
Большая медицинская энциклопедия вирусы
Обновлено: 19.04.2024
Вирусология (вирус[ы] + греч. logos учение) — учение о вирусах; самостоятельная область биологии, развивающая знания о субмикроскопических внутриклеточных паразитах, поражающих людей, животных, растения и микроорганизмы. К этой же области биологии относится учение о вирусах бактерий.
Вирусология занимает значительное место в биологии и медицине, т. к. вирусы вызывают многие заболевания людей, животных, растений, поражают плесневые грибы, простейшие организмы и бактерии, а также в связи с тем, что на модели вирусов изучаются основные проблемы генетики и молекулярной биологии.
История
Основоположник Вирусологии — русский ученый Д. И. Ивановский. Изучая мозаичную болезнь табака и использовав при этом метод фильтрации, он установил в 1892 г., что фильтрат из растертой взвеси листьев, пораженных этой болезнью, не содержал видимых в микроскоп микроорганизмов, однако вызывал типичные признаки мозаичной болезни у здоровых растений. На основании этих опытов Ивановский сделал вывод, что мозаичная болезнь табака вызывается мельчайшими микроорганизмами, проходящими через керамические фильтры, задерживающие все известные в то время бактерии, что они не способны расти на искусственных питательных средах, применяемых в бактериологии, и передаются в серии последовательных пассажей (прививок). В 1902 г. Ивановский обнаружил кристаллические включения в клетках табачных растений, пораженных мозаичной болезнью, в дальнейшем другими учеными было подтверждено, что это скопление вирусных частиц.
Использование метода фильтрации позволило в дальнейшем установить прохождение через керамические фильтры возбудителей других известных заболеваний человека и животных: ящура [Ф. Леффлер и Фрош (P. Frosch), 1898], желтой лихорадки [Рид (W. Reed, 1901) с сотр.]. В 1911 г. Ф. Раус доказал вирусную этиологию саркомы кур, т. е. впервые экспериментально установил, что вирусы могут вызывать неопластические процессы.
Для изучения вирусов, поражающих животных и растения, использовались в качестве модели соответствующие виды животных и растений. Для изучения и выделения вирусов, вызывающих заболевания человека, применялись восприимчивые к этому вирусу лабораторные животные (мыши, крысы, морские свинки, кролики, хорьки и т. д.). Широко использовались приемы введения различного инфекционного материала в роговицу глаза, кожу, мозг, дыхательные пути, а также принцип повторных пассажей на различных видах животных. Так, используя экспериментальных животных, выделили и изучили вирусы бешенства, оспы, герпеса, ящура, гриппа, энцефалитов, полиомиелита, хориоменингита и др. Однако к концу 30-х годов возможности этого метода были исчерпаны, т. к. не удавалось выделить многие вирусы, к к-рым экспериментальные животные были невосприимчивы, или нельзя было получить большого количества вирусов, очищенных от тканевых элементов, и в высоких концентрациях.
В 1931 г. был предложен метод культивирования вирусов на 8—13-дневном курином эмбрионе Вудраффом (М. F. Woodruff) и Э. Гудпасчером. В 40-х годах метод получил широкое распространение в вирусологии, т. к. имел ряд преимуществ: простота применения, большая чувствительность, возможность накопления большого количества вируса, относительная герметичность, предохраняющая от контаминации, относительная простота очистки от примесей, возможность быстрого определения наличия вируса в жидкостях эмбриона по данным реакции гемагглютинации.
Методом культивирования в курином эмбрионе (в клетках амниотической оболочки, в отдельных органах зародыша и клетках желточного мешка) были изучены вирусы гриппа человека и животных, чумы птиц, коровьей оспы, герпеса человека, энцефаломиелита лошадей и др. Эндерс, Роббинс, Уэллер (J. F. Enders, F. С. Robbins, Т. H. Weller, 1948—1952) применили для выделения и изучения вирусов метод культур клеток и тканей. Этот метод стал широко использоваться в различных вирусологических исследованиях и за несколько лет обогатил науку не только открытием сотен неизвестных ранее вирусов, но расширил возможности производства более качественных вирусных вакцин и диагностических препаратов; метод тканевых культур открыл новые возможности изучения различных аспектов и этапов процесса взаимодействия вируса и клетки (см. Культивирование вирусов, Культуры клеток и тканей).
Дальнейший прогресс В., и в частности изучение структуры, физиологии, биохимии и генетики вирусов, зависел от получения их в концентрированном и очищенном виде и был связан с внедрением новых физ.-хим. методов исследования: дифференциального и градиентного центрифугирования, молекулярно-адсорбционной и ионообменной хроматографии, электрофореза на бумаге и в полиакриламидном геле, радиоактивных изотопов и ряда других.
Быстрый прогресс В. был обусловлен применением электронных микроскопов с высокой разрешающей способностью (до 1,0—0,5 нм, в сочетании с методами оттенения и двойного оттенения, ультратонких срезов, позитивного и негативного контрастирования, а также авторадиографии, цитохимических и иммунохимических методов исследования. Использование комплекса перечисленных методов позволило изучить структурную организацию вирионов различных вирусов, предложить новую классификацию вирусов, основанную на их строении и биохимическом составе, изучить закономерности репродукции вирусов и определить детали их онтогенеза, охарактеризовать основные параметры субвирусных компонентов (нуклеиновых кислот, белков и др.), начать углубленные исследования по генетике вирусов и приступить к разработке рациональных подходов к химиотерапии вирусных инфекций.
Развитие Вирусологии способствовало изучению и решению общебиологических проблем: доказательству генетической функции нуклеиновых кислот, расшифровке генетического кода, пониманию важнейших механизмов регуляции синтеза клеточных макромолекул, установлению передачи информации от клетки к клетке и др.
Практическое здравоохранение получило ряд надежных вакцин для специфической профилактики не только оспы, что было известно еще задолго до рождения В. как науки, но и желтой лихорадки, полиомиелита, кори; появились новые средства для неспецифического воздействия на вирусные инфекции, напр, интерферон (см.).
Основные направления современной вирусологии
Основные направления современной общей и мед. вирусологии: дальнейшее изучение тонкой структуры вирусов, их биохимии и генетики, репликации вирусных нуклеиновых кислот, взаимодействия вируса с клеткой, углубленное изучение противовирусного иммунитета, совершенствование методов выделения вирусов и диагностики вирусных заболеваний, разработка основ химиотерапии и химиопрофилактики вирусных инфекций; изучение экологии вирусов, разработка более совершенных методов профилактики, поиски и испытание препаратов для лечения вирусных заболеваний.
Особое внимание будет сосредоточено на изучении вирусов, вызывающих неопластические процессы, а также латентных вирусных инфекций и скрытого вирусного носительства, поисках возбудителей инфекционного и сывороточного гепатита, разработке профилактики гриппа.
В 30-х годах в СССР были созданы первые вирусологические лаборатории: по изучению вирусов растений— при Украинском институте защиты растений (1930), по изучению вирусов животных — в Ин-те экспериментальной ветеринарии в Москве в 1930 г. (Н. Ф. Гамалея), Центральная вирусологическая лаборатория НКЗ РСФСР в Москве (Л. А. Зильбер) и отдел вирусологии в Ин-те эпидемиологии и микробиологии им. Л. Пастера в Ленинграде (А. А. Смородинцев) в 1935 г. В послевоенные годы в СССР созданы и функционируют профильные научно-исследовательские, научно-производственные и практические учреждения. По данным на 1-е января 1973 г., в СССР исследования по общей и мед. В. проводились в 60 научных, научно-производственных учреждениях и учебных заведениях. Наиболее значительные: Ин-т вирусологии им. Д. И. Ивановского АМН СССР, Ин-т полиомиелита и вирусных энцефалитов АМН СССР, Ин-т эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР, Ин-т экспериментальной и клинической онкологии АМН СССР, Ин-т молекулярной биологии АН СССР, Ин-т микробиологии АН СССР, Всесоюзный ин-т гриппа М3 СССР, Московский научно-исследовательский ин-т вирусных препаратов М3 СССР, Свердловский научно-исследовательский ин-т вирусных инфекций М3 РСФСР, Ин-т вирусологии и микробиологии АН Украинской ССР, Одесский научно-исследовательский ин-т вирусологии и эпидемиологии им. И. И. Мечникова М3 Украинской ССР, Институт инфекционных болезней М3 Украинской ССР, Ин-т микробиологии им. А. Кирхенштейна АН Латвийской ССР; во всех научно-исследовательских ин-тах микробиологии и эпидемиологии союзных республик созданы вирусологические лаборатории и отделы.
Наиболее крупные зарубежные учреждения, проводящие научные исследования по общей и мед. В.: National Institute for Medical Research (Лондон), National Communicable Disease Centre (Атланта, США), National Institute of Health (Токио), National Institute of Health (Бетесда, США), Institute of Epidemiology and Microbiology (Прага), Institute of Virology (Братислава), Institute Pasteur (Париж), Institute Inframicrobiology (Бухарест), Institute of Virology (Глазго, Англия), State Institute of Hygiene (Будапешт), Virus Research Centre (Пуна, Индия), Queensland Institute of Medical Research (Брисбейн, Австралия).
Результаты научных исследований по общей и мед. В. публикуются в следующих научных журналах: Доклады АН СССР (Москва), Бюллетень экспериментальной биологии и медицины (Москва), Вопросы вирусологии (Москва), Журнал микробиологии, эпидемиологии и иммунологии (Москва), Вестник АМН СССР (Москва), Archiv fur die gesamte Virusforschung (Вена), Acta Virologica (Прага), Virology (Нью-Йорк), Ann. Institute Pasteur (Париж), Revue Romanine de Virologie (Бухарест), Inter. Journal of Cancer (Хельсинки), Journal of Virology (Вашингтон), Advances Virus Research (Питтсбург, США), Journal of the National Cancer Institute (Бетесда, США), Intervirology (Берн).
Первые монографии по вирусологии: Rivers Т., Filterable Viruses, Baltimore, 1928; Hauduroy P., Les Ultra Virus, Paris, 1929; Гамалея H. Ф. Фильтрующиеся вирусы, М., 1930.
Результаты научных исследований по В. обсуждаются на конференциях, сессиях, проводимых профильными ин-тами, а также на международных конгрессах.
В СССР первая научная конференция по вирусным болезням растений состоялась в марте 1935 г. в Харькове, первая научная конференция по ультрамикробам, фильтрующимся вирусам и бактериофагам — в декабре 1935 г. в Москве. В 1966 г. на 9-м Международном конгрессе по микробиологии впервые был избран Международный комитет по номенклатуре вирусов.
1-й Международный конгресс по В. состоялся в 1968 г. в Хельсинки, 2-й — в 1971 г. в Будапеште (был принят устав секции вирусологов, учрежденной в рамках Международной ассоциации микробиологов), 3-й в 1975 г. в Мадриде.
Развитие В. привело к открытию новых вирусов, количество которых быстро возрастало, в связи с чем создавались коллекции вирусов — музеи, где хранились вирусы, выделенные как в данной стране, так и полученные из других стран. Наиболее крупные коллекции вирусов: в СССР (Москва, Ин-т вирусологии АМН СССР) — Государственная коллекция вирусов, основана в 1956 г. как филиал Всесоюзного музея живых культур и условнопатогенных микроорганизмов; в США (Вашингтон) — коллекция вирусов и риккетсий, основана в 1959 г. на базе коллекции типовых культур (American type culture collection, Washington 7, Rockville, Maryland, USA); в ЧССР (Прага, Ин-т эпидемиологии и микробиологии) — Чехословацкая национальная коллекция типовых культур, основана в 1969 г. (Czechoslovak National collection of type cultures of the Institute Epidemiology and Microbiology, Prague); в Японии (Токио) — Японская коллекция культур микроорганизмов, основана в 1962 г. (The Japanes Federation of Culture collection of Microorganisms, Tokyo, Japan); в Англии (Лондон) — каталог национальной коллекции типовых культур, основан в 1936 г. (Medical Research Council, Catalog of the National collection of Type cultures, London, England); в Швейцарии (Лозанна, Международный центр живых культур) имеется международный каталог вирусов.
Преподавание Вирусологии в медицинских вузах СССР проводится кафедрами микробиологии на II и III курсах, а по вирусным инфекциям лекции и клинические занятия проводят кафедры инфекционных болезней на V курсе.
На биол, ф-тах Московского и Киевского ун-тов созданы в течение последних 10 лет кафедры В., где готовят специалистов-вирусологов и ведется преподавание В. в течение одного семестра студентам других ф-тов.
Прогресс мед. В. в СССР сопровождался ростом числа специалистов высокой квалификации: с 1946 по 1960 г. подготовлено 16 докт, наук, с 1961 по 1972 г.— 140, кандидатов наук соответственно 217 и 836 (из них 54% путем обучения в аспирантуре). Важное значение в подготовке кадров вирусологов (специализация и усовершенствование) сыграла созданная в 1955 г. кафедра В. при ЦИУ, к-рая подготовила с октября 1955 г. по 1964 г.— 688 специалистов, а с 1965 г. по январь 1974 г.— 933, гл. обр. для обеспечения вирусологической работы в сан.-эпид, станциях.
Библиография: Авакян А. А. и Быковский А. Ф. Атлас анатомии и онтогенеза вирусов человека и животных, М., 1970, библиогр.; Бешенство, под ред. В. Д. Соловьева, М., 1954, библиогр.; Гаврилов В. И., Семенов Б. Ф. и Жданов В. М. Хронические вирусные инфекции и их моделирование, М., 1974, библиогр.; Гамалея Н. Ф. Фильтрующиеся вирусы, М.—Л., 1930; Гендон Ю. 3. Генетика вирусов человека и животных, М., 1967, библиогр .; Жданов В. М. и Гайдамович С. Я. Вирусология, М., 1966; Жданов В.М.,Соловьев В. Д. и Эпштейн Ф. Г. Учение о гриппе, М., 1958; Зильбер Л. А. Учение о вирусах (общая вирусология), М., 1956; Ивановский Д. И. О двух болезнях табака, Сельское хоз. и лесоводство, т. 169, № 2, с. 104, 1892; Косяков П. Н. и Ровнова 3. И. Противовирусный иммунитет, М., 1972; Морозов М. А. и Соловьев В. Д. Оспа, М., 1948; Першин Г. Н. и Богданова Н. С. Химиотерапия вирусных инфекций, М., 1973, библиогр.; С о-ловьев В. Д. Весенне-летний клещевой энцефалит, М., 1944, библиогр.; Соловьев В. Д. и Баландин PI. Г. Биохимические основы взаимодействия вируса и клетки, М., 1969, библиогр.; они же, Клетка и вирус, М., 1973, библиогр.; Соловьев В. Д. и Бектемиров Т. А. Интерферон в теории и практике медицины, М., 1970, библиогр.; Тихоненко Т. И. Биохимия вирусов, М., 1965, библиогр.; Шубладзе А. К. и Гайдамович С. Я. Краткий курс практической вирусологии, 2-е изд., М., 1954; Шубладзе А. К., Бычкова E. Н. и Баринский И. Ф. Вирусемия при острых и хронических инфекциях, М., 1974; Comprehen sive virology, ed. by H. Fraenkel-Conrat a. R. R. Wagner, v. 1 — 4, N. Y., 1974, bibliogr.; Starke G. u. HlinakP. Grundriss der allgemeinen Virologie, Jena, 1974, Bibliogr.
Периодические издания — Вопросы вирусологии, М., с 1956; Медицинский реферативный журнал, Раздел III — Медицинская микробиология, Вирусология, Медицинская паразитология, Эпидемиология, Инфекционные болезни, М., с 1960; Acta virologica, Praha, с 1957; Archiv fiir die gesamte Virusforschung, Wein, с 1939; Excerpta medica, Section IV — Microbiology, Bacteriology, Virology, Mycology and Parasitology, Amsterdam, с 1968; Journal of General Virology, L., с 1967; Journal of Virology, Baltimore , с 1969; Virology, N. Y., с 1955.
Вирусология (вирус[ы] + греч. logos учение) — учение о вирусах; самостоятельная область биологии, развивающая знания о субмикроскопических внутриклеточных паразитах, поражающих людей, животных, растения и микроорганизмы. К этой же области биологии относится учение о вирусах бактерий.
Вирусология занимает значительное место в биологии и медицине, т. к. вирусы вызывают многие заболевания людей, животных, растений, поражают плесневые грибы, простейшие организмы и бактерии, а также в связи с тем, что на модели вирусов изучаются основные проблемы генетики и молекулярной биологии.
История
Основоположник Вирусологии — русский ученый Д. И. Ивановский. Изучая мозаичную болезнь табака и использовав при этом метод фильтрации, он установил в 1892 г., что фильтрат из растертой взвеси листьев, пораженных этой болезнью, не содержал видимых в микроскоп микроорганизмов, однако вызывал типичные признаки мозаичной болезни у здоровых растений. На основании этих опытов Ивановский сделал вывод, что мозаичная болезнь табака вызывается мельчайшими микроорганизмами, проходящими через керамические фильтры, задерживающие все известные в то время бактерии, что они не способны расти на искусственных питательных средах, применяемых в бактериологии, и передаются в серии последовательных пассажей (прививок). В 1902 г. Ивановский обнаружил кристаллические включения в клетках табачных растений, пораженных мозаичной болезнью, в дальнейшем другими учеными было подтверждено, что это скопление вирусных частиц.
Использование метода фильтрации позволило в дальнейшем установить прохождение через керамические фильтры возбудителей других известных заболеваний человека и животных: ящура [Ф. Леффлер и Фрош (P. Frosch), 1898], желтой лихорадки [Рид (W. Reed, 1901) с сотр.]. В 1911 г. Ф. Раус доказал вирусную этиологию саркомы кур, т. е. впервые экспериментально установил, что вирусы могут вызывать неопластические процессы.
Для изучения вирусов, поражающих животных и растения, использовались в качестве модели соответствующие виды животных и растений. Для изучения и выделения вирусов, вызывающих заболевания человека, применялись восприимчивые к этому вирусу лабораторные животные (мыши, крысы, морские свинки, кролики, хорьки и т. д.). Широко использовались приемы введения различного инфекционного материала в роговицу глаза, кожу, мозг, дыхательные пути, а также принцип повторных пассажей на различных видах животных. Так, используя экспериментальных животных, выделили и изучили вирусы бешенства, оспы, герпеса, ящура, гриппа, энцефалитов, полиомиелита, хориоменингита и др. Однако к концу 30-х годов возможности этого метода были исчерпаны, т. к. не удавалось выделить многие вирусы, к к-рым экспериментальные животные были невосприимчивы, или нельзя было получить большого количества вирусов, очищенных от тканевых элементов, и в высоких концентрациях.
В 1931 г. был предложен метод культивирования вирусов на 8—13-дневном курином эмбрионе Вудраффом (М. F. Woodruff) и Э. Гудпасчером. В 40-х годах метод получил широкое распространение в вирусологии, т. к. имел ряд преимуществ: простота применения, большая чувствительность, возможность накопления большого количества вируса, относительная герметичность, предохраняющая от контаминации, относительная простота очистки от примесей, возможность быстрого определения наличия вируса в жидкостях эмбриона по данным реакции гемагглютинации.
Методом культивирования в курином эмбрионе (в клетках амниотической оболочки, в отдельных органах зародыша и клетках желточного мешка) были изучены вирусы гриппа человека и животных, чумы птиц, коровьей оспы, герпеса человека, энцефаломиелита лошадей и др. Эндерс, Роббинс, Уэллер (J. F. Enders, F. С. Robbins, Т. H. Weller, 1948—1952) применили для выделения и изучения вирусов метод культур клеток и тканей. Этот метод стал широко использоваться в различных вирусологических исследованиях и за несколько лет обогатил науку не только открытием сотен неизвестных ранее вирусов, но расширил возможности производства более качественных вирусных вакцин и диагностических препаратов; метод тканевых культур открыл новые возможности изучения различных аспектов и этапов процесса взаимодействия вируса и клетки (см. Культивирование вирусов, Культуры клеток и тканей).
Дальнейший прогресс В., и в частности изучение структуры, физиологии, биохимии и генетики вирусов, зависел от получения их в концентрированном и очищенном виде и был связан с внедрением новых физ.-хим. методов исследования: дифференциального и градиентного центрифугирования, молекулярно-адсорбционной и ионообменной хроматографии, электрофореза на бумаге и в полиакриламидном геле, радиоактивных изотопов и ряда других.
Быстрый прогресс В. был обусловлен применением электронных микроскопов с высокой разрешающей способностью (до 1,0—0,5 нм, в сочетании с методами оттенения и двойного оттенения, ультратонких срезов, позитивного и негативного контрастирования, а также авторадиографии, цитохимических и иммунохимических методов исследования. Использование комплекса перечисленных методов позволило изучить структурную организацию вирионов различных вирусов, предложить новую классификацию вирусов, основанную на их строении и биохимическом составе, изучить закономерности репродукции вирусов и определить детали их онтогенеза, охарактеризовать основные параметры субвирусных компонентов (нуклеиновых кислот, белков и др.), начать углубленные исследования по генетике вирусов и приступить к разработке рациональных подходов к химиотерапии вирусных инфекций.
Развитие Вирусологии способствовало изучению и решению общебиологических проблем: доказательству генетической функции нуклеиновых кислот, расшифровке генетического кода, пониманию важнейших механизмов регуляции синтеза клеточных макромолекул, установлению передачи информации от клетки к клетке и др.
Практическое здравоохранение получило ряд надежных вакцин для специфической профилактики не только оспы, что было известно еще задолго до рождения В. как науки, но и желтой лихорадки, полиомиелита, кори; появились новые средства для неспецифического воздействия на вирусные инфекции, напр, интерферон (см.).
Основные направления современной вирусологии
Основные направления современной общей и мед. вирусологии: дальнейшее изучение тонкой структуры вирусов, их биохимии и генетики, репликации вирусных нуклеиновых кислот, взаимодействия вируса с клеткой, углубленное изучение противовирусного иммунитета, совершенствование методов выделения вирусов и диагностики вирусных заболеваний, разработка основ химиотерапии и химиопрофилактики вирусных инфекций; изучение экологии вирусов, разработка более совершенных методов профилактики, поиски и испытание препаратов для лечения вирусных заболеваний.
Особое внимание будет сосредоточено на изучении вирусов, вызывающих неопластические процессы, а также латентных вирусных инфекций и скрытого вирусного носительства, поисках возбудителей инфекционного и сывороточного гепатита, разработке профилактики гриппа.
В 30-х годах в СССР были созданы первые вирусологические лаборатории: по изучению вирусов растений— при Украинском институте защиты растений (1930), по изучению вирусов животных — в Ин-те экспериментальной ветеринарии в Москве в 1930 г. (Н. Ф. Гамалея), Центральная вирусологическая лаборатория НКЗ РСФСР в Москве (Л. А. Зильбер) и отдел вирусологии в Ин-те эпидемиологии и микробиологии им. Л. Пастера в Ленинграде (А. А. Смородинцев) в 1935 г. В послевоенные годы в СССР созданы и функционируют профильные научно-исследовательские, научно-производственные и практические учреждения. По данным на 1-е января 1973 г., в СССР исследования по общей и мед. В. проводились в 60 научных, научно-производственных учреждениях и учебных заведениях. Наиболее значительные: Ин-т вирусологии им. Д. И. Ивановского АМН СССР, Ин-т полиомиелита и вирусных энцефалитов АМН СССР, Ин-т эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР, Ин-т экспериментальной и клинической онкологии АМН СССР, Ин-т молекулярной биологии АН СССР, Ин-т микробиологии АН СССР, Всесоюзный ин-т гриппа М3 СССР, Московский научно-исследовательский ин-т вирусных препаратов М3 СССР, Свердловский научно-исследовательский ин-т вирусных инфекций М3 РСФСР, Ин-т вирусологии и микробиологии АН Украинской ССР, Одесский научно-исследовательский ин-т вирусологии и эпидемиологии им. И. И. Мечникова М3 Украинской ССР, Институт инфекционных болезней М3 Украинской ССР, Ин-т микробиологии им. А. Кирхенштейна АН Латвийской ССР; во всех научно-исследовательских ин-тах микробиологии и эпидемиологии союзных республик созданы вирусологические лаборатории и отделы.
Наиболее крупные зарубежные учреждения, проводящие научные исследования по общей и мед. В.: National Institute for Medical Research (Лондон), National Communicable Disease Centre (Атланта, США), National Institute of Health (Токио), National Institute of Health (Бетесда, США), Institute of Epidemiology and Microbiology (Прага), Institute of Virology (Братислава), Institute Pasteur (Париж), Institute Inframicrobiology (Бухарест), Institute of Virology (Глазго, Англия), State Institute of Hygiene (Будапешт), Virus Research Centre (Пуна, Индия), Queensland Institute of Medical Research (Брисбейн, Австралия).
Результаты научных исследований по общей и мед. В. публикуются в следующих научных журналах: Доклады АН СССР (Москва), Бюллетень экспериментальной биологии и медицины (Москва), Вопросы вирусологии (Москва), Журнал микробиологии, эпидемиологии и иммунологии (Москва), Вестник АМН СССР (Москва), Archiv fur die gesamte Virusforschung (Вена), Acta Virologica (Прага), Virology (Нью-Йорк), Ann. Institute Pasteur (Париж), Revue Romanine de Virologie (Бухарест), Inter. Journal of Cancer (Хельсинки), Journal of Virology (Вашингтон), Advances Virus Research (Питтсбург, США), Journal of the National Cancer Institute (Бетесда, США), Intervirology (Берн).
Первые монографии по вирусологии: Rivers Т., Filterable Viruses, Baltimore, 1928; Hauduroy P., Les Ultra Virus, Paris, 1929; Гамалея H. Ф. Фильтрующиеся вирусы, М., 1930.
Результаты научных исследований по В. обсуждаются на конференциях, сессиях, проводимых профильными ин-тами, а также на международных конгрессах.
В СССР первая научная конференция по вирусным болезням растений состоялась в марте 1935 г. в Харькове, первая научная конференция по ультрамикробам, фильтрующимся вирусам и бактериофагам — в декабре 1935 г. в Москве. В 1966 г. на 9-м Международном конгрессе по микробиологии впервые был избран Международный комитет по номенклатуре вирусов.
1-й Международный конгресс по В. состоялся в 1968 г. в Хельсинки, 2-й — в 1971 г. в Будапеште (был принят устав секции вирусологов, учрежденной в рамках Международной ассоциации микробиологов), 3-й в 1975 г. в Мадриде.
Развитие В. привело к открытию новых вирусов, количество которых быстро возрастало, в связи с чем создавались коллекции вирусов — музеи, где хранились вирусы, выделенные как в данной стране, так и полученные из других стран. Наиболее крупные коллекции вирусов: в СССР (Москва, Ин-т вирусологии АМН СССР) — Государственная коллекция вирусов, основана в 1956 г. как филиал Всесоюзного музея живых культур и условнопатогенных микроорганизмов; в США (Вашингтон) — коллекция вирусов и риккетсий, основана в 1959 г. на базе коллекции типовых культур (American type culture collection, Washington 7, Rockville, Maryland, USA); в ЧССР (Прага, Ин-т эпидемиологии и микробиологии) — Чехословацкая национальная коллекция типовых культур, основана в 1969 г. (Czechoslovak National collection of type cultures of the Institute Epidemiology and Microbiology, Prague); в Японии (Токио) — Японская коллекция культур микроорганизмов, основана в 1962 г. (The Japanes Federation of Culture collection of Microorganisms, Tokyo, Japan); в Англии (Лондон) — каталог национальной коллекции типовых культур, основан в 1936 г. (Medical Research Council, Catalog of the National collection of Type cultures, London, England); в Швейцарии (Лозанна, Международный центр живых культур) имеется международный каталог вирусов.
Преподавание Вирусологии в медицинских вузах СССР проводится кафедрами микробиологии на II и III курсах, а по вирусным инфекциям лекции и клинические занятия проводят кафедры инфекционных болезней на V курсе.
На биол, ф-тах Московского и Киевского ун-тов созданы в течение последних 10 лет кафедры В., где готовят специалистов-вирусологов и ведется преподавание В. в течение одного семестра студентам других ф-тов.
Прогресс мед. В. в СССР сопровождался ростом числа специалистов высокой квалификации: с 1946 по 1960 г. подготовлено 16 докт, наук, с 1961 по 1972 г.— 140, кандидатов наук соответственно 217 и 836 (из них 54% путем обучения в аспирантуре). Важное значение в подготовке кадров вирусологов (специализация и усовершенствование) сыграла созданная в 1955 г. кафедра В. при ЦИУ, к-рая подготовила с октября 1955 г. по 1964 г.— 688 специалистов, а с 1965 г. по январь 1974 г.— 933, гл. обр. для обеспечения вирусологической работы в сан.-эпид, станциях.
Библиография: Авакян А. А. и Быковский А. Ф. Атлас анатомии и онтогенеза вирусов человека и животных, М., 1970, библиогр.; Бешенство, под ред. В. Д. Соловьева, М., 1954, библиогр.; Гаврилов В. И., Семенов Б. Ф. и Жданов В. М. Хронические вирусные инфекции и их моделирование, М., 1974, библиогр.; Гамалея Н. Ф. Фильтрующиеся вирусы, М.—Л., 1930; Гендон Ю. 3. Генетика вирусов человека и животных, М., 1967, библиогр .; Жданов В. М. и Гайдамович С. Я. Вирусология, М., 1966; Жданов В.М.,Соловьев В. Д. и Эпштейн Ф. Г. Учение о гриппе, М., 1958; Зильбер Л. А. Учение о вирусах (общая вирусология), М., 1956; Ивановский Д. И. О двух болезнях табака, Сельское хоз. и лесоводство, т. 169, № 2, с. 104, 1892; Косяков П. Н. и Ровнова 3. И. Противовирусный иммунитет, М., 1972; Морозов М. А. и Соловьев В. Д. Оспа, М., 1948; Першин Г. Н. и Богданова Н. С. Химиотерапия вирусных инфекций, М., 1973, библиогр.; С о-ловьев В. Д. Весенне-летний клещевой энцефалит, М., 1944, библиогр.; Соловьев В. Д. и Баландин PI. Г. Биохимические основы взаимодействия вируса и клетки, М., 1969, библиогр.; они же, Клетка и вирус, М., 1973, библиогр.; Соловьев В. Д. и Бектемиров Т. А. Интерферон в теории и практике медицины, М., 1970, библиогр.; Тихоненко Т. И. Биохимия вирусов, М., 1965, библиогр.; Шубладзе А. К. и Гайдамович С. Я. Краткий курс практической вирусологии, 2-е изд., М., 1954; Шубладзе А. К., Бычкова E. Н. и Баринский И. Ф. Вирусемия при острых и хронических инфекциях, М., 1974; Comprehen sive virology, ed. by H. Fraenkel-Conrat a. R. R. Wagner, v. 1 — 4, N. Y., 1974, bibliogr.; Starke G. u. HlinakP. Grundriss der allgemeinen Virologie, Jena, 1974, Bibliogr.
Периодические издания — Вопросы вирусологии, М., с 1956; Медицинский реферативный журнал, Раздел III — Медицинская микробиология, Вирусология, Медицинская паразитология, Эпидемиология, Инфекционные болезни, М., с 1960; Acta virologica, Praha, с 1957; Archiv fiir die gesamte Virusforschung, Wein, с 1939; Excerpta medica, Section IV — Microbiology, Bacteriology, Virology, Mycology and Parasitology, Amsterdam, с 1968; Journal of General Virology, L., с 1967; Journal of Virology, Baltimore , с 1969; Virology, N. Y., с 1955.
мельчайшие микроорганизмы, не имеющие клеточного строения, белоксинтезирующей системы и способные к воспроизведению лишь в клетках высокоорганизованных форм жизни. Они широко распространены в природе, поражают животных, растения и другие микроорганизмы. В. характеризуются рядом уникальных свойств, отличающих их от простейших, грибков, бактерий — микроорганизмов, имеющих клеточное строение и генетический материал, представленный двунитчатыми ДНК. Вирусы не имеют рибосом и цитоплазматических органелл, их воспроизводство обеспечивает клетка-хозяин. Молекула вирусного генома наделена необычайной способностью перестраивать жизнедеятельность клетки таким образом, что она перестает узнавать собственную генетическую информацию и функционирует в соответствии с генетической программой вируса, синтезируя вирусоспецифические молекулы. С этой точки зрения В. являются генетическими паразитами клетки.
Вирусы содержат нуклеиновую кислоту только одного типа: либо ДНК, либо РНК. РНК-содержащие В. — единственные представители в природе, имеющие генетический материал, представленный РНК. Вирусные геномы гаплоидны, т.е. содержат только одну копию генов, за исключением ретровирусов, геном которых является диплоидным. Генетический материал может иметь вид разнообразных структур (двунитчатых, однонитчатых, линейных, кольцевых, фрагментированных молекул). В основе необычного способа воспроизводства В. лежит разобщенный во времени и пространстве (на территории клетки) синтез вирусных нуклеиновых кислот и белков, которые затем независимо друг от друга прибывают к местам сборки вирусных частиц.
Природу В. как генетических паразитов клетки характеризует их способность к интеграции, т.е. к объединению вирусного генома с клеточным. Группа вирусных генов, являющихся частью клеточного генома, называется провирусом. Провирус способен длительное время существовать в виде так называемых молчащих генов, однако в соответствующих условиях он может активироваться, что приводит к развитию болезни. На способности В. к интеграции основан механизм персистенции В. в организме, с которой связано возникновение персистентных вирусных инфекций. Интеграция характерна для умеренных ДНК-содержащих бактериофагов, онкогенных ДНК-содержащих вирусов, для вируса гепатита В, обязательна для ретровирусов, к которым относятся онкогенные РНК-содержащие В. и вирусы иммунодефицита человека. Персистенция В. в организме возможна также при существовании их в клетке в виде кольцевых нуклеиновых кислот типа плазмид бактериальной клетки, реплицируемых самой клеткой. Такие кольцевые ДНК, лишенные собственных белков, описаны при персистенции паповавирусов, В. герпеса.
К вирусам близки вироиды и вирусоиды. Вироиды представляют собой небольшие по размерам молекулы кольцевой суперспирализованной РНК, лишенные белка: они вызывают болезни растений. Вирусоиды также являются лишенными белков молекулами РНК, они отличаются от вироидов зависимостью от вируса-помощника (дефектностью) и отсутствием способности кодировать синтез собственных белков.
Вирусы, вироиды, вирусоиды и даже прионы имеют нечто общее, их объединяющее. Они представляют собой автономные генетические структуры, способные функционировать и репродуцироваться в восприимчивых к ним клетках животных, растений и микроорганизмов.
Несмотря на чрезвычайную простоту строения, В. обладают всеми признаками жизни: способностью размножаться, наследственностью, изменчивостью, приспособляемостью к условиям окружающей среды, они занимают определенную экологическую нишу в природе, на них распространяются законы эволюции органического мира. Однако В. не являются организмами из-за отсутствия собственных белок-синтезирующих систем, разобщенного способа репродукции, способности интегрировать с клеточным геномом, наличия вироидов, прионов, вирусов-сателлитов и дефектных вирусов, генетических феноменов (множественной реактивации и комплементации).
Зрелая вирусная частица называется вирионом. У простых вирусов (например, вирусов полиомиелита) вирион состоит из одной молекулы нуклеиновой кислоты, окруженной белковым футляром — капсидом. Капсид вместе с нуклеиновой кислотой называется нуклеокапсидом. У более сложных вирусов (например, аденовирусов, ротавирусов) капсид окружает сердцевину, содержащую помимо нуклеиновой кислоты внутренние белки. У еще более сложно устроенных В. (вирусы гриппа, кори, бешенства, вирус иммунодефицита человека) капсид окружен липопротеиновой оболочкой — суперкапсидом (рис. 1). Капсид состоит из определенного количества капсомеров, связанных нековалентными связями, каждый из которых содержит несколько симметрично расположенных полипептидных цепей. Вирионы патогенных для человека В. имеют два типа симметрии. При спиральной симметрии капсомеры ассоциируются с геномом и образуют спиралевидную винтообразную структуру, при кубическом типе симметрии капсомеры формируют изометрическое полое тело, внутри которого находится геном Изометрические вирусные частицы имеют форму икосаэдра — многогранника, состоящего обычно из 60 или кратных 60 геометрически идентичных элементов.
Липопротеиновые оболочки В. являются дериватами плазматической мембраны или мембран цитоплазматических вакуолей, куда почкуются В. Поэтому липиды вирусной оболочки имеют такой же состав, как и липиды клетки-хозяина. В липопротеиновые оболочки В. встроены наружные вирусные белки, которые обычно представлены гликопротеидами. Эти белки формируют шипы на поверхности вирусной частицы, их функция связана с проникновением вируса в клетку. Наружные вирусные белки вызывают образование защитных, вирус-нейтрализующих антител.
Вирусные нуклеиновые кислоты характеризуются разнообразием форм. Вирусный геном может быть представлен как однонитчатыми, так и двунитчатыми молекулами РНК или ДНК, ДНК может быть как линейной, так и кольцевой молекулой, РНК — как линейной, так и фрагментированной, и вместе с белками нуклеокапсида образовывать кольцевые структуры В., содержащие однонитчатые РНК, делят на две группы. К одной группе относятся В., геном которых обладает свойствами информационной РНК, т.е. может связываться с рибосомами и кодировать вирусные белки. Такие В. обозначают как плюс-нитевые, или плюс-геномные (условно информационная РНК обозначена знаком плюс). Это пикорнавирусы, тогавирусы, коронавирусы, ретровирусы. Другая группа включает В., у которых функции информационной РНК выполняет РНК, комплементарная геному. Эти В. называют минус-нитевыми, или минус-геномными. Минус-нитевыми являются ортомиксовирусы, парамиксо-вирусы, рабдовирусы. Существуют В., содержащие как плюс-нитевые, так и минус-нитевые гены (амбисенс-вирусы). К ним относятся аренавирусы и буньявирусы.
В зараженной клетке вирусный геном кодирует синтез двух групп белков: структурных, входящих в состав образующихся вирионов, и неструктурных, которые обнаруживаются в зараженной клетке, но в состав вирионов не входят. Число структурных белков варьирует от 2—3 у простых В. до 100 и более у сложно организованных, например вирусов оспы. В зависимости от локализации в вирионе различают капсидные и суперкапсидные структурные белки. В составе капсида, помимо идентичных белков, образующих капсомеры, находится ряд других белков: геномные белки, ковалентно связанные с геномом, ферменты, осуществляющие транскрипцию и репликацию вирусного генома, белки с другими ферментативными функциями. Эти белки представлены в виде единичных молекул. Основной функцией собственно капсидных белков является защита генома от факторов внешней среды. Суперкапсидные белки (обычно гликопротеиды) относятся к типичным внутримембранным белкам. Их гликозилирование осуществляют клеточные ферменты в процессе синтеза и транспорта полипептида. Углеводный компонент защищает полипептид от протеаз, придает определенную конформацию молекуле, влияет на ее антигенные свойства. Все циркулирующие в природе В. принято делить на вирусы, поражающие позвоночных и человека, насекомых, растения и бактерии. Вирусы классифицируют на большие группы, называемые семействами (-viridae), которые подразделяют на подсемейства (-virinae), роды (genus) и типы. Основными критериями для характеристики семейства являются морфология вирионов, природа генома, репликация генома. Морфология вирионов включает размер, форму, симметрию нуклеокапсида, наличие суперкапсида. Деление на подсемейства и роды обусловлено такими критериями, как круг восприимчивых хозяев, патогенность, патологические изменения в клетках, феномены генетических взаимодействий, географическое распространение, способ передачи; наиболее частым признаком при делении на роды являются антигенные свойства (антигенные перекресты). Дифференциация на типы, подтипы, штаммы и варианты также основана на антигенных свойствах при использовании как поликлональных, так и моноклональных антител. Современная классификация охватывает 4 /5 всех известных В. позвоночных и человека, из них 6 семейств включают ДНК-содержащие вирусы и 12 — РНК-содержащие.
Некоторые свойства этих семейств приведены в таблице. Схематическое изображение строения В., патогенных для человека, показано на рис. 2.
Ви́рус (от лат. virus — яд) — микроскопическая частица, способная инфицировать клетки живых организмов. Вирусы являются облигатными паразитами — они не способны размножаться вне клетки. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружен также вирус, поражающий другие вирусы [1] . Вирусы представляют собой молекулы нуклеиновых кислот (ДНК или РНК), заключённые в защитную белковую оболочку (капсид). Наличие капсида отличает вирусы от других инфекционных агентов, вироидов. Вирусы содержат только один тип нуклеиновой кислоты: либо ДНК , либо РНК. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые белки и не содержат нуклеиновых кислот.
Содержание
Положение вирусов в системе живого
Вирусы являются одной из самых распространённых форм существования органической материи на планете по численности: воды мирового океана содержат колоссальное количество бактериофагов (около 10 11 частиц на миллилитр воды).
Вирусы имеют генетические связи с представителями флоры и фауны Земли. Согласно последним исследованиям, геном человека более чем на 32 % состоит из информации, кодируемой вирус-подобными элементами и транспозонами. С помощью вирусов может происходить так называемый горизонтальный перенос генов (ксенология), то есть передача генетической информации не от отца к сыну и так далее, а между двумя неродственными (или даже относящимися к разным видам) особями. Так в геноме высших приматов существует белок синцитин, который, как считается, был привнесён ретровирусом. Иногда вирусы образуют с животными симбиоз [2] . Так, например, яд некоторых паразитических ос содержит структуры, называемые поли-ДНК-вирусами (Polydnavirus, PDV), имеющие вирусное происхождение.
Происхождение вирусов
Вирусы — сборная группа, не имеющая общего предка. В настоящее время существует несколько гипотез, объясняющих происхождение вирусов.
Считается, что крупные ядерно-цитоплазматические ДНК-содержащие вирусы происходят от более сложных (и, возможно, клеточных, таких как современные микоплазмы и риккетсии), внутриклеточных паразитов, утративших значительную часть своего генома. И действительно, некоторые крупные ДНК-содержащие вирусы (мимивирус, вирус оспы) кодируют функционально избыточные на первый взгляд ферменты, по-видимому, оставшиеся им в наследство от более сложных форм существования. Следует также отметить, что некоторые вирусные белки не обнаруживают никакой гомологии с белками бактерий, архей и эукариот, что свидетельствует о сравнительно давнем обособлении этой группы.
ДНК-содержащие бактериофаги и некоторые ДНК-содержащие вирусы эукариот, возможно происходят от мобильных элементов, участков ДНК способных к самостоятельной репликации в клетке.
Структура

Примеры структур икосаэдрических вирионов.
А. Вирус, не имеющий липидной оболочки (например, пикорнавирус).
B. Оболочечный вирус (например, герпесвирус).
Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки.
Вирусные частицы (вирио́ны) представляют собой белковую капсулу — капсид, содержащую геном вируса, представленный одной или несколькими молекулами ДНК или РНК. Капсид построен из капсомеров — белковых комплексов, состоящих в свою очередь из протомеров. Нуклеиновая кислота в комплексе с белками обозначается термином нуклеокапсид. Некоторые вирусы имеют также внешнюю липидную оболочку. Размеры различных вирусов колеблются от 20 (пикорнавирусы) до 500 (мимивирусы) и более нанометров. Вирионы часто имеют правильную геометрическую форму (икосаэдр, цилиндр). Такая структура капсида предусматривает идентичность связей между составляющими её белками, и, следовательно, может быть построена из стандартных белков одного или нескольких видов, что позволяет вирусу экономить место в геноме.
Механизм инфицирования
Условно процесс вирусного инфицирования в масштабах одной клетки можно разбить на несколько взаимоперекрывающихся этапов:

Палочковидная частица вируса табачной мозаики.
Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида.
- Присоединение к клеточной мембране — так называемая адсорбция. Обычно для того, чтобы вирион адсорбировался на поверхности клетки, она должна иметь в составе своей плазматической мембраны белок (часто гликопротеин) — рецептор, специфичный для данного вируса. Наличие рецептора нередко определяет круг хозяев данного вируса, а также его тканеспецифичность.
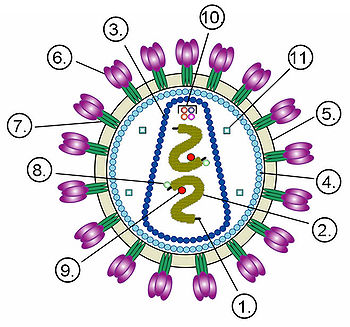
Структура вириона неикосаэдрического оболочечного вируса на примере ВИЧ.
Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — трансмембранный гликопротеин.
Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза.
Классификация
В таксономии живой природы вирусы выделяются в отдельный таксон Vira, образующий в классификации Systema Naturae 2000 вместе с доменами Bacteria, Archaea и Eukaryota корневой таксон Biota. В течение XX века в систематике выдвигались предложения о создании выделенного таксона для неклеточных форм жизни (Aphanobionta Novak, 1930; надцарство Acytota Jeffrey, 1971; Acellularia), однако такие предложения не были кодифицированы.
Систематику и таксономию вирусов кодифицирует и поддерживает Международный Комитет по Таксономии Вирусов (International Committee on Taxonomy of Viruses, ICTV), поддерживающий также и таксономическую базу The Universal Virus Database ICTVdB.
ICTV классификация
Международным Комитетом по Таксономии Вирусов в 1966 году была принята система классификации вирусов основанная на различии типа (РНК и ДНК), количества молекул нуклеотических кислот (одно- и двух-цепочечные) и на наличии или отсутствии оболочки ядра. Система классификации представляет собой серию иерархичных таксонов:
Отряд (-virales) Семейство (-viridae) Подсемейство (-virinae) Род (-virus) Вид (-virus)
Классификация Балтимора
Нобелевский лауреат, биолог Дэвид Балтимор, предложил свою схему классификации вирусов, основываясь на различиях в механизме продукции мРНК.Эта система включает в себя семь основных групп: [3] [4]
- (I) Вирусы, содержащие двуцепочечную ДНК и не имеющие РНК-стадии (например, герпесвирусы, поксвирусы, паповавирусы, мимивирус).
- (II) Вирусы, содержащие двуцепочечную РНК (например, ротавирусы).
- (III) Вирусы, содержащие одноцепочечную молекулу ДНК (например, парвовирусы).
- (IV) Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности (например, пикорнавирусы, флавивирусы).
- (V) Вирусы, содержащие одноцепочечную молекулу РНК негативной или двойной полярности (например, ортомиксовирусы, филовирусы).
- (VI) Вирусы, содержащие одноцепочечную молекулу РНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретровирусы (например, ВИЧ).
- (VII) Вирусы, содержащие двуцепочечную ДНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретроидные вирусы (например, вирус гепатита B).
В настоящее время, для классификации вирусов используются обе системы одновременно, как дополняющие друг друга. [5] [6] [7]
Дальнейшее деление производится на основе таких признаков как структура генома (наличие сегментов, кольцевая или линейная молекула), генетическое сходство с другими вирусами, наличие липидной оболочки, таксономическая принадлежнось организма-хозяина и так далее.
История
В 2002 в университете Нью-Йорка был создан первый синтетический вирус (вирус полиомиелита).
фильтрующиеся вирусы, ультравирусы, возбудители инфекционных болезней растений, животных и человека, размножающиеся только в живых клетках. В. мельче большинства известных микробов; почти все В. проходят через Бактериальные фильтры. В отличие от бактерий, В. не удаётся культивировать на обычных питательных средах. Для экспериментальных и медицинских целей (получения вакцин (См. Вакцина) и др.) В. культивируют в животных и растительных организмах, куриных эмбрионах и в культурах тканей (См. Культуры тканей) и клеток. В. вызывают многие заболевания: оспу, корь, грипп, полиомиелит, чуму рогатого скота и птиц, бешенство, ряд заболеваний рыб и земноводных, желтуху шелкопряда, мозаичную болезнь табака, закукливание овса, многие заболевания грибов и сине-зелёных водорослей и др. (см. Вирусные болезни, Вирусные болезни растений). Обширный отряд В., поражающих бактерии, составляют Бактериофаги.
Строение и состав частиц В. Форма вирионов очень разнообразна. У многих бактериофагов они состоят из головки и отростка, у В. оспы они прямоугольные, у В. герпеса и гриппа — шарообразные, у В. мозаичной болезни табака — палочкообразные, у В. мозаичной болезни картофеля — нитевидные, у В. полиомиелита и жёлтой мозаики турнепса — многогранные шарики, у В. бешенства, а также мозаики пшеницы и люцерны — очертаниями похожи на палочки бактерий или напоминают пулю. По размерам В. делят на крупные (300—400 нм в диаметре), средние (80—125 нм) и мелкие (20—30 нм). Крупные В. можно видеть в световой микроскоп (обычный, фазово-контрастный, люминесцентный); остальные изучают только с помощью электронного микроскопа. Данные о размерах частиц В. получены методами ультрафильтрации, фракционного и аналитического ультрацентрифугирования, электрофореза в гелях и электронной микроскопии (табл.).
* Разные авторы в зависимости от применяемых ими методов и др. условий получали величины, отклоняющиеся от приводимых, однако порядок величин во всех случаях сохраняется.
В строении разных вирионов есть много общего. Все они имеют белковую оболочку — капсид и внутреннее содержимое — нуклеокапсид, состоящее главным образом из нуклеиновой кислоты (См. Нуклеиновые кислоты) (НК) — ДНК или РНК. Многие В. имеют поверхностную оболочку, покрывающую белковую. Отдельные элементы белковой оболочки называются капсомерами. У некоторых В. (например, мозаичной болезни табака) НК в виде спирали включена в белковую оболочку, без разрушения которой не может быть освобождена. У других В. (например, жёлтой мозаики турнепса) спирально закрученная нить НК лежит в капсиде, как в коробочке, и может выйти оттуда без разрушения оболочки. НК — носители наследственной информации о строении и свойствах В.; белки В. защищают НК, а также обусловливают ферментативные и антигенные свойства В. (см. Антигены, Ферменты). Строение вирусных частиц, приспособленных к перенесению неблагоприятных условий, может быть и более сложным; таковы, например, полиэдры, образуемые некоторыми В. насекомых (они состоят из оболочки, кристаллической белковой массы и включенных в неё частиц В.).
Химический состав разных В. неодинаков. Одни В. содержат липиды; среди них есть В. с ДНК (оспы, герпеса и др.), с РНК (гриппа, птичьей чумы, саркомы Рауса, бронзовости помидора, жёлтой карликовости картофеля и др.). У других. В. липиды отсутствуют. В этой группе также есть В. с ДНК (аденовирусы, большинство бактериофагов, В. желтухи шелкопряда) и с РНК (полиомиелита, ящура; большинство В., вызывающих болезни растений; некоторые бактериофаги). Кроме липидов, белка и нуклеиновой кислоты, в В. встречаются в небольшом количестве полиамины (путресцин, спермидин и др.), иногда витамины (витамин B2, фолиевая кислота), а также ряд металлов; в некоторых В. содержатся соединения белка с полисахаридами.
Размножение В. происходит в клетках. Бактериофаги растворяют оболочку бактерии и вводят в бактерию нить НК, причём капсид фага остаётся вне клетки. Многие В. поглощаются клеткой путём Пиноцитоза. Попав в клетку, они освобождаются от оболочки. Первые этапы развития В. в клетке в общих чертах состоят в том, что строятся так называемые ранние белки, т. е. белки-ферменты, необходимые В. для репликации (удвоения) их НК. Так называемые поздние белки участвуют в образовании белковых оболочек дочерних вироспор. Из ферментов у В., содержащих ДНК, одним из первых синтезируется полимераза РНК, которая строит на нити ДНК информационную РНК (и-РНК). Эта РНК попадает на Рибосомы клетки, где и происходит синтез других белков вирусной частицы (см. Белки, раздел Биосинтез). В., содержащие РНК, синтезируют полимеразу, катализирующую синтез новых частиц вирусной РНК; эта РНК переходит на рибосомы и контролирует синтез белка капсида. Таким образом, В., содержащие РНК, не нуждаются в ДНК для размножения и передачи генетической информации потомству (см. схему).
От этой общей схемы размножения В. имеются различные отклонения. Так, некоторые В. содержат белки-ферменты; В. осповакцины синтезирует в клетке хозяина двойные нити РНК и т.д. Многие особенности размножения В. ещё не выяснены. Существуют, например, особые очаги размножения нитей НК, и при созревании частиц В. синтезируется белок, охватывающий отдельные отрезки НК. Иногда этот процесс идет несовершенно, образуются неполноценные частицы В., в которых нет или мало содержимого, это — так называемые неинфекционные В. Во многих случаях очаги размножения В. хорошо видны в клетке под микроскопом. Эти очаги называются внутриклеточными включениями, или Х-телами. Когда Х-тело заканчивает свое развитие, в нём образуется вироспора. У многих В. вироспоры образуют в Х-телах кристаллические агрегаты, у других В. они неизвестны. Некоторые В. размножаются в ядре клетки другие — в ее цитоплазме, третьи — и в ядре, и в цитоплазме. НК находится в вироспоре в специально закрученном состоянии. Длина нити НК у разных В. различна. Так, у В. оспы она достигает 83 мкм, у крупных бактериофагов, например Т4, — 70 мкм. У мельчайших бактериофагов нить НК имеет длину около 2 мкм. В зависимости от длины нити НК (что определяет объем наследственной информации, которой располагает тот или иной В.), т. е. от способности В. синтезировать более или менее разнообразные молекулы белков, различна степень участия составных частей клетки-хозяина в размножении В. и их построении. В., имеющие нить НК значительной длины, могут синтезировать многие вещества. Так, некоторые бактериофаги синтезируют в клетке несколько десятков разных белков. Все В., содержащие ДНК, синтезируют собственную РНК. Даже если клетка-хозяин имеет необходимые для В. ферменты, В. очень часто синтезируют собственные ферменты, обладающие подобным действием. Мельчайшие фаги обладают информацией для синтеза только трёх собственных белков; например, фаг МЗ-2 синтезирует зависящую от РНК полимеразу и два белка, необходимые для построения зрелых частиц В. Таким образом, степень зависимости В. от различных ферментов клетки-хозяина различна. Некоторые В. так бедны наследственной (генетической) информацией, что могут размножаться в клетке только в присутствии др. В. Зависимость В. не только от клетки, но и от других В. существует, например, между В. некроза табака и его спутником, вироспоры которого мельче вироспор некроза табака. Ещё более тесные взаимоотношения существуют между некоторыми В., поражающими животных и человека. Среди В., способных вызывать злокачественные опухоли (см. Опухолеродные вирусы), известны В. с дефектной частицей, которая не может образовывать собственную белковую оболочку. Эти В. достигают зрелого состояния, только если они размножаются в присутствии других В. (таковы отношения, например, между опухолеродным обезьяньим вирусом S-40 и некоторыми аденовирусами (См. Аденовирусы)). НК опухолеродного В. в этом случае включается в капсид аденовируса и вместе с ним попадает в чувствительную клетку. Выход В. из клетки в одних случаях совершается только при разрушении клетки (многие фаги, В. оспы), в других — частицы В. покидают клетку, не убивая её при этом (Миксовирусы, некоторые мелкие фаги).
Если в клетку попадают В., различающиеся по тем же или другим Генам (различие может быть результатом Мутации), то в потомстве можно наблюдать В., соединяющие свойства двух и больше исходных форм. Это указывает на наличие обмена (перекомбинации) признаков таких форм при размножении В. в одной клетке. Закономерности этих процессов изучает генетика В. (см. Генетика микроорганизмов).
Устойчивость вироспор к внешним воздействиям различна, но по большей части велика. Некоторые В. инактивируются только при нагревании до 90°С (В. мозаичной болезни табака), легко переносят очень низкие температуры (—70°С и ниже), а также высушивание.
Способы распространения В. в природе различны: многие из них могут непосредственно заражать чувствительный организм (В. гриппа, оспы, мозаичной болезни табака, бактериофаги), иные циркулируют в природе более сложным образом и переносятся при помощи других организмов. Так, В. некроза табака передаётся при помощи обитающего в почве грибка (Olpidium): последний, проникая в корни растения, вносит и В. Многие В. передаются паразитирующими у растений нематодами (См. Нематоды). В. животных, человека и растений переносят также клещи и насекомые. Передача одних В. сосущими членистоногими носит механический характер; в других случаях В. проделывают часть своего развития в переносчике и даже могут передаваться с яйцами переносчика из поколения в поколение. Многие В., поражающие человека и домашних животных, обитают и в диких животных; поражающие культурные растения — в диких растениях и сорняках.
Попытки обнаружить жизнедеятельность вироспор вне клетки, естественно, не увенчались успехом: известно, что покоящиеся формы жизни вообще не обнаруживают жизнедеятельности (см. Анабиоз). В бесклеточных системах можно воспроизвести отдельные этапы размножения В., получить саморепродукцию вирусной НК, а также под контролем этой НК — синтез белков, характерных для В. Но эти процессы идут только в присутствии извлечённых из клетки рибосом; следовательно, эти системы, хотя и являются бесклеточными, не могут рассматриваться как вполне искусственно синтезируемые.
О происхождении В. имеются различные предположения. Некоторые считают, что В. могут спонтанно зарождаться в организме хозяина под влиянием неблагоприятных условий. Но это мнение опровергается следами длительной эволюции В. (их приспособление к циркуляции в природе), а также отсутствием переходных форм между В. и органоидами клетки. Другие исследователи думают, что В. — потомки простейших форм жизни, однако и это предположение маловероятно, так как выраженный паразитический характер В. предполагает существование более высокоорганизованных существ, в которых В. могли бы жить и размножаться. Поэтому наиболее вероятно, что В. возникли от свободно живущих более сложно организованных форм, и простота В. вторична, она — результат приспособления к паразитическому образу жизни. Такая вторичная простота, связанная с утратой приспособленности к самостоятельному питанию и усилением способности к размножению, вообще очень характерна для паразитов. В пользу древности В. и длительной их эволюции говорит также то, что они вступают в сложные взаимоотношения с другими видами животных и растений (трансмиссивные В., передаваемые различными животными).

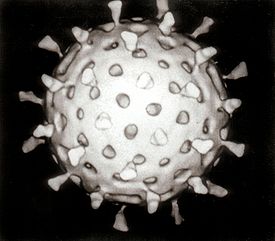
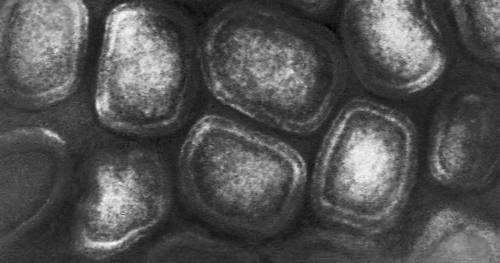
Различные типы вирионов под электронным микроскопом. Бактериофаг с длинным отростком. Увеличено в 240 000 раз.
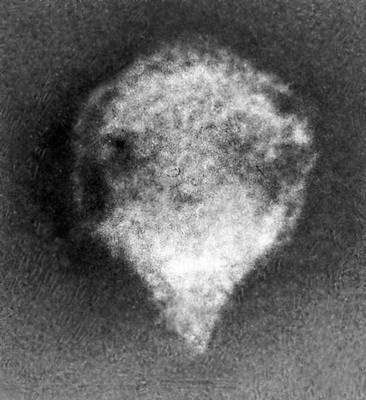
Различные типы вирионов под электронным микроскопом. Бактериофаг с коротким отростком. Увеличено в 600 000 раз.

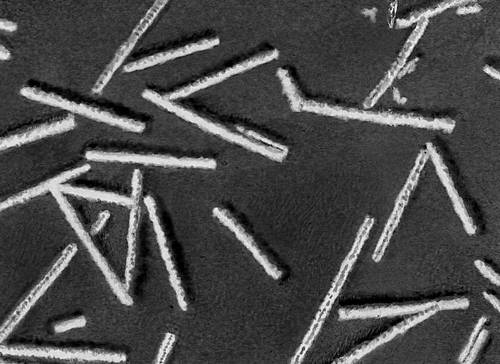
Модели вирусных частиц. Часть палочки вируса мозаичной болезни табака; видны капсомеры и инкрустированные в них витки нуклеиновой кислоты.

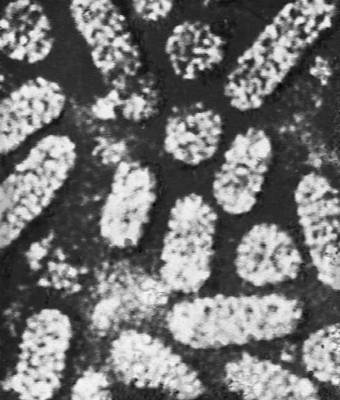
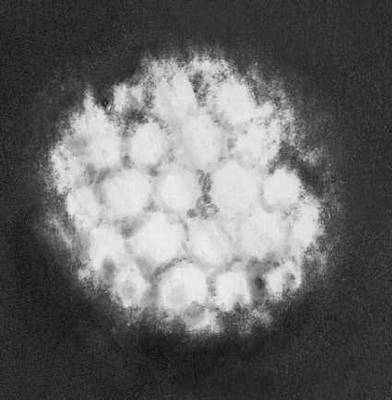
Различные типы вирионов под электронным микроскопом. Вирус мозаичной болезни табака. Увеличено в 50 000 раз.
Различные типы вирионов под электронным микроскопом. Вирус кольцевой пятнистости малины. Увеличено в 135 000 раз.

Схема размножения вирусов, содержащих в вирионе одну нить ДНК (I) или одну нить РНК (II). ДНК изображена сплошной линией, РНК — пунктиром; А — нуклеиновая к-та вириона; Б — удвоенная нить нуклеиновой кислоты при ее репликации; В — информационная РНК, (и-РНК), копирующая вирусную ДНК; Г — цепочка рибосом (полисома), соединенная и-РНК или вирусной РНК (на рибосомах растет полипептидная цепочка из остатков аминокислот); Д — рибосома с полипептидом, отделившаяся от полисомы; Е — белковая молекула, образованная полипептидными цепочками; Ж — построение дочерней нити нуклеиновой к-ты между двумя материнскими; З — зрелый вирион. Стадия В у вирусов с РНК отсутствует, т. к. их собственная РНК выполняет при синтезе белков роль и-РНК.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Полезное
Смотреть что такое "Вирусы" в других словарях:
ВИРУСЫ — (от лат. virus яд), неклеточные формы жизни, способные проникать в определённые живые клетки и размножаться только внутри этих клеток. Подобно всем др. организмам В. обладают собств. генетич. аппаратом, к рый кодирует синтез вирусных частиц из… … Биологический энциклопедический словарь
ВИРУСЫ — (от лат. virus яд) мельчайшие неклеточные частицы, состоящие из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки (капсида). Форма палочковидная, сферическая и др. Размер 15 350 нм и более. Открыты (вирусы табачной мозаики) Д. И. Ивановским в … Большой Энциклопедический словарь
ВИРУСЫ — (от латинского virus яд), неклеточные формы жизни. Состоят из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки (капсида). Открыты в 1892 российским ученым Д.И. Ивановским. Вирусы внутриклеточные паразиты: они размножаются только в живых… … Современная энциклопедия
вирусы — неклеточные формы жизни, способные проникать в определенные живые клетки и размножаться только внутри этих клеток. В. обладают собственным генетическим аппаратом, который кодирует синтез вирусных частиц из биохим. предшественников, находящихся в… … Словарь микробиологии
Вирусы — Вирусы: Группа ультрамикроскопических внутриклеточных паразитов, состоящих из нуклеиновой кислоты, окруженной защитной протеиновой или смешанной оболочкой из протеинов, липидов и углеводов. Источник: ВОДА И ВОДОПОДГОТОВКА. ТЕРМИНЫ И ОПРЕДЕЛЕНИЯ … Официальная терминология
ВИРУСЫ — мельчайшие возбудители инфекционных болезней. В переводе с латинского virus означает яд, ядовитое начало . До конца 19 в. термин вирус использовался в медицине для обозначения любого инфекционного агента, вызывающего заболевание. Современное… … Энциклопедия Кольера
вирусы — (от лат. virus яд), мельчайшие неклеточные частицы, состоящие из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки (капсида). Форма палочковидная, сферическая и др. Размер 15 350 нм и более. Открыты (вирусы табачной мозаики) Д. И. Ивановским … Энциклопедический словарь
ВИРУСЫ — Схематическое изображение сферического вируса и вируса со спиральной симметрией. Схематическое изображение сферического вируса (а) и вируса со спиральной симметрией (б): 1 структурная единица, или субъединица; 2 морфологическая… … Ветеринарный энциклопедический словарь
Читайте также: