
Белые мыши восприимчивы к вирусу
Обновлено: 23.04.2024
Энцефаломиелит мыши
Энцефаломиелит мышей — инфекция лабораторных мышей, проявляющаяся вялыми параличами преимущественно задних конечностей. Заболевание вызывают несколько вирусов: оригинальный шт. ТО-Тейлора, а также шт. FA, GD-VII и родственные им штаммы. Встречается повсеместно.
Штамм ТО впервые выделил в 1933 г. Тейлор из головного мозга белых мышей у которых наблюдал клинические признаки энцефалита. Аналогичный вирус Тейлора можно легко выделить из организма клинически здоровых белых мышей, что свидетельствует о широком вирусоносительстве.
Клинические признаки и патологоанатомические изменения. Вирус ТО обычно вызывает латентную инфекцию у лабораторных мышей. Аналогичные вирусы выделяли от диких мышей, хлопковых крыс и мешотчатых грызунов. Патологические изменения в ЦНС от шт. ТО сходны с поражениями, вызываемыми вирусом полиомиелита у человека. Отмечается деструкция клеток передних рогов, особенно спинного мозга, и связанная с ней воспалительная реакция. Изменения, обусловленные шт. FA и GD-VII, характеризуются диффузным энцефалитом и не совсем сходны с изменениями при полиомиелите у человека, как в случаях с вирусом ТО.
Штаммы вируса энцефаломиелита мышей Тейлора распределены в 2 группы: GDVII и DA (или ТО). Нейровирулентная группа, которая быстро убивает мышей, и демиелинизирующая группа, которая вызывает нелетальную инфекцию двигательных нервов с последующей персистентной инфекцией белого вещества с демиелини-зирующими повреждениями, сходными с множественным склерозом. На основании исследования 3-мерной структуры вируса предполагают, что различия в патогенезе двух групп вируса Тейлора являются результатом различий в процессах иммунологического или рецептор опосредованного узнавания.
Референтные шт. GDVII и DA. Штаммы подгруппы GDVII являются высоковирулентными и вызывают острый энцефаломиелит у мышей. При этом не наблюдается ни персистенции, ни демиелинизации. Штаммы DA менее вирулентны и устанавливают персистентную инфекцию в спинном мозге мышей несмотря на энергичную гуморальную иммунную реакцию, сопровождающуюся хронической демиелинизацией. Вирус представляет собой превосходную модель для изучения множественного склероза человека, при котором также наблюдается демиелинизация. Большинство клеток, инфицированных вирусом, производит 100—500 копий генома, что намного меньше, чем необходимо для определяемой экспрессии вирусных белков. Ограниченный синтез вирусных белков ведет к уменьшению цитопатического эффекта. Анализ последовательностей мутантных вирусов, устойчивых к нейтрализующим МКА к БД идентифицировал 4 аминокислотных мутации капсидных белков и все они локализовались на рецепторе или вблизи связывающего вирус. Мутантные вирусы показали уменьшение демиелинизирующей активности. Эти находки предполагают, что нейтрализация антител зависит от связывания вируса с клеточным рецептором и что демиелинизация и вирусная персистенция могут интерферировать с предотвращением прикрепления вируса к рецепторам особых типов клеток. Белок 17 кД (L-белок) синтезируется в штаммах субгруппы DA. Изучение вирусного мутанта DA без L-npo-теинового синтеза показало, что этот белок является важным для вирусного роста в особых типах клеток и является критическим для демиелинизирующей болезни и вирусной персистенции. Инактивация гена рецептора у-интерферона делает мышей I29/SV чувствительными к персистентнОой инфекции и ДБ, а инактивация гена у-интерферона у мышей 157ВL/6 придает им чувствительность к персистентной инфекции, а не к клинической ДБ. Эти различия фенотипа обусловлены различиями генетического фона. Полное секвенирование генома в потомстве F2 из двух скрещиваний показало, что ДБ является результатом двух независимых факторов: высокой вирусной нагрузки и локуса Tmevd5 в хромосоме 11.
Морфология, химический состав и устойчивость. РНК-содержащий вирус Тейлора имеет размеры 20—25 нм, устойчив в диапазоне рН 3—10, а также к эфиру, хлороформу, фенолу, гипертоническому раствору NaCl, глицерину, алюминиевым квасцам. Разрушается под воздействием 20 %-ного раствора этилового спирта, 50 %-ного ацетона, 2 %-ного раствора КМп04, 1,5 %-ного раствора формалина. Длительно сохраняется при низких температурах (-40 ° — 70 °С), неустойчив к прогреванию (50 °-70 °С) и высокочувствителен к лиофильному высушиванию. АГ структура не изучена. У больных мышей старше года обнаруживают ВНА, ПА, анти-ГА.
Штаммы ТО, FA и GD-VII родственны, но не идентичны. ВЭМТ индуцирует ген c-fos в чистых культурах покоящихся астроцитов мозга мыши. Экспрессия c-fos зависит от дозы ВЭМТ и достигает максимума при множественности заражения 100. Экспрессия c-fos максимальная через 30 мин и исчезает через 2 ч. Методом проточной цитометрии показано, что ВЭМТ быстро интернализируется после связывания со специфическим рецептором и активно реплицируется в цитоплазме астроцитов и что мРНК c-fos может транслироваться в ядре. Индукция c-fos нейтрализуется антителами к ВЭМТ. Очищенные белки VP1, VP2 и VP3 ВЭМТ не индуцируют экспрессию ВЭМТ. Быстрая индукция c-fos может являться первой стадией заражения астроцитов ВЭМТ.
Изучение структуры ВЭМТ позволило предположить, что глубокая впадина на поверхности вирионов участвует в прикреплении вируса к рецептору. Для проверки этого предположения у менее вирулентного варианта ВеАп ВЭМТ сконструировали замены четырех аминокислотных остатков (V1091, Р1153, А1225 и Р1379). В каждом из этих сайтов сделаны три аминокислотные замены: одна консервативная и две более радикальные. Получены семь жизнеспособных мутантов вируса и изучены их связывание с клетками ВНК-21, стабильность капсидов при 40 °, репликация РНК, кинетика роста в одиночном и множественных циклах заражения и трансляция вирусных белков. Три из указанных остатков участвуют в прикреплении вируса к клеточным рецепторам.
Шт. GD-VII в отличие от других (FA и ТО) агглютинирует эритроциты человека группы 0 при температуре 4 °С до разведения выше 1:2000. После обработки трипсином ГА-активность штамма повышается и проходит при температуре 20 °С с эритроцитами человека и 4 °С с эритроцитами обезьяны. После обработки трипсином два других штамма (FA и ТО) также приобретают ГА-свойства при температуре 20 °С.
Латентно инфицированные мыши выделяют вирус с фекалиями в течение 53 дней.
Экспериментальная инфекция. Экспериментальная инфекция шт. ТО воспроизводится на мышах 3—4-недельного возраста при интрацеребральном, интраназальном, интраперитонеальном и внутримышечном введениях (гибель при первом методе введения в 100 % случаев). Взрослые мыши не заболевают или легко переболевают и становятся вирусоносителями. Вирус проникает В ЦНС и другие органы через 2—5 дней после заражения. Он локализуется, в основном, в стенке кишечника и оттуда проникает в мезентериальные лимфоузлы и далее в ЦНС. У зараженных мышей старшего возраста может развиваться прогрессирующий вялый паралич, у еще более старших — латентная инфекция. После первичного выделения шт. GD-VII обычно вызывает вялый паралич, после пассирования на мышах — генерализованный энцефалит. Шт. FA также постоянно вызывает только диффузный энцефалит. Вялые параличи отмечают только после интраперитонеальной инокуляции.
Внутримышечная инокуляция любого из трех штаммов вызывает локальный миозит, по крайней мере шт. GD-VII, видимо, размножается в мышцах. Получена серия мутантов штамма DA со специфическими мутациями вирусного капсидного белка VP1, определяющего связь вируса с рецепторами клетки. Мутант вируса с заменой в петле IJ/PJ треонина на аспарагин (T81D) формирует крупные бляшки, но обладает пониженной способностью к репликации in vitro. При инъекции этого мутанта восприимчивым мышам выявляется измененный тропизм в острой стадии заболевания и не развивается хроническое демиелинизирующее заболевание. При замене треонина на валин (T81V) вирус не вызывает изменения, характерные для заболевания мышей. При замене в этой позиции триптофана (T81W) развивается сходное острое заболевание, но не развивается хроническая болезнь. Изменение аминокислот в гидрофобной области вдавления стенки, в позиции 91 VP1, в гидрофильный треонин (V91T), приводит к острому или хроническому заболеванию в отсутствие персистен-ции вируса. Таким образом, замены аминокислот в области петли I VP1 вируса приводят к изменению взаимодействия вируса с хозяином.
Культивирование. Вирус FA размножается в КЭ, зараженных на ХАО, в желточный мешок или в аллантоисную полость. Кроме эмбрионов, вирус размножается в культуре фибробластов КЭ, в культуре клеток печени и почек мышей, а также в ВНК-21. Особенности внутриклеточной репродукции не изучены. Шт. DA вызывает в клетках L929 персистенцию, которая обусловлена продукцией интерферона. Синтез РНК В клетках ВНК-21, зараженных шт. GDVI1 Или DA снижается через 3 ч после заражения, достигает максимального уровня через 6 ч, затем снижается. Изучена чувствительность первичных культур нейронов штаммами DAh GDVTI ВЭМТ. Высоковирулентный штамм GDVII заражает более 90 % Нейронов через 12 ч после инокуляции, а через 24 час. после заражения вызывает сильный цитопатический эффект. Антигены ВЭМТ обнаруживаются только через 24 ч после заражения вызывающим хрони-ческеую демиелинизирующую болезнь штаммом DA, который инфицирует не более 2 % нейронов даже через 6 дней. Заражение штаммом DA не вызывает цитопатичес-кого эффекта. С использованием рекомбинантных ВЭМТ показано, что капсид действительно отвечает за картину заражения, наблюдаемую in vitro. Таким образом, уровень нейровирулентности штаммов DA и GDVII коррелируете их способностью заражать культуры нейронов и контролируется капсидом. Штамм GD VII не индуцирует наработку полипептида вследствии его плохого размножения в мышиных L-клетках.
Источники и пути передачи инфекции. Источником инфекции при спонтанном энцефалите служат больные мыши, выделяющие вирус во внешнюю среду с калом и мочой, а также мыши-вирусоносители. Они являются основным резервуаром вирусов в природе. Заражаются мыши в основном алиментарным путем, через инфицированный экскрементами корм. Передавать вирус здоровым животным могут кровососущие насекомые.
Кроме белых мышей, восприимчивы серые дикие мыши, хлопковые крысы. У морских свинок удавалось вызвать латентную инфекцию. Кролики, белые и водяные крысы, лесные мыши и обезьяны к вирусу резистентны. На ранней стадии инфииирования мышей штаммом Даниеля вируса энцефаломиелита мышей вирус обнаруживается в нейронах и глии. В белом веществе вирус персистирует дольше, тогда как под воздействием иммунной системы происходит его ранняя элиминация из серого вещества. Из рыжей полевки выделен неизвестный ранее пикорнавирус, названный вирусом Люген. Принадлежность к пикорнавирусам была установлена на основании данных ЭМ и секвенирования. Рассчитанные аминокислотные последовательности капсидных белков VP2 и VP3 вируса Люген были сходны с таковыми вируса ЕСНО-22 (сходство около 70 %). 5'-некодирующая область РНК вируса Люген характеризовалась наибольшим сходством с кардиовирусами.
Иммунитет и специфическая профилактика. Иммунитет не изучен, специфическая профилактика не разработана. Новорожденные мышата и мышата-сосунки приобретают пассивный иммунитет с материнскими AT. Вскоре после отъема они становятся вирусоносителями и обладают повышенной резистентностью к инфекции. С возрастом процент вирусоносителей уменьшается: в 7,5 месяца он составлял всего 50 % и в дальнейшем понижался. Инфекция мышей вирусом энцефаломиелита Тейлора напоминает по клиническим проявлениям рассеянный склероз (PC) человека. Сконструированы экспрессирующие эукариотические векторы, кодирующие капсидные белки VP1, VP2 и VP3. Вакцинировали мышей SJL/J внутримышечно однократно, дважды или трехкратно кДНК различных капсидных белков. Затем мышей заражали в мозг. Трехкратная вакцинация мышей кДНК, кодирующей VP2 вируса, приводит к частичной защите их ЦНС от демиелинизирующего заболевания: уменьшались клинические симптомы и гистопатология. Вакцинация с помощью кДНК, кодирующей VP3, также приводила к уменьшению клинических проявлений. В то же время, вакцинация кДНК, кодирующей VP1, вызывала более тяжелое заболевание, причем симптомы заболевания появлялись раньше, а гистопатология усиливалась в сравнении с контролем. Это не коррелировало с титрами антител против и течением болезни. Таким образом, вакцинация с помощью ДНК способна изменять течение хронического демиелинизирующего заболевания. Это может найти применение в лечении PC.

Прежде чем любая вакцина для Ковид-19 сможет увидеть свет, она должна пройти тщательное тестирование. Первым делом на животных. В основном исследования проводятся на мышах, крысах, хомяках, обезьянах и даже хорьках. Этот список может расшириться в ближайшее время.
Под моделями животных в научной номенклатуре подразумеваются животные, которые благодаря своим особым характеристикам могут использоваться для исследований заболеваний, для испытаний пробных лекарств и экспериментальных вакцин. Прежде чем какое-либо лекарственное средство вступает в фазу клинических испытаний на человеке, его сначала тестируют на животных. Только положительный результат таких доклинических исследований разрешает дальнейшие попытки, уже с участием людей.
Почему лабораторные исследования проводятся только на определенных видах животных?
Для экспериментальных лекарств или вакцин, которые будут испытаны на животных, эти животные должны отличаться определенными признаками. Проще говоря, их организмы должны реагировать совершенно аналогично человеческим. В мире существует несколько десятков официальных биолабораторий, занимающихся разведением животных в исследовательских целях. Теперь каждая из этих биолораторий хочет предоставлять ученым животных с такими функциями, которые окажутся полезными в борьбе с Ковид-19 и помогут понять, как коронавирус атакует организм и как его победить. В настоящее время приоритетом является тестирование имитационных вакцин путем иммунизации животных и затем намеренного заражения коронавирусом.
Более раннее тестирование данного препарата на животных важно не только с точки зрения его эффективности, но и безопасности. Например, на этом этапе исследования можно увидеть серьезные побочные эффекты от пробного препарата или вакцины. Во время работы над вакциной против SARS было отмечено, например, что препарат, который давали животным, даже усугублял болезнь, вызывая неподходящие антитела.
Эксперименты на животных помогают объяснить, как дети с меньшей вероятностью получают Ковид-19, а также как распространяется инфекция и какие генетические факторы делают некоторых людей более восприимчивыми к тяжелым заболеваниям. Даже приобретение устойчивости к коронавирусу было впервые продемонстрировано в исследованиях с участием обезьян.
Сирийские хомяки страдают от Ковид-19, как люди

Ученые возлагают большие надежды на сирийских хомяков. При попытке понять, как коронавирус действует на организм и как его преодолеть, используются те животные, которые восприимчивы к вирусу и имеют симптомы инфекции, которые как можно ближе напоминают симптомы у людей. Хомяк уже использовался для исследования коронавируса во время эпидемии атипичной пневмонии несколько лет назад. Затем исследователи обнаружили, что они могут легко заразиться коронавирусом, но симптомы болезни умеренные. Однако теперь выяснилось, что SARS-CoV-2 вызывает гораздо более сильные симптомы в их организме. После заражения животных оказалось, что вирус накапливается в их легких и кишечнике, то есть там, где у них больше всего рецепторов ACE2 (рецепторов, к которым присоединяется коронавирус).
Лабораторные мыши в исследованиях на COVID-19 - только генетически модифицированные
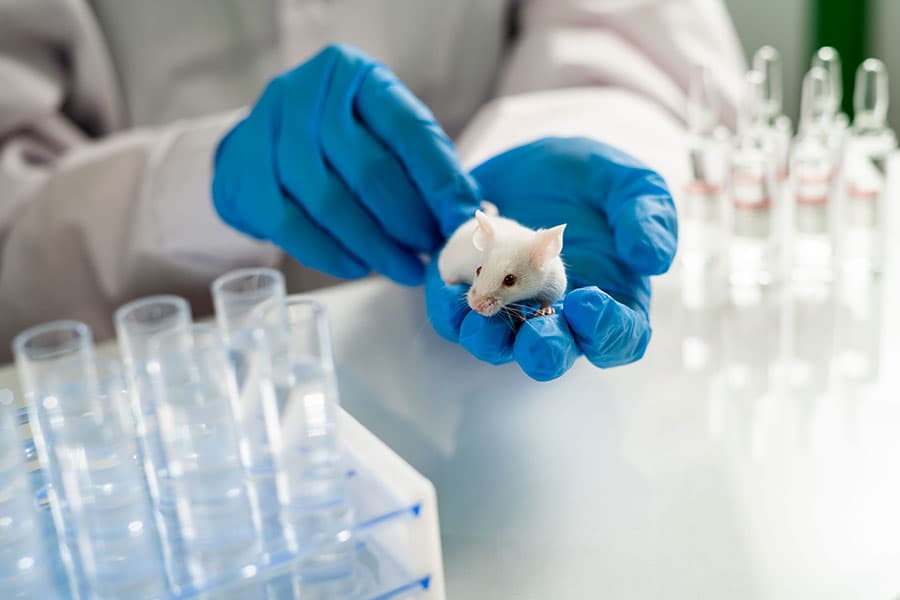
Нормальные мыши бесполезны для коронавируса, потому что их рецептор ACE2 не похож на человека и поэтому не имеет точки прикрепления. Но мыши чрезвычайно популярны в лабораториях и их легко разводить, поэтому ученые нашли выход. Таким образом, выращивают мышей, которые посредством генной инженерии приобретают ген рецептора ACE2 человека.
Крысы для целей Ковид-19 также должны быть трансгенными

В некоторых лабораториях для тестирования используют крыс, потому что они крупнее мышей, и тогда некоторые тесты легче выполнять. Крыс для исследования препарата на Ковид-19 также необходимо специально разводить, чтобы получить человеческий рецептор ACE2. В вакцинных и токсикологических исследованиях в основном используют этих животных.
Хорьки - животные, которые более восприимчивые к вирусу гриппа
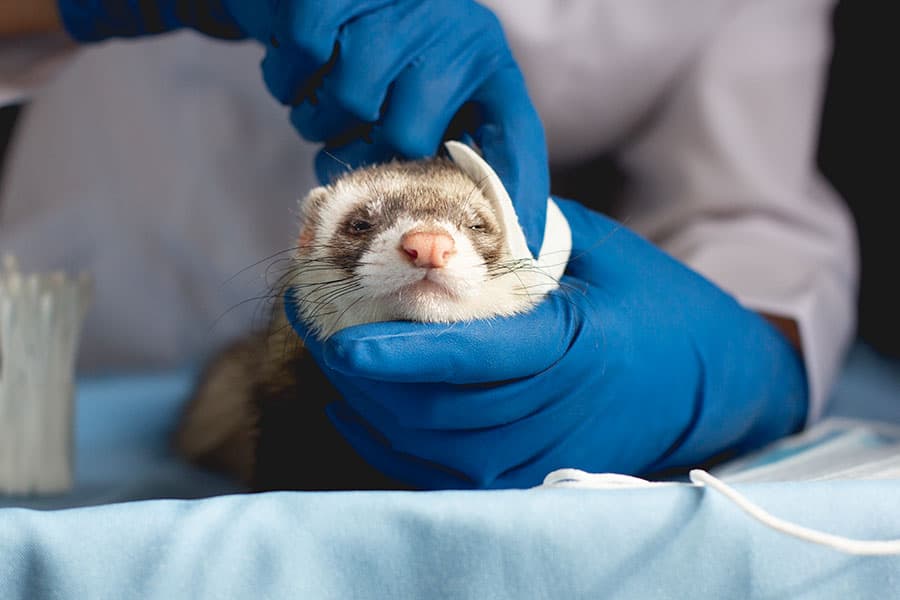
Хорьков часто используют для изучения гриппа, их легко заразить этим вирусом, их реакции очень похожи на человеческие. Видимо хорьки даже чихают как люди. Они плохо реагирует на новый коронавирус, и нет никаких серьезных симптомов заболевания, кроме более высокой температуры. Но это исследование хорьков помогло сделать вывод, что заражение может происходить не только через капельный маршрут, но и через так называемый аэрозоль.
Некоторые исследователи также считают, что тестирование хорька может помочь нам понять, почему Ковид-19 намного опаснее для пожилых людей.
Обезьяны из-за их генетического сходства с людьми наиболее ценны при тестировании лекарств и вакцин

На сегодняшний день попытки заразить обезьян новым коронавирусом относятся к четырем видам, среди которых африканская зеленая обезьяна (Chlorocebus sabaeus), резус, циномолгус и мартышка. Именно благодаря исследованиям на обезьянах было показано, что включение Ковид-19 обеспечивает, по меньшей мере, кратковременный иммунитет, что является ключевой новостью для разработчиков вакцин.>
Обезьян также используют для изучения распространения SARS-CoV-2 в форме аэрозоля (туман). Также считается, что обезьяны могут быть хорошим источником знаний о взаимосвязи между основными заболеваниями, такими как гипертония и диабет, и тяжелой формой Ковид-19.
Информация о производителе
Компания БиоПитомник СТЕЗАР предлагает купить лабораторных животных. Мы производим на собственных площадях, доставляем по всей России и предлагаем приобрести:
- Мыши нелинейные ICR (CD-1), SHK
- Мыши Линейные: CBA (Lac), c57Bl/6, Balb/c, гибриды F1

Все наши лабораторные животные выращены в питомнике, расположенном в экологически чистой природной зоне. Вся продукция компании сертифицирована в соответствии с Положением о контроле качества питомников и экспериментально-биологических клиник (вивариев), а также в соответствии с Национальными, Региональными, Международными, Отраслевыми стандартами и техническими регламентами.
Для доставки используется собственный изотермический транспорт с подогревом
Энцефаломиелит мыши
Энцефаломиелит мышей — инфекция лабораторных мышей, проявляющаяся вялыми параличами преимущественно задних конечностей. Заболевание вызывают несколько вирусов: оригинальный шт. ТО-Тейлора, а также шт. FA, GD-VII и родственные им штаммы. Встречается повсеместно.
Штамм ТО впервые выделил в 1933 г. Тейлор из головного мозга белых мышей у которых наблюдал клинические признаки энцефалита. Аналогичный вирус Тейлора можно легко выделить из организма клинически здоровых белых мышей, что свидетельствует о широком вирусоносительстве.
Клинические признаки и патологоанатомические изменения. Вирус ТО обычно вызывает латентную инфекцию у лабораторных мышей. Аналогичные вирусы выделяли от диких мышей, хлопковых крыс и мешотчатых грызунов. Патологические изменения в ЦНС от шт. ТО сходны с поражениями, вызываемыми вирусом полиомиелита у человека. Отмечается деструкция клеток передних рогов, особенно спинного мозга, и связанная с ней воспалительная реакция. Изменения, обусловленные шт. FA и GD-VII, характеризуются диффузным энцефалитом и не совсем сходны с изменениями при полиомиелите у человека, как в случаях с вирусом ТО.
Штаммы вируса энцефаломиелита мышей Тейлора распределены в 2 группы: GDVII и DA (или ТО). Нейровирулентная группа, которая быстро убивает мышей, и демиелинизирующая группа, которая вызывает нелетальную инфекцию двигательных нервов с последующей персистентной инфекцией белого вещества с демиелини-зирующими повреждениями, сходными с множественным склерозом. На основании исследования 3-мерной структуры вируса предполагают, что различия в патогенезе двух групп вируса Тейлора являются результатом различий в процессах иммунологического или рецептор опосредованного узнавания.
Референтные шт. GDVII и DA. Штаммы подгруппы GDVII являются высоковирулентными и вызывают острый энцефаломиелит у мышей. При этом не наблюдается ни персистенции, ни демиелинизации. Штаммы DA менее вирулентны и устанавливают персистентную инфекцию в спинном мозге мышей несмотря на энергичную гуморальную иммунную реакцию, сопровождающуюся хронической демиелинизацией. Вирус представляет собой превосходную модель для изучения множественного склероза человека, при котором также наблюдается демиелинизация. Большинство клеток, инфицированных вирусом, производит 100—500 копий генома, что намного меньше, чем необходимо для определяемой экспрессии вирусных белков. Ограниченный синтез вирусных белков ведет к уменьшению цитопатического эффекта. Анализ последовательностей мутантных вирусов, устойчивых к нейтрализующим МКА к БД идентифицировал 4 аминокислотных мутации капсидных белков и все они локализовались на рецепторе или вблизи связывающего вирус. Мутантные вирусы показали уменьшение демиелинизирующей активности. Эти находки предполагают, что нейтрализация антител зависит от связывания вируса с клеточным рецептором и что демиелинизация и вирусная персистенция могут интерферировать с предотвращением прикрепления вируса к рецепторам особых типов клеток. Белок 17 кД (L-белок) синтезируется в штаммах субгруппы DA. Изучение вирусного мутанта DA без L-npo-теинового синтеза показало, что этот белок является важным для вирусного роста в особых типах клеток и является критическим для демиелинизирующей болезни и вирусной персистенции. Инактивация гена рецептора у-интерферона делает мышей I29/SV чувствительными к персистентнОой инфекции и ДБ, а инактивация гена у-интерферона у мышей 157ВL/6 придает им чувствительность к персистентной инфекции, а не к клинической ДБ. Эти различия фенотипа обусловлены различиями генетического фона. Полное секвенирование генома в потомстве F2 из двух скрещиваний показало, что ДБ является результатом двух независимых факторов: высокой вирусной нагрузки и локуса Tmevd5 в хромосоме 11.
Морфология, химический состав и устойчивость. РНК-содержащий вирус Тейлора имеет размеры 20—25 нм, устойчив в диапазоне рН 3—10, а также к эфиру, хлороформу, фенолу, гипертоническому раствору NaCl, глицерину, алюминиевым квасцам. Разрушается под воздействием 20 %-ного раствора этилового спирта, 50 %-ного ацетона, 2 %-ного раствора КМп04, 1,5 %-ного раствора формалина. Длительно сохраняется при низких температурах (-40 ° — 70 °С), неустойчив к прогреванию (50 °-70 °С) и высокочувствителен к лиофильному высушиванию. АГ структура не изучена. У больных мышей старше года обнаруживают ВНА, ПА, анти-ГА.
Штаммы ТО, FA и GD-VII родственны, но не идентичны. ВЭМТ индуцирует ген c-fos в чистых культурах покоящихся астроцитов мозга мыши. Экспрессия c-fos зависит от дозы ВЭМТ и достигает максимума при множественности заражения 100. Экспрессия c-fos максимальная через 30 мин и исчезает через 2 ч. Методом проточной цитометрии показано, что ВЭМТ быстро интернализируется после связывания со специфическим рецептором и активно реплицируется в цитоплазме астроцитов и что мРНК c-fos может транслироваться в ядре. Индукция c-fos нейтрализуется антителами к ВЭМТ. Очищенные белки VP1, VP2 и VP3 ВЭМТ не индуцируют экспрессию ВЭМТ. Быстрая индукция c-fos может являться первой стадией заражения астроцитов ВЭМТ.
Изучение структуры ВЭМТ позволило предположить, что глубокая впадина на поверхности вирионов участвует в прикреплении вируса к рецептору. Для проверки этого предположения у менее вирулентного варианта ВеАп ВЭМТ сконструировали замены четырех аминокислотных остатков (V1091, Р1153, А1225 и Р1379). В каждом из этих сайтов сделаны три аминокислотные замены: одна консервативная и две более радикальные. Получены семь жизнеспособных мутантов вируса и изучены их связывание с клетками ВНК-21, стабильность капсидов при 40 °, репликация РНК, кинетика роста в одиночном и множественных циклах заражения и трансляция вирусных белков. Три из указанных остатков участвуют в прикреплении вируса к клеточным рецепторам.
Шт. GD-VII в отличие от других (FA и ТО) агглютинирует эритроциты человека группы 0 при температуре 4 °С до разведения выше 1:2000. После обработки трипсином ГА-активность штамма повышается и проходит при температуре 20 °С с эритроцитами человека и 4 °С с эритроцитами обезьяны. После обработки трипсином два других штамма (FA и ТО) также приобретают ГА-свойства при температуре 20 °С.
Латентно инфицированные мыши выделяют вирус с фекалиями в течение 53 дней.
Экспериментальная инфекция. Экспериментальная инфекция шт. ТО воспроизводится на мышах 3—4-недельного возраста при интрацеребральном, интраназальном, интраперитонеальном и внутримышечном введениях (гибель при первом методе введения в 100 % случаев). Взрослые мыши не заболевают или легко переболевают и становятся вирусоносителями. Вирус проникает В ЦНС и другие органы через 2—5 дней после заражения. Он локализуется, в основном, в стенке кишечника и оттуда проникает в мезентериальные лимфоузлы и далее в ЦНС. У зараженных мышей старшего возраста может развиваться прогрессирующий вялый паралич, у еще более старших — латентная инфекция. После первичного выделения шт. GD-VII обычно вызывает вялый паралич, после пассирования на мышах — генерализованный энцефалит. Шт. FA также постоянно вызывает только диффузный энцефалит. Вялые параличи отмечают только после интраперитонеальной инокуляции.
Внутримышечная инокуляция любого из трех штаммов вызывает локальный миозит, по крайней мере шт. GD-VII, видимо, размножается в мышцах. Получена серия мутантов штамма DA со специфическими мутациями вирусного капсидного белка VP1, определяющего связь вируса с рецепторами клетки. Мутант вируса с заменой в петле IJ/PJ треонина на аспарагин (T81D) формирует крупные бляшки, но обладает пониженной способностью к репликации in vitro. При инъекции этого мутанта восприимчивым мышам выявляется измененный тропизм в острой стадии заболевания и не развивается хроническое демиелинизирующее заболевание. При замене треонина на валин (T81V) вирус не вызывает изменения, характерные для заболевания мышей. При замене в этой позиции триптофана (T81W) развивается сходное острое заболевание, но не развивается хроническая болезнь. Изменение аминокислот в гидрофобной области вдавления стенки, в позиции 91 VP1, в гидрофильный треонин (V91T), приводит к острому или хроническому заболеванию в отсутствие персистен-ции вируса. Таким образом, замены аминокислот в области петли I VP1 вируса приводят к изменению взаимодействия вируса с хозяином.
Культивирование. Вирус FA размножается в КЭ, зараженных на ХАО, в желточный мешок или в аллантоисную полость. Кроме эмбрионов, вирус размножается в культуре фибробластов КЭ, в культуре клеток печени и почек мышей, а также в ВНК-21. Особенности внутриклеточной репродукции не изучены. Шт. DA вызывает в клетках L929 персистенцию, которая обусловлена продукцией интерферона. Синтез РНК В клетках ВНК-21, зараженных шт. GDVI1 Или DA снижается через 3 ч после заражения, достигает максимального уровня через 6 ч, затем снижается. Изучена чувствительность первичных культур нейронов штаммами DAh GDVTI ВЭМТ. Высоковирулентный штамм GDVII заражает более 90 % Нейронов через 12 ч после инокуляции, а через 24 час. после заражения вызывает сильный цитопатический эффект. Антигены ВЭМТ обнаруживаются только через 24 ч после заражения вызывающим хрони-ческеую демиелинизирующую болезнь штаммом DA, который инфицирует не более 2 % нейронов даже через 6 дней. Заражение штаммом DA не вызывает цитопатичес-кого эффекта. С использованием рекомбинантных ВЭМТ показано, что капсид действительно отвечает за картину заражения, наблюдаемую in vitro. Таким образом, уровень нейровирулентности штаммов DA и GDVII коррелируете их способностью заражать культуры нейронов и контролируется капсидом. Штамм GD VII не индуцирует наработку полипептида вследствии его плохого размножения в мышиных L-клетках.
Источники и пути передачи инфекции. Источником инфекции при спонтанном энцефалите служат больные мыши, выделяющие вирус во внешнюю среду с калом и мочой, а также мыши-вирусоносители. Они являются основным резервуаром вирусов в природе. Заражаются мыши в основном алиментарным путем, через инфицированный экскрементами корм. Передавать вирус здоровым животным могут кровососущие насекомые.
Кроме белых мышей, восприимчивы серые дикие мыши, хлопковые крысы. У морских свинок удавалось вызвать латентную инфекцию. Кролики, белые и водяные крысы, лесные мыши и обезьяны к вирусу резистентны. На ранней стадии инфииирования мышей штаммом Даниеля вируса энцефаломиелита мышей вирус обнаруживается в нейронах и глии. В белом веществе вирус персистирует дольше, тогда как под воздействием иммунной системы происходит его ранняя элиминация из серого вещества. Из рыжей полевки выделен неизвестный ранее пикорнавирус, названный вирусом Люген. Принадлежность к пикорнавирусам была установлена на основании данных ЭМ и секвенирования. Рассчитанные аминокислотные последовательности капсидных белков VP2 и VP3 вируса Люген были сходны с таковыми вируса ЕСНО-22 (сходство около 70 %). 5'-некодирующая область РНК вируса Люген характеризовалась наибольшим сходством с кардиовирусами.
Иммунитет и специфическая профилактика. Иммунитет не изучен, специфическая профилактика не разработана. Новорожденные мышата и мышата-сосунки приобретают пассивный иммунитет с материнскими AT. Вскоре после отъема они становятся вирусоносителями и обладают повышенной резистентностью к инфекции. С возрастом процент вирусоносителей уменьшается: в 7,5 месяца он составлял всего 50 % и в дальнейшем понижался. Инфекция мышей вирусом энцефаломиелита Тейлора напоминает по клиническим проявлениям рассеянный склероз (PC) человека. Сконструированы экспрессирующие эукариотические векторы, кодирующие капсидные белки VP1, VP2 и VP3. Вакцинировали мышей SJL/J внутримышечно однократно, дважды или трехкратно кДНК различных капсидных белков. Затем мышей заражали в мозг. Трехкратная вакцинация мышей кДНК, кодирующей VP2 вируса, приводит к частичной защите их ЦНС от демиелинизирующего заболевания: уменьшались клинические симптомы и гистопатология. Вакцинация с помощью кДНК, кодирующей VP3, также приводила к уменьшению клинических проявлений. В то же время, вакцинация кДНК, кодирующей VP1, вызывала более тяжелое заболевание, причем симптомы заболевания появлялись раньше, а гистопатология усиливалась в сравнении с контролем. Это не коррелировало с титрами антител против и течением болезни. Таким образом, вакцинация с помощью ДНК способна изменять течение хронического демиелинизирующего заболевания. Это может найти применение в лечении PC.
Африканская чума однокопытных
[Pferdepest, Pestis equorum (лат.); African horse sicknes, Equine plaque (англ.); Africanische Pferdesterbe (нем.); Peste equine (франц.); Peste equina (исп.)]
Африканская чума однокопытных (африканская чума лошадей) (АЧЛ) — вирусная болезнь, протекающая остро и подостро, характеризуется лихорадкой, отеками подкожной клетчатки и кровоизлияниями во внутренних органах. Болезнь относится к группе трансмиссивных инфекций, передается кровососущими насекомыми, носит сезонный характер, проявляясь в теплое, влажное время года (7).
Вирусную природу болезни впервые установили Александер и др. в 1934 г.
До 1958 г. АЧЛ была распространена, в основном, в Африке. Начиная с 1959 г. инфекцию начали регистрировать в странах Ближнего и Среднего Востока. Осенью 1966 г. ее диагностировали в Испании. Экономический ущерб от этой инфекции слагается из падежа животных и затрат, связанных с проведением противоэпизоотических мероприятий.
Клинические признаки и патологоанатомические изменения. Различают лихорадочную, легочную и сердечную, или отечную, формы болезни. Описана также смешанная форма АЧЛ.
Лихорадочная форма протекает сверхостро и характеризуется высокой температурой, конъюнктивитом, учащенным дыханием и ускоренным пульсом. Погибают животные на 5—7-й день после появления клинических признаков.
Легочная форма болезни протекает остро и регистрируется в период эпизоотии среди животных, очень чувствительных к инфекции. После инкубационного периода резко повышается температура тела, дыхание становится затрудненным, появляются одышка, сухой болезненный кашель и желтоватые истечения из носа. Погибают больные животные быстро.
Сердечная или отечная форма протекает подостро и отличается сильным отеком головы и шеи. Инкубационный период продолжается до 20 дней. Наблюдается расстройство сердечной деятельности. Почти все больные животные погибают. Патологоанатомические изменения соответствуют форме болезни. При легочной форме отмечают отечность легких и скопление жидкости в плевральной полости, а также инфильтрацию соединительной ткани у основания сердца. При разрезе легких из ткани выделяется желтоватая пенистая жидкость. При сердечной форме наблюдают отечность соединительной подкожной, мышечной тканей и лимфоузлов. Отечность охватывает голову, шею и доходит иногда до грудной клетки и плечевых суставов. Иногда у павших лошадей бывает отек век, височных впадин, губ и межчелюстного пространства.
Морфология и химический состав. Вирионы АЧЛ характеризуются икосаэдр&ть-ным типом симметрии, диаметр их 70—80 нм, капсомеров 32. Капсид вириона состоит из одного слоя. Вирус содержит 2-нитевую РНК. В градиенте сахарозы удается разделить ее на пять различных по размеру фрагментов. РНК вируса состоит из 10 сегментов, которые кодируют шесть структурных белков. Неструктурные белки кодируются сегментами 6, 8 и 10. Сегмент 5 кодирует неструктурный белок NS1. Показана высокая консервативность гена NS1, который рекомендуют использовать для гибридизации in situ с диагностической целью. У вируса АЧЛ наружный капсидныи слой вириона образован полипептидами VP3 и VP5, остальные полипептиды (VP1, VP2, VP7) находятся в структуре нуклеокапсида. Белок VP2 — главный белок вируса АЧЛ, наиболее доступный на поверхности вириона. Главный сердцевинный белок VP7 вируса АЧЛ серотипа 4 экспрессирован в клетках насекомых Sf, зараженных вирусом ядерного полиэдроза Autographa californica. Сердцевинный белок VP7 может быть использован для выявления AT к различным АГ вируса АЧЛ.
Установлено АГ-родство между вирусами АЧЛ и другими орбивирусами: (КЛО и ЭГБО. Методом ЭФ в ПААГ с додецилсульфатом натрия анализировали структурные и неструктурные белки, индуцированные в клетках Vero серотипом 4 вируса АЧЛ. Выявлены 22 вирусспецифических полипептида. Методами иммуноблотинга и радиоиммунологически идентифицировано четыре больших (VP2, VP3, VP5 и VP7) и три малых (VP1, VP4, VP6) структурных белка и четыре неструктурных белка (Р58, Р48, Р21 и Р20). Наибольший иммунный ответ вызывают вирионные белки VP2, VP5 и VP7 и четыре основных неструктурных белка. При сравнении в перекрестных серологических реакциях серотипа 4 вируса АЧЛ и семь других серотипов показано, что АГ VP5, VP7 и Р48, Р21 и Р21 консервативны. Они могут быть использованы для диагностики всех 8 изученных серотипов вируса АЧЛ.
Нейротропные штаммы вируса инактивируются за 5—15 мин при 60 "С. Штаммы, адаптированные к культуре клеток, сохраняются при 4 °С до 90 дней. При хранении в культуральной среде при температуре от -20 * до -30 °С вирулентность вируса снижается. Инактивацию вируса при -22 °С можно предотвратить добавлением 5 % лактозы, сахарозы или глюкозы. В лиофилизированном виде вируссодержащий материал сохраняет инфекционную активность в течение нескольких лет. Вируссодержащая кровь больных лошадей, смешанная в равном объеме с раствором оксалатфенол-глицерина, при 4 °С Остается инфекционной несколько лет. Нейротропные штаммы вируса, выращенные в культуре клеток MS и сохраняемые при 37 °С, утрачивают свою активность через 40 дней. При хранении в условиях 25 "С титр вируса в том же материале за тот же срок снижается на 1—2 lg, а при 4 °С инфекционная активность суспензии сохраняется в течение 40 дней. Лиофилизированная полиштаммная вакцина в условиях 4 "С сохраняет активность в течение 9 месяцев и лишь к 18 месяцам хранения теряет 2 lg своего исходного титра.
Инактивация вируса связана с присутствием в среде NaCl, CaCb, MgCh. В растворах без этих солей вирус длительно сохраняется при -20 "С. Он стабилен в щелочной зоне с рН 7,0—8,0, устойчив к действию эфира и быстро инактивируется при рН 6,0. Чувствителен к УФ-лучам, причем степень чувствительности различных в АГ-отно-шении штаммов вируса неодинакова. На этом свойстве основана методика их дифференциации штаммов АЧЛ. Штаммы, идентичные в АГ-отношении, одинаково чувствительны к УФ лучам. Формалин инактивирует вирус в концентрации до 1:8000. 50 %-ный раствор глицерина используют как стабилизатор вируса, а 5—10 %-ный сапонин — как инактиватор.
У естественно переболевших и искусственно зараженных лошадей, мулов, а также зараженных морских свинок, кроликов и овец вырабатываются ВНА, КСА и ПА. Кролики дают высокоактивную сыворотку, пригодную для типирования отдельных штаммов вируса в РН. Положительную сыворотку для РСК (контрольную) получают от искусственно зараженных лошадей. Кровь берут через 30 дней после заражения. Для получения АГ мышат заражают в мозг.
Существует 10 серотипов вируса АЧЛ, различающихся в перекрестной РН, РТГА и иммунитете, и один общий КС АГ. Лошади, переболевшие чумой, становятся невосприимчивыми только к вирусу, вызвавшему заболевание. Прототипными штаммами выделенных АГ типов являются следующие: I-A501, 11-ОД, 1II-L, lV-Vryh, V-VH, VI-114, VII-Karen, VIII-18/60, IX-7/6. Различие в АГ структуре вируса носит скорее количественный, чем качественный характер. Штаммы, принадлежащие к различным типам, содержат одинаковые АГ компоненты, но в различных пропорциях. При серийном пассировании на лошадях вирус претерпевает иммунологические изменения, и лошадь, иммунизированная против определенного штамма, может не иметь иммунитета при заражении ее этим же штаммом, но прошедшим определенное число пассажей на лошадях.
Висцеротропные эпизоотические и нейротропные вакцинные штаммы вируса АЧЛ агглютинируют 0,5 % эритроциты лошади при рН 6,4.
У больных лошадей вирус содержится во всех органах и тканях. В крови естественно больных лошадей вирус достигает титра 2—3 ig ЛД 50/мл - Однако в наибольшем количестве он находится и в легких павших животных (до 3—4 lg ЛД so/мл). У отдельных переболевших лошадей отмечается вирусоносительство и вирусовыделение в течение 3 месяцев.
Экспериментальная инфекция. Неиммунных лошадей удается заразить, если вводить вирус подкожно, внутрикожно, внутривенно, интратрахеально, интрапульмонально, внутрибрюшинно и per os. При внутривенном введении вируссодержащего материала в объеме 5 мл на 4—5-е сутки у лошади повышается температура тела, а на 10—15-е сутки она погибает.
Культивирование. Вирус АЧЛ размножается в организме лошадей, мулов, морских свинок, белых мышей, КЭ, а также в первичных культурах клеток почки хомяка, ягнят, в фибробластах КЭ, в перевиваемых клетках MS (клетки почки обезьяны), ВНК-21 (клетки почки хомяка), LMK (клетки почки молодой обезьяны) и Vero. Перед заражением культур клеток почки ягнят и эмбрионов овец удается выделять и культивировать вирус, выделенный из крови и органов больных и погибших от чумы животных. Пассирование вируса в тканевых культурах также сопровождается его аттенуацией с сохранением АГ и иммуногенных свойств.
Нейротропные вирусы размножаются в мозге КЭ, сохраняя свои нейротропные свойства, но не вызывая гибели эмбрионов. Висцеротропные штаммы вызывают гибель КЭ и с увеличением числа пассажей аттенуируются. Титр вируса в зараженных эмбрионах достигает максимума на 4—5-й день после заражения, затем быстро снижается. Наиболее пригодными для культивирования вируса АЧЛ оказались клетки линии ППК (почки поросенка, казанская линия). Установлена возможность использования монослойных и суспензионных клеток ВНК-21 для изготовления вирус-вакцины против АЧЛ. Показана также возможность выращивания вируса АЧЛ различных серотипов в суспензии клеток ППК-666/17 в объемах 10—20 л при использовании питательной среды с гидролизатом белков крови КРС. В ходе пассирования вируса в течение 5 пассажей способность к репродукции и АГ специфичность вируса сохранялись. В инфицированных клетках MS и Vero вирус образует включения через 24 ч после заражения.
Источники и пути передачи инфекции. Источник инфекции — больные животные. Основная роль в переносе вируса от больных животных здоровым принадлежит мокрецам из рода Culicoides. Мокрецы (С. nubeculosis и С. variipennis) способны репродуцировать вирус АЧЛ серотипа 9. Однако вопросы, связанные с длительностью переживания вируса в мокрецах, возможностью его трансовариальной передачи, а также с ослаблением или усилением патогенности при передаче насекомыми не изучены. Не исключена возможность участия в переносе вируса комаров Anopheles stephensi и Culex pipiens. Экспериментально доказана возможность заражения здоровых лошадей комарами (Aedes egypti), накормленными кровью больных лошадей.
Возможно заражение собак при поедании конины, зараженной вирусом АЧЛ. В межэпизоотические периоды вирус, по-видимому, поддерживается в организме каких-то животных или птиц. АЧЛ является эндемичной инфекцией в прилегающих к пустыне Сахара странах. В передаче вируса могут также участвовать москиты и мухи. Периодически вирус АЧЛ распространяется за пределы энзоотических зон, но до настоящего времени он был нештатен, сохраняясь вне этих зон более 2—3 лет. Это так же обусловлено отсутствием достаточного количества позвоночных, способностью к распространению и сезонностью существования переносчиков, а также проведением эффективных мер контроля. Недавние эпизоотии на Пиренейском полуострове и в Северной Африке, как оказалось, привели к новому способу существования вируса АЧЛ. Вероятно, в этих регионах он связан с постоянным присутствием Culicoides imicola, которые предпочитают теплый климат. В связи с этим, его постоянное присутствие на Пиренейском полуострове может быть обусловлено изменением климата в этом районе. В 1987—1991 гг. наблюдали вспышки АЧЛ в Испании, Португалии и Марокко. Изменения климатических условий могут привести к дальнейшему распространению на север Culicoides imicola и превращению Европы в зону риска по АЧЛ, а также КЛО.
Мокрецов 17 видов, собранных в естественных условиях в ЮАР, в лабораторных условиях кормили суспензией вируса Африканской болезни лошадей (ВАБЛ) в крови через мембрану. В работе использовали ВАБЛ серотипов 3, 5 и 8 (соответственно ВАБЛ-3, -5, -8). ВАБЛ был выявлен через 10 дней после питания и содержания при 23,5 ° С у мокрецов Culicoides imicola при их кормлении ВАБЛ-5 (8,5 %) и ВАБЛ-8 (26,8 %), а также C. bolitinus после кормления ВАБЛ-8 (1,7 %). Показано, что сразу после питания 44,4 % С. imicola содержали ВАБЛ, однако через 10 дней это число сократилось до указанных выше цифр. У остальных 15 видов мокрецов ВАБЛ выявлен не был. Полученные результаты, особенно касающиеся мокрецов C. bolitinos, важны в связи с вопросами эпизоотологии Африканской болезни лошадей в ЮАР
В естественных условиях наиболее восприимчивы лошади, мулы менее чувствительны, ослы не заболевают даже при экспериментальном заражении. Гибель лошадей зависит от породы и вирулентности возбудителя.
Иммунитет и специфическая профилактика. Переболевшие чумой животные нечувствительны к вирусу того типа, который вызвал заболевание, но восприимчивы к вирусу других типов. Реконвалесценты приобретают иммунитет несколько лет. Новорождённые жеребята не имеют AT. Они появляются после первого кормления материнским молоком. В дальнейшем титр гуморальных AT у жеребят, полученных от матери, постепенно снижается и к 5-6 месяцам угасает.
Специфическую профилактику осуществляют инактивированными и живыми вакцинами. Из инактивированных чаще применяют формолвакцину, в которой в качестве АГ используется вируссодержащая селезёнка больной лошади. Вакцина не вызывает осложнений, и иммуногенность её достаточно надёжна. Успешно испытана инактивированная димером этиленимина вакцина. Показана возможность стимуляции иммунитета при использовании полиштаммных вирусвакцин. Установлена возможность использования монослойных и суспензионных культур клеток ВН К для изготовления вакцины против АЧЛ. Получены индикаторные для морских свинок штаммы вируса серотипов 2 и 6, пригодные для изучения иммунитета на вирусвакцину против АЧЛ и перспективные для разработки более экономичных методов контроля. На основании оценки степени нейропатогенности вируса АЧЛ по индексу церебральной патогенности и нейропатогенной активности, конкурирующих с иммуногенностью вакцины, разработана технологическая схема изготовления моно - и полиштаммных вакцин против АЧЛ и поддержания вакцинных штаммов посредством чередования (ограниченного до 10 пассажей) культивирования вируса в клетках П П К с освежением его на мышах. Схема включает процедуры контроля вакцинного материала и допускает хранение готовой вакцины до 5 лет при -40 °С.
Живую нейтропную вакцину готовят из мозга мышей, заражённых аттенуированными штаммами. Лошади, привитые полиштаммной мозговой мышиной вакциной, приобретают иммунитет к 10—14-му дню и остаются невосприимчивы в течение нескольких лет. Помимо мышиной мозговой вакцины, в ветеринарной практике некоторых стран используют вакцину из мозга морских свинок. Вакцина ареактогенна и иммуногенна. Современная живая вакцина получена из аттенуированного вируса, выращенного в культуре перевиваемых клеток MS или ВНК-21. В районах, неблагополучных по АЧЛ, животных вакцинируют за 1—2 месяца до появления кровососущих насекомых. Иммунитет развивается через 15—20 дней после вакцинации и сохраняется не менее года.
Отсутствие AT к вирусу АЧЛ после первичного применения вакцины не обязательно свидетельствует об отсутствии защиты.
Африканская чума однокопытных
[Pferdepest, Pestis equorum (лат.); African horse sicknes, Equine plaque (англ.); Africanische Pferdesterbe (нем.); Peste equine (франц.); Peste equina (исп.)]
Африканская чума однокопытных (африканская чума лошадей) (АЧЛ) — вирусная болезнь, протекающая остро и подостро, характеризуется лихорадкой, отеками подкожной клетчатки и кровоизлияниями во внутренних органах. Болезнь относится к группе трансмиссивных инфекций, передается кровососущими насекомыми, носит сезонный характер, проявляясь в теплое, влажное время года (7).
Вирусную природу болезни впервые установили Александер и др. в 1934 г.
До 1958 г. АЧЛ была распространена, в основном, в Африке. Начиная с 1959 г. инфекцию начали регистрировать в странах Ближнего и Среднего Востока. Осенью 1966 г. ее диагностировали в Испании. Экономический ущерб от этой инфекции слагается из падежа животных и затрат, связанных с проведением противоэпизоотических мероприятий.
Клинические признаки и патологоанатомические изменения. Различают лихорадочную, легочную и сердечную, или отечную, формы болезни. Описана также смешанная форма АЧЛ.
Лихорадочная форма протекает сверхостро и характеризуется высокой температурой, конъюнктивитом, учащенным дыханием и ускоренным пульсом. Погибают животные на 5—7-й день после появления клинических признаков.
Легочная форма болезни протекает остро и регистрируется в период эпизоотии среди животных, очень чувствительных к инфекции. После инкубационного периода резко повышается температура тела, дыхание становится затрудненным, появляются одышка, сухой болезненный кашель и желтоватые истечения из носа. Погибают больные животные быстро.
Сердечная или отечная форма протекает подостро и отличается сильным отеком головы и шеи. Инкубационный период продолжается до 20 дней. Наблюдается расстройство сердечной деятельности. Почти все больные животные погибают. Патологоанатомические изменения соответствуют форме болезни. При легочной форме отмечают отечность легких и скопление жидкости в плевральной полости, а также инфильтрацию соединительной ткани у основания сердца. При разрезе легких из ткани выделяется желтоватая пенистая жидкость. При сердечной форме наблюдают отечность соединительной подкожной, мышечной тканей и лимфоузлов. Отечность охватывает голову, шею и доходит иногда до грудной клетки и плечевых суставов. Иногда у павших лошадей бывает отек век, височных впадин, губ и межчелюстного пространства.
Морфология и химический состав. Вирионы АЧЛ характеризуются икосаэдр&ть-ным типом симметрии, диаметр их 70—80 нм, капсомеров 32. Капсид вириона состоит из одного слоя. Вирус содержит 2-нитевую РНК. В градиенте сахарозы удается разделить ее на пять различных по размеру фрагментов. РНК вируса состоит из 10 сегментов, которые кодируют шесть структурных белков. Неструктурные белки кодируются сегментами 6, 8 и 10. Сегмент 5 кодирует неструктурный белок NS1. Показана высокая консервативность гена NS1, который рекомендуют использовать для гибридизации in situ с диагностической целью. У вируса АЧЛ наружный капсидныи слой вириона образован полипептидами VP3 и VP5, остальные полипептиды (VP1, VP2, VP7) находятся в структуре нуклеокапсида. Белок VP2 — главный белок вируса АЧЛ, наиболее доступный на поверхности вириона. Главный сердцевинный белок VP7 вируса АЧЛ серотипа 4 экспрессирован в клетках насекомых Sf, зараженных вирусом ядерного полиэдроза Autographa californica. Сердцевинный белок VP7 может быть использован для выявления AT к различным АГ вируса АЧЛ.
Установлено АГ-родство между вирусами АЧЛ и другими орбивирусами: (КЛО и ЭГБО. Методом ЭФ в ПААГ с додецилсульфатом натрия анализировали структурные и неструктурные белки, индуцированные в клетках Vero серотипом 4 вируса АЧЛ. Выявлены 22 вирусспецифических полипептида. Методами иммуноблотинга и радиоиммунологически идентифицировано четыре больших (VP2, VP3, VP5 и VP7) и три малых (VP1, VP4, VP6) структурных белка и четыре неструктурных белка (Р58, Р48, Р21 и Р20). Наибольший иммунный ответ вызывают вирионные белки VP2, VP5 и VP7 и четыре основных неструктурных белка. При сравнении в перекрестных серологических реакциях серотипа 4 вируса АЧЛ и семь других серотипов показано, что АГ VP5, VP7 и Р48, Р21 и Р21 консервативны. Они могут быть использованы для диагностики всех 8 изученных серотипов вируса АЧЛ.
Нейротропные штаммы вируса инактивируются за 5—15 мин при 60 "С. Штаммы, адаптированные к культуре клеток, сохраняются при 4 °С до 90 дней. При хранении в культуральной среде при температуре от -20 * до -30 °С вирулентность вируса снижается. Инактивацию вируса при -22 °С можно предотвратить добавлением 5 % лактозы, сахарозы или глюкозы. В лиофилизированном виде вируссодержащий материал сохраняет инфекционную активность в течение нескольких лет. Вируссодержащая кровь больных лошадей, смешанная в равном объеме с раствором оксалатфенол-глицерина, при 4 °С Остается инфекционной несколько лет. Нейротропные штаммы вируса, выращенные в культуре клеток MS и сохраняемые при 37 °С, утрачивают свою активность через 40 дней. При хранении в условиях 25 "С титр вируса в том же материале за тот же срок снижается на 1—2 lg, а при 4 °С инфекционная активность суспензии сохраняется в течение 40 дней. Лиофилизированная полиштаммная вакцина в условиях 4 "С сохраняет активность в течение 9 месяцев и лишь к 18 месяцам хранения теряет 2 lg своего исходного титра.
Инактивация вируса связана с присутствием в среде NaCl, CaCb, MgCh. В растворах без этих солей вирус длительно сохраняется при -20 "С. Он стабилен в щелочной зоне с рН 7,0—8,0, устойчив к действию эфира и быстро инактивируется при рН 6,0. Чувствителен к УФ-лучам, причем степень чувствительности различных в АГ-отно-шении штаммов вируса неодинакова. На этом свойстве основана методика их дифференциации штаммов АЧЛ. Штаммы, идентичные в АГ-отношении, одинаково чувствительны к УФ лучам. Формалин инактивирует вирус в концентрации до 1:8000. 50 %-ный раствор глицерина используют как стабилизатор вируса, а 5—10 %-ный сапонин — как инактиватор.
У естественно переболевших и искусственно зараженных лошадей, мулов, а также зараженных морских свинок, кроликов и овец вырабатываются ВНА, КСА и ПА. Кролики дают высокоактивную сыворотку, пригодную для типирования отдельных штаммов вируса в РН. Положительную сыворотку для РСК (контрольную) получают от искусственно зараженных лошадей. Кровь берут через 30 дней после заражения. Для получения АГ мышат заражают в мозг.
Существует 10 серотипов вируса АЧЛ, различающихся в перекрестной РН, РТГА и иммунитете, и один общий КС АГ. Лошади, переболевшие чумой, становятся невосприимчивыми только к вирусу, вызвавшему заболевание. Прототипными штаммами выделенных АГ типов являются следующие: I-A501, 11-ОД, 1II-L, lV-Vryh, V-VH, VI-114, VII-Karen, VIII-18/60, IX-7/6. Различие в АГ структуре вируса носит скорее количественный, чем качественный характер. Штаммы, принадлежащие к различным типам, содержат одинаковые АГ компоненты, но в различных пропорциях. При серийном пассировании на лошадях вирус претерпевает иммунологические изменения, и лошадь, иммунизированная против определенного штамма, может не иметь иммунитета при заражении ее этим же штаммом, но прошедшим определенное число пассажей на лошадях.
Висцеротропные эпизоотические и нейротропные вакцинные штаммы вируса АЧЛ агглютинируют 0,5 % эритроциты лошади при рН 6,4.
У больных лошадей вирус содержится во всех органах и тканях. В крови естественно больных лошадей вирус достигает титра 2—3 ig ЛД 50/мл - Однако в наибольшем количестве он находится и в легких павших животных (до 3—4 lg ЛД so/мл). У отдельных переболевших лошадей отмечается вирусоносительство и вирусовыделение в течение 3 месяцев.
Экспериментальная инфекция. Неиммунных лошадей удается заразить, если вводить вирус подкожно, внутрикожно, внутривенно, интратрахеально, интрапульмонально, внутрибрюшинно и per os. При внутривенном введении вируссодержащего материала в объеме 5 мл на 4—5-е сутки у лошади повышается температура тела, а на 10—15-е сутки она погибает.
Культивирование. Вирус АЧЛ размножается в организме лошадей, мулов, морских свинок, белых мышей, КЭ, а также в первичных культурах клеток почки хомяка, ягнят, в фибробластах КЭ, в перевиваемых клетках MS (клетки почки обезьяны), ВНК-21 (клетки почки хомяка), LMK (клетки почки молодой обезьяны) и Vero. Перед заражением культур клеток почки ягнят и эмбрионов овец удается выделять и культивировать вирус, выделенный из крови и органов больных и погибших от чумы животных. Пассирование вируса в тканевых культурах также сопровождается его аттенуацией с сохранением АГ и иммуногенных свойств.
Нейротропные вирусы размножаются в мозге КЭ, сохраняя свои нейротропные свойства, но не вызывая гибели эмбрионов. Висцеротропные штаммы вызывают гибель КЭ и с увеличением числа пассажей аттенуируются. Титр вируса в зараженных эмбрионах достигает максимума на 4—5-й день после заражения, затем быстро снижается. Наиболее пригодными для культивирования вируса АЧЛ оказались клетки линии ППК (почки поросенка, казанская линия). Установлена возможность использования монослойных и суспензионных клеток ВНК-21 для изготовления вирус-вакцины против АЧЛ. Показана также возможность выращивания вируса АЧЛ различных серотипов в суспензии клеток ППК-666/17 в объемах 10—20 л при использовании питательной среды с гидролизатом белков крови КРС. В ходе пассирования вируса в течение 5 пассажей способность к репродукции и АГ специфичность вируса сохранялись. В инфицированных клетках MS и Vero вирус образует включения через 24 ч после заражения.
Источники и пути передачи инфекции. Источник инфекции — больные животные. Основная роль в переносе вируса от больных животных здоровым принадлежит мокрецам из рода Culicoides. Мокрецы (С. nubeculosis и С. variipennis) способны репродуцировать вирус АЧЛ серотипа 9. Однако вопросы, связанные с длительностью переживания вируса в мокрецах, возможностью его трансовариальной передачи, а также с ослаблением или усилением патогенности при передаче насекомыми не изучены. Не исключена возможность участия в переносе вируса комаров Anopheles stephensi и Culex pipiens. Экспериментально доказана возможность заражения здоровых лошадей комарами (Aedes egypti), накормленными кровью больных лошадей.
Возможно заражение собак при поедании конины, зараженной вирусом АЧЛ. В межэпизоотические периоды вирус, по-видимому, поддерживается в организме каких-то животных или птиц. АЧЛ является эндемичной инфекцией в прилегающих к пустыне Сахара странах. В передаче вируса могут также участвовать москиты и мухи. Периодически вирус АЧЛ распространяется за пределы энзоотических зон, но до настоящего времени он был нештатен, сохраняясь вне этих зон более 2—3 лет. Это так же обусловлено отсутствием достаточного количества позвоночных, способностью к распространению и сезонностью существования переносчиков, а также проведением эффективных мер контроля. Недавние эпизоотии на Пиренейском полуострове и в Северной Африке, как оказалось, привели к новому способу существования вируса АЧЛ. Вероятно, в этих регионах он связан с постоянным присутствием Culicoides imicola, которые предпочитают теплый климат. В связи с этим, его постоянное присутствие на Пиренейском полуострове может быть обусловлено изменением климата в этом районе. В 1987—1991 гг. наблюдали вспышки АЧЛ в Испании, Португалии и Марокко. Изменения климатических условий могут привести к дальнейшему распространению на север Culicoides imicola и превращению Европы в зону риска по АЧЛ, а также КЛО.
Мокрецов 17 видов, собранных в естественных условиях в ЮАР, в лабораторных условиях кормили суспензией вируса Африканской болезни лошадей (ВАБЛ) в крови через мембрану. В работе использовали ВАБЛ серотипов 3, 5 и 8 (соответственно ВАБЛ-3, -5, -8). ВАБЛ был выявлен через 10 дней после питания и содержания при 23,5 ° С у мокрецов Culicoides imicola при их кормлении ВАБЛ-5 (8,5 %) и ВАБЛ-8 (26,8 %), а также C. bolitinus после кормления ВАБЛ-8 (1,7 %). Показано, что сразу после питания 44,4 % С. imicola содержали ВАБЛ, однако через 10 дней это число сократилось до указанных выше цифр. У остальных 15 видов мокрецов ВАБЛ выявлен не был. Полученные результаты, особенно касающиеся мокрецов C. bolitinos, важны в связи с вопросами эпизоотологии Африканской болезни лошадей в ЮАР
В естественных условиях наиболее восприимчивы лошади, мулы менее чувствительны, ослы не заболевают даже при экспериментальном заражении. Гибель лошадей зависит от породы и вирулентности возбудителя.
Иммунитет и специфическая профилактика. Переболевшие чумой животные нечувствительны к вирусу того типа, который вызвал заболевание, но восприимчивы к вирусу других типов. Реконвалесценты приобретают иммунитет несколько лет. Новорождённые жеребята не имеют AT. Они появляются после первого кормления материнским молоком. В дальнейшем титр гуморальных AT у жеребят, полученных от матери, постепенно снижается и к 5-6 месяцам угасает.
Специфическую профилактику осуществляют инактивированными и живыми вакцинами. Из инактивированных чаще применяют формолвакцину, в которой в качестве АГ используется вируссодержащая селезёнка больной лошади. Вакцина не вызывает осложнений, и иммуногенность её достаточно надёжна. Успешно испытана инактивированная димером этиленимина вакцина. Показана возможность стимуляции иммунитета при использовании полиштаммных вирусвакцин. Установлена возможность использования монослойных и суспензионных культур клеток ВН К для изготовления вакцины против АЧЛ. Получены индикаторные для морских свинок штаммы вируса серотипов 2 и 6, пригодные для изучения иммунитета на вирусвакцину против АЧЛ и перспективные для разработки более экономичных методов контроля. На основании оценки степени нейропатогенности вируса АЧЛ по индексу церебральной патогенности и нейропатогенной активности, конкурирующих с иммуногенностью вакцины, разработана технологическая схема изготовления моно - и полиштаммных вакцин против АЧЛ и поддержания вакцинных штаммов посредством чередования (ограниченного до 10 пассажей) культивирования вируса в клетках П П К с освежением его на мышах. Схема включает процедуры контроля вакцинного материала и допускает хранение готовой вакцины до 5 лет при -40 °С.
Живую нейтропную вакцину готовят из мозга мышей, заражённых аттенуированными штаммами. Лошади, привитые полиштаммной мозговой мышиной вакциной, приобретают иммунитет к 10—14-му дню и остаются невосприимчивы в течение нескольких лет. Помимо мышиной мозговой вакцины, в ветеринарной практике некоторых стран используют вакцину из мозга морских свинок. Вакцина ареактогенна и иммуногенна. Современная живая вакцина получена из аттенуированного вируса, выращенного в культуре перевиваемых клеток MS или ВНК-21. В районах, неблагополучных по АЧЛ, животных вакцинируют за 1—2 месяца до появления кровососущих насекомых. Иммунитет развивается через 15—20 дней после вакцинации и сохраняется не менее года.
Отсутствие AT к вирусу АЧЛ после первичного применения вакцины не обязательно свидетельствует об отсутствии защиты.
Читайте также: