
Что такое адсорбция вируса
Обновлено: 19.04.2024
ГЕМАДСОРБЦИЯ (греч. haima кровь + адсорбция) — способность клеток, зараженных нек-рыми вирусами (гл. обр. гемагглютинирующими), фиксировать на своей поверхности эритроциты.
Разработанные на основе этого явления реакция Г. и реакция торможения Г. служат для обнаружения, титрования и идентификации ряда вирусов, а также для выявления соответствующих антител в сыворотках.
Г. с целью обнаружения вирусов в исследуемом материале от больных чаще всего используют при диагностике оспы и парагриппозных заболеваний.
Г. впервые описали Фогель и Щелоков (J. Е. Vogel, A. Shelokov) в 1957 г. Они обнаружили адсорбцию эритроцитов морской свинки однослойными культурами почечных клеток обезьян, зараженными вирусом гриппа.
Достоинство реакций Г. и торможения Г. заключается в более раннем появлении у зараженных клеточных культур способности к Г. по сравнению с развитием выраженного цитопатического эффекта, а также в возможности выявления некоторых вирусов, размножение которых в культурах вызывает слабо выраженные и нерегулярные цитопатические изменения (напр., парагриппозных вирусов).
Способность к Г. обнаружена у различных вирусов, принадлежащих к разным классификационным группам (табл.). Большинство из них способно вызывать гемагглютинацию (см.), т. е. соединяться с эритроцитами и вызывать их склеивание. Способность этих вирусов к Г. также обусловлена их гемагглютинирующей активностью и, вероятно, вызывается вирусными частицами, еще не полностью освободившимися из клетки и расположенными у ее поверхности. Механизм соединения клеток с эритроцитами такой же, как при гемагглютинации, т. е. вирионы вступают в контакт с рецепторами эритроцитов.
ВИРУСЫ, ВЫЗЫВАЮЩИЕ ГЕМАДСОРБЦИЮ, И УСЛОВИЯ ЕЕ ВЫЯВЛЕНИЯ
Классификационная группа (род вирусов)
Выявление гемадсорбирующей активности вирусов
на культурах клеток
Африканской чумы свиней
Лейкоциты и костный мозг свиней
Инфекционного бронхита птиц
Почки эмбрионов кур
Почки обезьян и плода человека
Морских свинок, кур, человека
Почки поросят, телят, обезьян, хомячков, мышей, плода человека
Морских свинок, кур, человека
Почки обезьян, кожно-мышечная ткань эмбриона человека, амнион человека
Почки обезьян и плода человека
Парагриппа-2 обезьян (SV5, SV41)
Парагриппа-3 крупного рогатого скота
Почки телят, кроликов, собак, обезьян, цыплят
Морских свинок, кроликов, овец, крупного рогатого скота
Почки обезьян, морских свинок, кожно-мышечная ткань эмбрионов кур и японских перепелов
Кожно-мышечная ткань эмбрионов кур и японских перепелов
Кожно-мышечная ткань эмбрионов кур
Парвовирус-1 крупного рогатого скота
Почки крупного рогатого скота
Морских свинок, собак, человека
Исключение составляют возбудители инфекционного бронхита птиц и африканской чумы свиней, поскольку выделившиеся из клеток вирионы не обладают гемагглютинирующими свойствами. У вируса бронхита птиц гемагглютинин можно обнаружить путем дезинтеграции внешней оболочки трипсином. По-видимому, способность к Г. у зараженных этим вирусом клеток связана e одной из стадий синтеза вириона. Механизм Г., к-рая наблюдается в культурах, зараженных вирусом африканской чумы свиней, также не ясен. Однако не вызывает сомнения, что это явление связано с репродукцией вируса, поскольку эритроциты адсорбируются только на пораженных вирусом клетках, где развиваются специфические изменения и формируются включения.
Для выявления Г. в культурах, зараженных гемагглютинирующими вирусами, используют тот вид эритроцитов, который агглютинируется данным вирусом.. Однако спектр чувствительных эритроцитов, способных адсорбироваться на зараженных клетках, шире, чем чувствительных к агглютинирующему действию тех же вирусов. Подобное явление было выявлено, напр., в опытах с вирусами краснухи и осповакцины.
В пробирки с клеточным монослоем, зараженным вирусом, после удаления или без удаления питательной среды вносят 0,4—1% взвесь эритроцитов в объеме 0,2—0,5 мл. Пробиркам придают наклонное положение, чтобы взвесь эритроцитов покрывала клеточный слой, и выдерживают в течение определенного времени при t° 4° или 20—25°. Затем неадсорбированные эритроциты отделяют от поверхности клеток отмыванием культуры р-ром Ханкса или путем непродолжительного покачивания (или вращения) пробирок. Наличие Г. определяют при малом увеличении микроскопа.
Г. бывает островной и диффузной. Островной тип Г. наблюдается в культурах, зараженных, напр., вирусами гриппа, паротита, оспы. При этом эритроциты фиксируются на ограниченных участках культуры, соответствующих одной или нескольким клеткам. Парагриппозные вирусы вызывают диффузную адсорбцию эритроцитов на всем клеточном пласте.
Для обнаружения парагриппозных вирусов человека с целью диагностики заражают культуры почечных клеток обезьян или 12—30-недельного плода человека. Цитопатический эффект при этом появляется поздно или может вообще отсутствовать. Обычно реакцию Г. проводят на 4—5-й день культивирования с эритроцитами морской свинки.
При отсутствии Г. культивирование клеток продолжают до 20 дней и реакцию повторяют каждые 2—5 дней с полной или частичной сменой среды.
При диагностике оспы к Г. прибегают, если в зараженных исследуемым материалом культурах кожно-мышечных клеток эмбриона человека или почечных клеток обезьян цитопатические изменения слабо выражены (обычно они появляются через 24—96 час.).
Для выявления вируса клещевого энцефалита метод Г. не нашел широкого применения ввиду частых неспецифических реакций.
Г. используют при титровании на культурах клеток парагриппозных вирусов, а также возбудителей кори и паротита. При этом эритроциты вносят однократно в конце опыта.
Минимальная доза вируса, вызывающая отчетливую Г., носит название гемадсорбирующей единицы.
Вирусы, вызывающие островковую Г., можно титровать путем подсчета числа фокусов Г., т. е. участков, состоящих из одной или нескольких продуцирующих вирус клеток. Этот метод применяется редко.
Реакция торможения гемадсорбции служит для контроля специфичности наблюдаемой Г., идентификации выделенных вирусов и титрования иммунных сывороток.
Для идентификации и титрования вирусов их смешивают с соответствующей иммунной сывороткой в равном объеме и вносят в пробирки с клеточной культурой. Через 30 мин. инкубации жидкость отсасывают и наливают питательную среду. Эритроциты вносят после окончания срока инкубации (при работе с цитопатогенными вирусами — после появления соответствующих изменений).
С той же целью можно использовать культуры клеток, предварительно инфицированные испытуемым вирусом. Через определенный срок после заражения (в опытах с парагриппозными вирусами обычно на 5-й день) питательную среду удаляют и в культуры вносят иммунную сыворотку. После 15—30-минутной инкубации в комнате в пробирки наливают взвесь эритроцитов. Учет реакции производят через 3—5 мин.
С целью титрования антител реакцию задержки Г. используют гл. обр. в опытах с парагриппозными вирусами и возбудителем паротита. Для определения количества парагриппозных антител к двукратным разведениям испытуемой сыворотки прибавляют вирус в количестве 100 гемадсорбирующих единиц. Смеси выдерживают 2 часа в комнате, после чего помещают в пробирки с клеточной культурой. Эритроциты прибавляют через 4—5 дней.
При титровании антител к вирусу паротита смеси вируса с сывороткой вносят в культуры кожно-мышечных клеток куриных эмбрионов. Через 5—6 дней среду удаляют и в пробирки помещают 1% взвесь эритроцитов курицы в объеме 0,5 мл.. Этот метод определения количества паротитных антител дает более четкие результаты, чем учет реакции по цитопатическому действию вируса или по определению наличия гемагглютинина в культуральной жидкости.
При работе с гемадсорбирующими вирусами и особенно при попытках обнаружения вирусов в изучаемом материале следует иметь в виду возможность неспецифической фиксации эритроцитов на клетках, а также контаминацию (загрязнение) культур и питательных сред посторонними вирусами, вызывающими Г. Неспецифическая спонтанная Г. нередко имеет место, напр., при использовании эритроцитов старых морских свинок. Во избежание ошибок специфичность наблюдаемой Г. должна постоянно контролироваться в опытах нейтрализации с соответствующими иммунными сыворотками.
Библиография: Лабораторная диагностика вирусных и риккетсиозных заболеваний, под ред. Э. Леннета и Н. Шмидт, пер. с англ., М., 1974; Справочник по микробиологическим и вирусологическим методам исследования, под ред. М. О. Биргера, М., 1973; S t а г k e G. u. H 1 in a k P. Grundriss der allgemeinen Viro-logie, Jena, 1974, Bibliogr.
Типы инфицирования клеток вирусами. Репродуктивный цикл вирусов. Основные этапы репродукции вирусов. Адсорбция вириона к клетке.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Репродуктивный цикл вирусов
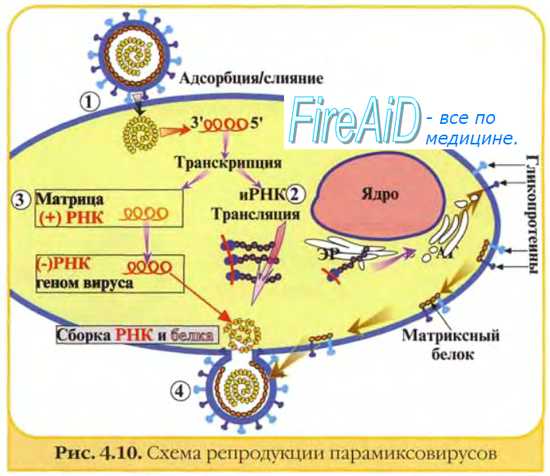
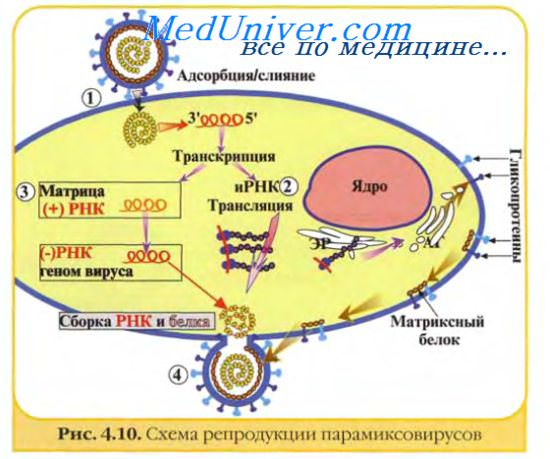
Изображённые на рис. 2-3 этапы репродукции (от адсорбции вирионов до высвобождения дочерней популяции) происходят при продуктивном взаимодействии вируса с клеткой.
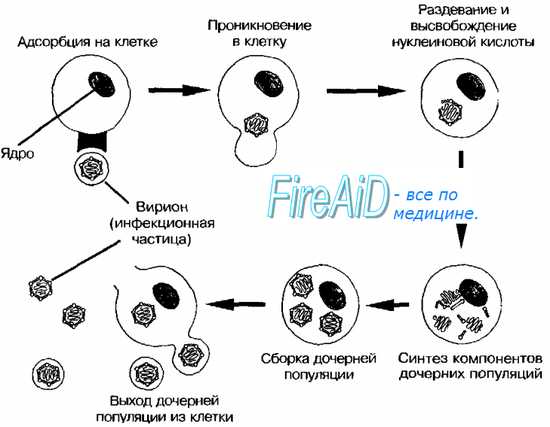
Рис. 2-3. Основные этапы репродукции вирусов.
Адсорбция вириона к клетке
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Цикл размножения вирусов. Прикрепление или адсорбция вируса.
Цикл репродукции вирусов при продуктивной инфекции представляет собой специфическую последовательность событий, приводящих к образованию нового потомства вирионов. Его можно разделить на несколько стадий: адсорбция вирионов на поверхности клетки, проникновение вируса или вирусного нуклеопротеида внутрь клетки, депротеинизация генома, синтез вирусных компонентов, формирование и выход из клетки зрелых вирионов. Иногда не все стадии протекают последовательно и полностью даже при продуктивной инфекции. При заражении вирусной нуклеиновой кислотой отсутствует стадия депротеинизации. Цикл репродукции вируса может прерываться на какой-либо промежуточной стадии, и в этом случае инфекционные вирионы не образуются (абортивная инфекция).
Классическое изучение кривой роста вируса в одном цикле репродукции предполагает одновременное инфицирование клеток культуры с использованием высокой множественности заражения (М=10—100) и определение накопления вируса через определенные промежутки времени.
Прикрепление (адсорбция) вирионов к поверхности клетки - первая стадия вирусной инфекции. Для того, чтобы началась инфекция, вирионы должны быть способны прикрепляться к клетке. Безусловно, этот процесс в организме является более сложным, чем в культуре клеток.
Детальное изучение процесса адсорбции вирусов показало, что он состоит из двух быстро следующих друг за другом периодов — обратимого и необратимого. В период обратимой адсорбции вирус можно удалить с поверхности клетки при обработке версеном, хемотрипсином и другими химическими веществами. При необратимой адсорбции вирус удалить с поверхности клетки не удается.
Первичный контакт с клеткой происходит в результате случайных столкновений вирионов с поверхностью клетки. Количество стабильных прикреплений вириона к клетке во много раз меньше количества случайных столкновений.
В основе прикрепления вируса к клетке лежат два механизма — неспецифический и специфический. Первый из них определяется силами электростатического взаимодействия, возникающими между разноименно заряженными группами, расположенными на поверхности клетки и вируса. В этом процессе, прежде всего, могут участвовать положительно заряженные аминные группы вирусного белка и отрицательно заряженные группы клеточной поверхности. В пользу электростатического взаимодействия между клеткой и вирусом свидетельствуют данные о зависимости присоединения вирионов к клеткам от ионной силы раствора и его рН, наличия в среде одно- и двухвалентных катионов и присутствия отрицательно заряженных групп на поверхности клетки.
Наиболее важным механизмом прикрепления вируса является специфическое взаимодействие рецепторов вируса с комплементарными рецепторами клетки.
Прикрепление происходит за счет связи между вирионными прикрепительными белками на поверхности вирионов и рецепторами на плазматической мембране клеток.
На поверхности вирионов многих семейств вирусов (все РНК-содержащие оболочечные вирусы и аденовирусы) имеются пепломеры (выступы и шипы) длиной от 10 до 30 нм, которые принимают непосредственное участие в прикреплении вируса к клетке. Отщепление пепломеров с помощью протеолитических ферментов приводит к потере вирионами способности адсорбироваться на клетках. На поверхности вирионов, не имеющих пепломеров, находятся участки связывания с клеточной поверхностью, которые отличаются сложным строением и, вероятно, состоят из нескольких полипептидов.
Спектр чувствительности клеток к вирусам в значительной мере определяется наличием соответствующих рецепторов. Рецепторный барьер может быть преодолен при заражении инфекционной нуклеиновой кислотой.
На поверхности клетки может находиться большое количество различных видов рецепторов. Полагают, что каждая клетка может адсорбировать от десятков до нескольких тысяч вирионов. На поверхности одной клетки находится около 10000 рецепторов для полиовируса. Каждый рецептор специфичен для одного или нескольких вирусов, как родственных между собой, так и относящихся к различным таксономическим группам. Например, аденовирус типа 2 и вирус Коксаки ВЗ связываются с одними и теми же клеточными рецепторами.
У герпесвирусов прикрепительными белками могут служить гликопротеины оболочки, которые соединяются с некоторыми рецепторами клетки. Клеточным рецептором для многих ортомиксовирусов являются концевые сиаловые кислоты олигосахаридов, входящих в структуру гликопротеринов или гликолипидов, экспонированных на поверхности клетки. Рецепторами для многих риновирусов являются иммуноглобулины. У вируса ВИЧ в прикреплении первоначально участвуют молекулы СД4, расположенные на поверхности клеток, особенно макрофагов и Т-хелперных лимфоцитов. Подобные взаимодействия рецептор-лиганд свойственны другим лентивирусным инфекциям.
Рецепторы для одних вирусов имеются на клетках нескольких или многих тканей, тогда как для других вирусов они обнаружены на клетках одного вида тканей или в один из периодов онтогенеза (новорожденные или взрослые животные).
Рецепторы клеток способны к регенерации после удаления их протеолитическими ферментами.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Клетки животных имеют свойство притягивать (адсорбировать) ряд вирусов, вероятно, вследствие взаимодействия ионизированных групп. Процесс проникновения точно не установлен, но, учитывая, что клетки способны захватывать мелкие капли жидкости, можно предположить, что вирусные частицы заглатываются таким же образом и попадают в пузырьки и в цитоплазму клетки. Замечено, что определенный вирус способен адсорбироваться только на ограниченном типе клеток. Например, вирус полиомиелита адсорбируется только на некоторых клетках приматов. Существуют фаги, которые адсорбируются только на определенных мутантах микроорганизмов, только на мужских (т. е. несущих половой фактор) или только на женских клетках и т. п. Таким образом, адсорбция вируса на клетке является высоко-специфическим процессом.
Каждая клетка может адсорбировать большое, но тем не менее ограниченное число вирусных частиц. Так, например, одна клетка бактерии связывает около 300 частиц бактериофага. Фактические подсчеты показали, что при этом вся поверхность клетки покрывается фагом.
Прикрепление вириона к клеточной поверхности происходит путем случайных столкновений. Но не при каждом столкновении вирион прикрепляется к клетке.
Рецепторы связывающие вирус
Клеточная поверхность также обладает особыми клеточными рецепторами — структурами, которые ответственны за связывание вируса. В некоторых случаях эти клеточные рецепторы сосредоточены в определенных морфологических образованиях. Например, некоторые фаги прикрепляются только к жгутикам клетки хозяина. Чаще вирусы прикрепляются к участкам клеточной поверхности, которые не имеют выраженных морфологических отличий от соседних участков. Вирусная частица может эффективно реагировать только с определенными, специфичными для данного вируса рецепторами клетки.
Таким образом, первой стадией взаимодействия вируса и клетки является реакция между вирусными и клеточными рецепторами. Наличие соответствующих рецепторов — один из важных факторов, определяющих возможность или невозможность для данного вируса вызвать инфекционный процесс в клетке.

Репродукция вируса – это процесс размножения вирусных частиц в чувствительных к ним клетках. Репродуцируются только вирулентные вирусы, обладающие высокой степенью патогенности [3] .
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций [3] .
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина [2] .
- Все вирусы, содержащие молекулу ДНК, кроме вирусаоспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирусоспы размножается только в цитоплазме клетки [2] .
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами [2] .
- Белки вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению) [2] .
- Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование (присоединении к полипептиду углеводных остатков) [2] .

Этапы репродукции (жизненного цикла) вируса

1. Адсорбция вируса на мембране клетки.; 2. Проникновение вируса в клетку.; 3. Депротеинизация.; 4. Синтез компонентов вирусов.; 5. Формирование дочерних вирионов.; 6. Выход вирионов [2] .
Этапы репродукции вируса
Репродукцию или жизненный цикл вируса делят на шесть последовательных этапов:
- Адсорбция на мембране клетки [2] .
- Проникновение в клетку [2] .
- Депротеинизация [2] .
- Синтез компонентов вирусов[2] .
- Формирование дочерних вирионов[2] .
- Выход вирионов[2] .

Процесс адсорбции вириона на поверхности клетки

1. Вирион.; 2. Клеточный рецептор.; 3. Прикрепительный белок; 4. Клеточная мембрана [2] .
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона [2] .
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур [2] .
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно [2] .
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 [2] .

Проникновение вируса в клетку (Путь I)

Слияние вирусной оболочки с клеточной мембраной
1. Вирион.; 2. Инфицируемая клетка.; 3. Ядро [2] .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона [2] . Существует два пути:
- Путь I – слиянии вирусной оболочки с клеточной мембраной [2] .
- Путь II – рецептор-опосредованный эндоцитоз [2] .
Путем слияния суперкапсида с клеточной мембраной (путь I) в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки [2] .
Путем рецептор-опосредованного эндоцитоза (путь II) в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении, вирусный геном в цитоплазме клетки освобождается из эндосомы [2] .

Проникновение вируса в клетку (Путь II)

1. Вирион.; 2. Клетка.; 3. Эндосома.; 4. Ядро [2] .
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы) [2] .
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона [2] .
Синтез компонентов вируса
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [1] .
Место синтеза компонентов дочерних вирионов зависит от типа генома:
- реализация генетической информации у ДНК-содержащих вирусов идет по пути: ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом идет по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок;
- РНК-содержащие ретровирусы идут по следующему пути передачи информации: РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок [2] .
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона [2] .
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках [2] .
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК [2] .
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается [2] .
РНК-содержащие ретровирусы отличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза [2] [4] .
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков, и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры [2] .
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более [2] .
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования[2] .
Путь лизиса клетки тесно связан ее деструкцией. Он характерен для безоболочечных вирусов, не имеющих суперкапсидной оболочки (суперкапсида) [2] .
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы, высвобождаются в течении 2–6 часов. В начале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида [2] .
Читайте также: