератор диоксида хлора производства Global Product Planning Co., Ltd.) извлекали из защитного пакета, выдерживали в течение 1 часа в открытом состоянии и помещали в аэрозольную камеру в воздухонепроницаемый пакет. Момент открывания пакета считали нулевой временной точкой эксперимента.
Исследование защитных свойств обеззараживателя воздуха индивидуального (генератора диоксида хлора) 3,0 мл суспензии вируса (5 х 10 7 ЕЮ50/мл) распыляли в течение 10 минут в камере объемом 0,4 м 3 с помощью вихревого пневматического генератора аэрозоля типа ВАГ-2, расположенного в центре камеры и обеспечивающего получение аэрозоля с расчетной концентрацией 3,8 х 10 4 ЕЮ50/л с массовым медианным диаметром частиц 3,6 мкм. Для равномерного распределения аэрозоля в объеме камеры использовали постоянно работающий вентилятор. Система для отбора проб воздуха включала насос, измеритель скорости потока и микроциклон для улавливания частиц аэрозоля, содержащий 10 мл физиологического раствора.
Отбор проб воздуха в микроциклоны для определения концентрации вируса, находящегося в воздушной фазе объема камеры, проводили в течение 2 минут при скорости потока 7,2 л/мин, что приблизительно соответствует режиму дыхания человека. Забор материала проводили в периоды 1-3, 3-5 и 10-12 мин после начала экспозиции обеззаражи-вателя воздуха индивидуального (генератора диоксида хлора).
В контрольной серии экспериментов те же манипуляции проводили в отсутствие изучаемого обеззараживателя воздуха индивидуального (генератора диоксида хлора).
Вирус
В работе использовали вирус гриппа A/California/07/09 (H1N1)pdm09. Вирус культивировали в течение 48 часов при 36 °С в аллантоисной полости 10-12-дневных куриных эмбрионов. В качестве исходного инфекционного материала использовалась аллантоисная жидкость.
Подготовка инфекционного материала
Вируссодержащую аллантоисную жидкость центрифугировали в течение 30 минут при 4 °С и 4000 об./мин. После осаждения фрагментов мембран и крупных контаминирующих частиц надосадок центрифугировали при 36 000 g и 4 °С в течение 1 часа. Осажденные вирионы ресуспендировали в физиологическом растворе, равном по объему исходному количеству аллантоисной жидкости. Полученная вирусная суспензия имела физические характеристики, близкие к таковым для воды. Это позволило использовать ее для создания аэрозоля, что было затруднительно при использовании аллантоисной жидкости с высоким содержанием белков и, как следствие, большой вязкостью. Для замедления высыхания частиц аэрозоля, приводящего к быстрой потере инфекционной активности, к вирусной суспензии добавляли глицерин до конечной концентрации 5%.
Титрование инфекционной активности вируса
Аликвоты физиологического раствора отбирали из микроциклонов и готовили из них серию десятикратных разведений на фосфатном буфере. 10-12-дневные куриные эмбрионы заражали серийными десятикратными разведениями вирусного материала от 100 до 10-6 по 0,2 мл на эмбрион и инкубировали в термостате при 36 °C в течение 48 часов. По окончании срока инкубации эмбрионы охлаждали, вскрывали и переносили аллантоисную жидкость (0,1 мл) в лунки планшета для иммунологических реакций, после чего добавляли равный объем 1% куриных эритроцитов в физиологическом растворе.
Уровень репродукции вируса в эмбрионах оценивали по реакции гемагглютинации эритроцитов. За титр вируса принимали величину, обратную наибольшему разведению вируса, способного вызвать положительную реакцию гемагглютинации, и выражали в логарифмах 50% инфекционной дозы вируса (lgID50). На основании полученных результатов на каждом сроке эксперимента оценивали эффективность инактивации — снижение титра вируса в% от контроля.
Статистическая обработка результатов
Статистическую обработку результатов оценки активности проводили с помощью непараметрического U-критерия Манна-Уитни. Различия считали значимыми при р < 0,05.
Результаты
Данные по титрованию проб воздуха на инфекционную активность вируса гриппа суммированы в табл. (для наглядности представлены на рис.).
Таблица.
Инактивирующая активность обеззараживателя воздуха индивидуального (генератора диоксида хлора) в отношении вируссодержащего аэрозоля
Мы кратко рассмотрели структуру вирионов гриппа и процесс кодирования вирусными РНК одного и более белков. Теперь обратимся к размножению вирусов гриппа.
Вирусы – это исключительно внутриклеточные паразиты, они не могут воспроизводиться вне клетки. Появление новых инфекционных частиц должно протекать внутри клетки. Войдя в клетки, вирусы начинают паразитировать в структуре хозяина для воспроизводства вирусного потомства. Все события, происходящие в инфицированной вирусом клетке, называются инфекционным циклом, или вирусной репликацией. Вирусологи искусственно разделяют инфекционный цикл на несколько стадий, чтобы проще было его изучить. Эти стадии включают прикрепление и вхождение вириона, трансляцию мРНК в белок, репликацию генома (создание большего количества РНК или ДНК), скопление новых частиц и выход частиц из клетки. Мы рассмотрим каждую из названных стадий, а затем обсудим, как вирус гриппа инфицирует нас и вызывает болезнь.
Остановимся на первой стадии – прикреплении вириона к клетке. Возьмем обычную клетку, с которой, я уверен, знаком каждый. Но не будет лишним повториться.
Рисунок 10.
Слева показан клеточный белок, прикрепленный к цитоплазматической мембране. Снизу изображена внутренняя часть клетки – цитоплазма. Часть белка пересекает мембрану, и также видны части цитоплазматической и внеклеточной сторон. Сферы – это сахара, прикрепленные ко множеству белков (белок + сахар = гликопротеин). Сиаловая кислота всегда является последним сахаром в цепи, которая прикрепляется к белку. Справа показана химическая структура сиаловой кислоты; следующий сахар (справа) – галактоза. Вирионы гриппа прикрепляются к клеткам, когда HA захватывает очень маленькую сиаловую кислоту.
Сахар в сравнении с HA в действительности очень мал – он подходит к небольшому кармашку на острие шипа. На рис. 11 изображена молекулярная модель, демонстрирующая, как HA прикрепляется к аналогу сиаловой кислоты. Шаровидный конец HA показан в верхней части рис. 11. Маленькие красные и белые сферы показывают места будущего прикрепления сиаловой кислоты – кармашки на верхнем конце HA.
Рисунок 13.
Процесс вхождения вируса гриппа в клетки – наиболее понятный из всех известных механизмов проникновения вирусов. После прикрепления вириона к сиаловой кислоте, имеющей рецепторы на поверхности клетки, вирус-рецепторный комплекс входит в клетки посредством эндоцитоза – процесса, в ходе которого клетки обычно берут молекулы из внеклеточной жидкости. Пока эндосомальные везикулы, содержащие частицы вируса, движутся к ядру клетки, их pH понижается. Это изменение в pH завершает клеточный канал, закачивающий протоны (H+) в везикулу. Когда эндосомальная pH достигает 5,0, вирусный белок HA подвергается конформационной перестройке. Это изменение подвергает пептид слияния на HA короткой гидрофобной последовательности, вставляющей его в эндосомальную мембрану, которая от этого сливается с вирусной оболочкой. Когда это происходит, вирусные РНК проникают в цитоплазму. Затем они транспортируются в ядро клетки, где начинается размножение.
В вирионе гриппа вирусные РНК не оголены, поскольку к ним прикреплены вирусные белки, включая белок M1. Этот белок образует оболочку, лежащую в основе липидной мембраны вириона. К сожалению, если вирусные РНК прикрепляются к белку M1, когда выходят из вириона, они не могут достичь ядра. Для решения этой проблемы вирион гриппа имеет в своей мембране несколько копий белка, называемого M2. Этот вирусный белок образует канал в мембране, который активно закачивает протоны из эндосомы во внутреннюю часть вириона. Данные протоны понижают pH внутри вириона, освобождая вирусные РНК от M1. В этом случае РНК могут войти в ядро.
Ионный канал M2, являющийся мишенью для противовирусных адамантанов, показан на рис. 14. Эти образования забивают канал и не позволяют протонам закачиваться в вирион. В присутствии адамантанов вирусным РНК нужно прикрепляться к M1, отчего они не могут достичь ядра. Поэтому вирусная репликация подавляется. Устойчивость к адамантанам возможна при изменении в аминокислотах, прокладывающих канал M2. Такие изменения не дают лекарству забить канал.
Рисунок 15.
На рис. 15 сферический конец белка HA, который прикрепляется к рецепторам клетки, изображен сверху, а вирусная мембрана – снизу. Для наглядности помечен только сайт расщепления HA. Нерасщепленная форма белка называется HA0; после расщепления клеточным ферментом образуются два белка, называемые HA1 (синий) и HA2 (красный). Две субъединицы остаются вместе на поверхности вирусной частицы. Новая амино(N)-концевая область HA2, образовавшегося при расщеплении, содержит последовательность гидрофобных аминокислот, называемых пептидом слияния. Во время вхождения вируса гриппа в клетки пептид слияния вставляется в эндосомальную мембрану и вызывает слияние вирусной и клеточной оболочек. Следовательно, вирусные РНК гриппа могут войти в цитоплазму. Процесс слияния описан в предыдущем параграфе.
Если белок HA не расщеплен, чтобы образовать HA1 и HA2, слияния не происходит. Поэтому вирусы гриппа с нерасщепленным HA не заразны. Расщепление вирусного HA происходит после того, как вновь синтезированные вирионы выходят из клетки. Вирусы гриппа эффективно размножаются в яйцах из-за присутствия протеазы в аллантоисной жидкости, способной расщеплять HA. Однако репликация многих штаммов вируса гриппа в культурах клеток требует добавления в среду соответствующей протеазы (как привило, трипсина).
У людей репликация вируса гриппа ограничена дыхательными путями, потому что это единственный участок, где производится протеаза, расщепляющая HA. Однако белок HA высокопатогенных штаммов H5 и H7 вируса птичьего гриппа может расщепляться протеазами, производимыми во многих различных тканях. В результате эти вирусы способны размножаться во многих органах птиц, включая селезенку, печень, легкие, почки и мозг. Это свойство объясняет способность штаммов H5N1 вируса птичьего гриппа размножаться вне дыхательных путей человека.
Подобно белкам HA высокопатогенных вирусов H5 и H7, HA штамма вируса гриппа 1918 г. может также расщепляться повсеместно присутствующими клеточными протеазами. Следовательно, вирус может размножаться в культурах клеток без добавления трипсина.
Белки HA H5 и H7 имеют множество основных аминокислотных остатков на сайте расщепления HA1-HA2, которые делают возможным расщепление посредством широко представленных протеаз. Но HA вируса гриппа H1 1918 г. не обладал этим свойством. Также N1 1918 г. не был в состоянии привлечь протеазы, которые расщепляли бы HA, т. е. не было механизма, позволяющего штамму A/WSN/33 вируса гриппа размножаться в клетках без трипсина. Понимание того, как белок HA вируса H1 1918 г. мог быть расщеплен протеазами – основной в понимании высокой патогенности данного штамма.
Рекомендуемая литература.
Chaipan, C., Kobasa, D., Bertram, S., Glowacka, I., Steffen, I., Solomon Tsegaye, T., Takeda, M., Bugge, T., Kim, S., Park, Y., Marzi, A., &Pohlmann, S. (2009). Proteolytic Activation of the 1918 Influenza Virus Hemagglutinin Journal of Virology, 83 (7), 3200-3211 DOI:10.1128/JVI.02205-08.
Читайте также: