Что такое репликация вирусной нуклеиновой кислоты
Обновлено: 24.04.2024
Вирусная репликация представляет собой набор процесса биохимического происходящие в клетке , инфицированной с помощью вируса и которые оказывают влияние на производстве новых единиц этого вируса (или вирионов ). Этот способ размножения паразитов, использующий механизм репликации ДНК, и определяет вирусы.
Вирусы должны сначала проникнуть в клетку, прежде чем может произойти вирусная репликация. Благодаря генерации большого количества копий своего генома и упаковке этих копий вирус может продолжать заражать новых хозяев. Репликация внутри вирусов очень разнообразна и зависит от типа вовлеченных в них генов . Большинство ДНК-вирусов собирается в ядре , в то время как большинство РНК-вирусов растут только в цитоплазме .
Резюме
Классический цикл репликации вируса
Каждый вирус имеет очень специфические способы репликации, в частности, в зависимости от того, является ли он ДНК-вирусом или РНК-вирусом . Вирусы размножаются только в живых клетках. Клетка-хозяин должна обеспечивать энергию и синтетические механизмы, а также низкомолекулярные предшественники для синтеза вирусных белков и нуклеиновых кислот . Репликация вируса представлена здесь только в общих чертах и состоит из семи этапов:
- Адсорбция;
- Проникновение в хозяйскую клетку;
- Декапсидирование;
- Репликация генома;
- Синтез вирусных компонентов;
- Сборка и упаковка;
- Релиз.
Адсорбция
На этом этапе и в двух упомянутых выше случаях происходит связывание белка вирусной оболочки с рецептором клеточной мембраны. Рецепторами эукариотических клеток могут быть гликопротеины или гликосфинголипиды . Рецепторы бактериофагов представляют собой гликопротеины или липополисахариды . Клетки растений не имеют специфических рецепторов к вирусам.
Проникновение
В зависимости от вируса существует несколько механизмов проникновения внутрь клетки. У бактериофагов в бактериальную клетку попадает только вирусный геном. В вирусы животных вирус может проникать по нескольким механизмам. Вирус может проникать путем пиноцитоза: вирусный нуклеокапсид , окруженный плазматической мембраной, проникает в клетку через пиноцитотическую вакуоль. Это защищает клетку от антител , как в случае с ВИЧ . Это часто бывает с незащищенными вирусами. В случае вирусов в оболочке вирус может проникать либо путем слияния (между вирусной оболочкой и плазматической мембраной клетки), либо путем эндоцитоза (накопление вирусных частиц в цитоплазматических везикулах).
Декапсидация
Клеточные ферменты (из лизосом ) удаляют белковую оболочку вируса. Это высвобождает или делает доступной нуклеиновую кислоту или геном вируса. В зависимости от вируса декапсидация может происходить в цитоплазме или ядре .
Репликация генома
В зависимости от типов вирусов и природы их генома этот шаг может варьироваться. Во время этого цикла ( транскрипция, затем перевод ) невозможно выделить вирусную частицу.
В некоторых РНК-вирусах заражающая РНК продуцирует информационную РНК (мРНК). Затем он будет использован для перевода генома в белковые продукты (см. Следующий шаг). Для других вирусов, таких как вирусы с РНК или ДНК с отрицательными цепями, вирусы образуются путем интеграции генома в геном клетки-хозяина (ретротранскрипция РНК в ДНК, затем вставка в ДНК клетки-хозяина в случае вирусов с отрицательной цепью РНК или прямая вставка вирусной ДНК в ДНК клетки-хозяина в случае вирусов с ДНК с отрицательной цепью) с последующей транскрипцией ДНК клетки-хозяина с помощью обычного механизма репликации. Полученные мРНК будут содержать мРНК, полученные из вируса, что позволит продуцировать вирусные белки.
Синтез вирусных компонентов (белков и нуклеиновой кислоты)
Следующие компоненты производятся вирусом через органеллы, существующие в клетке-хозяине.
- Синтез вирусных белков: мРНК вируса транслируется на уровне рибосом клеток в два типа вирусных белков:
- Структурный: белки, составляющие вирусную частицу, производятся и собираются.
- Неструктурные: они не обнаруживаются в вирусной частице; в основном это ферменты для репликации вирусного генома ( например, полимераза )
Сборка и упаковка
Вирионы собираются и созревают в инфицированных клетках. Геном инкапсидируется вирусными белками с образованием новых вирусных частиц.
Это может происходить в ядре клетки, цитоплазме или плазматической мембране для большинства вирусов в оболочке. Оболочечные вирусы приобретают свои оболочки путем почкования в ущерб плазматической мембране или ядерной мембране клетки-хозяина.
Выпуск вирионов
Восстановленные вирионы выходят за пределы клетки. Это происходит либо в результате внезапного разрушения клеток, либо в результате постепенного вытеснения (образования почки) оболочечных вирусов через клеточную мембрану.
Новые вирусы могут вторгаться или атаковать другие клетки или оставаться в спящем состоянии в исходной клетке-хозяине. В случае бактериофагов высвобождение вновь образованных вирионов происходит путем лизиса инфицированных бактерий . Однако в случае вирусов животных высвобождение обычно происходит без лизиса клеток.
Частный случай поксвирусов
Репликация Poxviridae необычна для вирусов с двухцепочечной ДНК (дцДНК), потому что она происходит в цитоплазме .
Частный случай гигантских вирусов
В мимивирусе имеет особенность репликации с помощью фагоцитоза .
Балтиморская система классификации
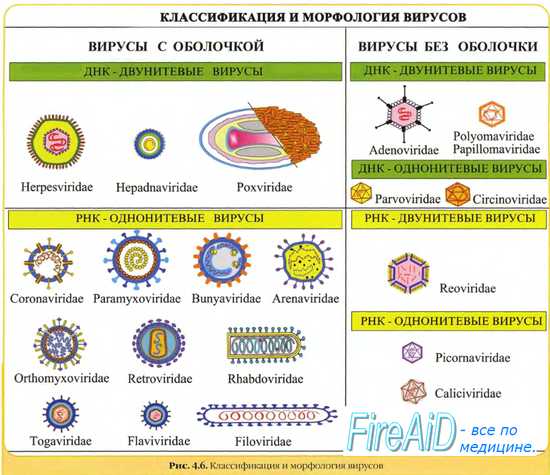
Вирусы подразделяются на 7 типов генов, каждый из которых имеет свои собственные семейства вирусов, которые, в свою очередь, имеют разные стратегии репликации. Лауреат Нобелевской премии биолог Дэвид Балтимор разработал систему классификации для сортировки различных вирусов на основе их уникальной стратегии репликации. На основе этой системы существует семь различных стратегий репликации (Класс I, II, III, IV, V, VI, VII). Здесь кратко и в общих чертах перечислены семь классов вирусов.
Класс I: двухцепочечный ДНК-вирус
Этот тип вируса обычно должен проникнуть в ядро хозяина, прежде чем он сможет реплицироваться. Некоторым из этих вирусов требуются полимеразы клеток-хозяев для репликации их генома, тогда как другие, такие как аденовирусы или герпесвирусы, кодируют свои собственные факторы репликации. Однако в обоих случаях репликация вирусного генома сильно зависит от состояния клетки, допускающего репликацию ДНК, и, следовательно, от клеточного цикла. Вирус может вызвать принудительное деление клетки, что может привести к трансформации клетки и, в конечном итоге, к раку . Примером семейства этой классификации являются аденовирусы.
Есть только один хорошо изученный пример, в котором семейство вирусов класса I не реплицируется в ядре: это семейство поксвирусов, которое включает высокопатогенные вирусы, поражающие позвоночных (см. Выше).
Класс II: вирус с одноцепочечной ДНК
В эту категорию попадают вирусы, которые не так хорошо изучены, как предыдущие, но все же оказывают сильное влияние на позвоночных. Двумя примерами являются Circoviridae и Parvoviridae . Они реплицируются в ядре и во время репликации образуют промежуточную двухцепочечную ДНК. Анелловирус человека, называемый TTV, включен в эту классификацию и обнаруживается почти у всех людей, бессимптомно заражая их почти все основные органы.
Класс III: двухцепочечный РНК-вирус
Как и большинство РНК-вирусов, двухцепочечные РНК-вирусы не зависят от полимеразы-хозяина для их репликации, в отличие от ДНК-вирусов. Вирусы с двухцепочечной РНК изучены не так хорошо, как другие классы. Этот класс включает два больших семейства: Reoviridae и Birnaviridae . Репликация является моноцистронной и включает отдельные и сегментированные геномы, что означает, что каждый из генов кодирует только один белок, в отличие от других вирусов, которые демонстрируют более сложную трансляцию.
Классы IV и V: вирус с одноцепочечной РНК.
![]()
Эти вирусы состоят из двух разных типов, но оба разделяют тот факт, что их репликация происходит в основном в цитоплазме, и что репликация не так сильно зависит от клеточного цикла клетки-хозяина, как от ДНК-вирусов. Эти классы вирусов также входят в число наиболее изученных типов вирусов, наряду с вирусами с двухцепочечной ДНК.
Класс IV: вирус с одноцепочечной РНК с положительным смыслом
Вирусы с положительной смысловой РНК (и все гены, определяемые как положительные) могут быть напрямую доступны рибосомам хозяина для немедленного образования белков. Их можно разделить на две группы, каждая из которых реплицируется в цитоплазме:
- Полицистронные мРНК вирусов, в которых РНК из генома образует мРНК, а затем транслируется в полипротеиновый продукт, который затем расщепляется с образованием зрелых белков. Это означает, что ген может использовать несколько методов для получения белков из одной и той же цепи РНК, тем самым уменьшая размер своего генома.
- Сложные транскрипционные вирусы, которые могут использовать субгеномные мРНК, фазовый сдвиг рибосом и протеолитический процессинг полипротеинов. Все эти механизмы различны и позволяют производить белки из одной и той же цепи РНК.
Семейства Coronaviridae , Flaviviridae и Picornaviridae являются примерами вирусов, принадлежащих к этому классу.
Класс V: вирус с одноцепочечной РНК с отрицательным смыслом
Вирусы с отрицательной смысловой РНК (и все гены, определяемые как отрицательные) не могут быть напрямую доступны рибосомам хозяина для немедленного образования белков. Затем они должны быть расшифрованы вирусными полимеразами в дополнительном "читаемом" положительном направлении. Их также можно разделить на две группы:
- Вирусы, содержащие несегментированные геномы, для которых первой стадией репликации является транскрипция генома с отрицательной цепью с помощью РНК-зависимой вирусной РНК-полимеразы с образованием моноцистронных мРНК, которые кодируют различные вирусные белки. Затем создается копия генома с положительным смыслом, которая служит шаблоном для производства генома с отрицательной цепью. Репликация происходит в цитоплазме.
- Вирусы с сегментированными геномами, репликация которых происходит в цитоплазме и для которых РНК-зависимая вирусная РНК-полимераза продуцирует моноцистронные мРНК из каждого сегмента генома.
Семейства Orthomyxoviridae , Paramyxoviridae , Bunyaviridae , Filoviridae и Rhabdoviridae (включая бешенство ) являются примерами вирусов, принадлежащих к этому классу.
Класс VI: вирус с одноцепочечной РНК с положительным смыслом, использующий промежуточную ДНК для репликации
Хорошо изученное семейство вирусов этого класса включает ретровирусы . Одна из их отличительных черт - использование обратной транскриптазы для преобразования позитивно-смысловой РНК в ДНК. Вместо того, чтобы использовать РНК для создания паттернов трансляции белков, они используют ДНК для создания этих паттернов, которые встраиваются в геном хозяина с помощью интегразы . Затем репликация может начаться с помощью полимераз клетки-хозяина.
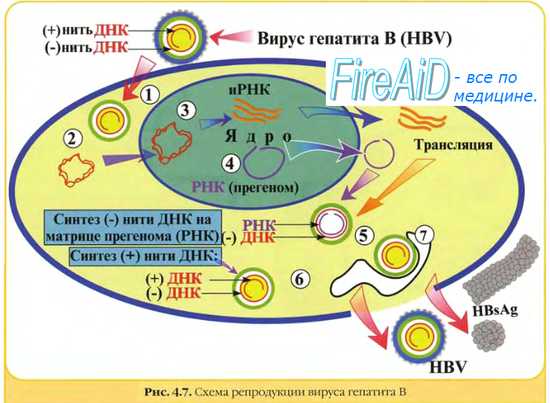
Класс VII: двухцепочечный ДНК-вирус с использованием промежуточной одноцепочечной РНК для репликации.
Эта небольшая группа вирусов, примером которой является вирус гепатита B , имеет вакантный двухцепочечный геном, который затем заполняется с образованием ковалентно замкнутой кольцевой ДНК (cfcDNA или cccDNA на английском языке), которая служит шаблоном для производство вирусных мРНК и субгеномных РНК. Прегеномная РНК служит матрицей для вирусной обратной транскриптазы и для производства ДНК-генома.
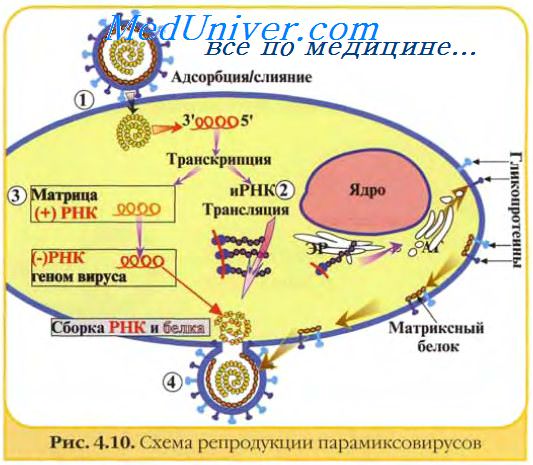
Репликация вирусных РНК молекул. Этапы репликации РНК генома вирусов.
Репликация вирусных РНК является уникальным феноменом. Существенное отличие механизма синтеза вирусных РНК от механизма синтеза клеточных РНК состоит в том, что в качестве матрицы в первом случае используется РНК, а во втором — ДНК.
Для транскрипции РНК на РНК-матрице необходима вирионная РНК-зависимая РНК-полимераза. Репликация вирусной РНК требует, прежде всего, синтеза комплементарной РНК, которая затем служит матрицей для производства большого количества вирусной РНК.
Когда вирусная РНК имеет отрицательную полярность (орто-, парамиксо-, рабдо-, фило-, борна-, арена- и буньявирусы), комплементарная РНК будет иметь положительную полярность и РНК-полимераза, подобно вирионной транскриптазе, используется для первичной транскрипции мРНК.
![репликация вирусной рнк у парамиксовирусов]()
Так как большинство транскриптов, синтезируемых на каждой вирусной (-)цепи РНК, являются молекулами субгеномной РНК, некоторые полноразмерные цепи служат матрицами для синтеза (репликации) вирусной РНК. Некоторые вирусы для транскрипции и репликации используют различные РНК-полимеразы, тогда как в других случаях одни и те же ферменты могут выполнять различные функции.
У многих РНК-вирусов, (пикорна-, калици-, астро-, тога-, флави-, корона-, артери-, нодавирусы) комплементарная РНК является отрицательно полярной. На одной комплементарной РНК-матрице может транскрибироваться одновременно несколько молекул вирусной РНК, а на каждом РНК-транскрипте начинается продукция полимеразы. Образуется структура, известная как реплика-тивный посредник, — частично двуцепочечная структура с одноцепочечными хвостами.
Для начала репликации РНК пикорнавирусов и калицивирусов, а также ДНК аденовирусов требуется небольшой белок, связанный ковалентно с 5'-концом вновь синтезированных (+) или (—) цепей РНК, так же как с родительской вирионной РНК, но не с мРНК.
Вновь синтезированные (+)РНК могут иметь разное назначение: включаться в репликативный комплекс и служить матрицей для синтеза комплементра-ных (—)РНК; выполнять функции мРНК; включаться в качестве генома в новые вирионы. Механизм, определяющий судьбу вновь синтезированных (+)РНК, не известен.
Ретровирусы имеют геномную (+) одноцепочечную РНК. В отличие от других РНК-вирусов, они реплицируются посредством ДНК-посредника. Вирионная обратная транскриптаза, используя РНК-молекулу как праймер, создает односпиральную ДНК-копию. Затем, функционируя как рибонуклеаза, тот же самый фермент удаляет родительскую молекулу РНК из ДНК-РНК-гибрида и копирует одноцепочечную ДНК-цепь, чтобы образовать линейную двуцепочечную ДНК, которая содержит дополнительную последовательность, известную как длинный концевой повтор (LTR) на каждом конце.
Эта двуцепочечная ДНК затем циркулирует и интегрирует с клеточной хромосомальной ДНК. Вирусная РНК транскрибируется с интегрированной (провирусной) ДНК.Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.В зависимости от типа генетического материала (ДНК или РНК), образование дочерних копий геномов протекает по-разному.
У ДНК-геномных вирусов репликация вирусных ДНК принципиально сходна с репликацией клеточных ДНК.
Репликацию РНК-геномных вирусов осуществляют вирусные РНК-зависимые РНК-полимеразы (репликазы). Исключение составляют ретровирусы, их +РНК служит матрицей для синтеза ДНК Синтез ДНК на матрице РНК осуществляет вирусная РНК-зависимая ДНК-полимераза (обратная транскриптаза), необходимая для переписывания информации с РНК на ДНК. Синтезируемая вирусная ДНК интегрируется в клеточный геном в форме ДНК-провируса.
![Репликация вируса в клетке. Сборка вирусов. Высвобождение дочерних вирионов из клетки.]()
Репликация однонитевых РНК вирусов. Репликация протекает в два этапа: первый включает образование матрицы, комплементарной геному; второй — образование копий РНК с этой матрицы. При репликации +РНК-вирусов количество копий -РНК (на матрице родительской нити +РНК) строго контролируется, а количество копий +РНК (с матрицы синтезированной нити -РНК) не контролируется.
Репликация двухнитевых РНК вирусов. В качестве матрицы для синтеза +РНК вирусные репликазы используют минус-нить РНК и наоборот. Часть молекул -РНК соединяется с +РНК и образует двухнитевую молекулу РНК, а другая часть молекул -РНК функционирует как матрица для синтеза мРНК.
Сборка вирусов
У просто устроенных вирусов, состоящих из нуклеиновой кислоты и нескольких белков, сборка состоит из упорядоченного взаимодействия этих молекул. У сложно устроенных вирусов сборка дочерних популяций протекает многоступенчато.
Взаимодействие нуклеиновых кислот с внутренними и оболочечными белками приводит к образованию нуклеокапсидов, или сердцевин. В процессе образования «одетых - вирусов полные нуклеокапсиды упорядочение выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками (М-белками). При нарушениях процесса самосборки могут образовываться пустые капсиды либо комплексы нуклеиновых кислот с внутренними белками.
![Репликация вируса в клетке. Сборка вирусов. Высвобождение дочерних вирионов из клетки.]()
Высвобождение дочерних вирионов из клетки
Высвобождение дочерних вирионов — конечная стадия репродуктивного цикла. Вирусы, лишённые суперкапсида, и поксвирусы обычно высвобождаются быстро; выход дочерних популяций сопровождается разрушением цитоплазматической мембраны (ЦПМ) и лизисом клетки. Вирусы, содержащие суперкапсид, высвобождаются медленнее. Модифицированные участки мембраны с заключёнными в них вирионами выпячиваются наружу и затем отпочковываются. Принцип высвобождения дочерних популяций почкованием во многом сходен с процессами, направленными на отторжение непригодного для клетки материала или обновление клеточных мембран. При высвобождении почкованием изменённая клетка иногда может сохранять жизнеспособность.
- Вернуться в оглавление раздела "Микробиология."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.Репликация вирусных ДНК. Механизм репликации вирусных молекул ДНК.
Синтез геномной вирусной ДНК в принципе не отличается от синтеза клеточной ДНК. У разных вирусов это происходит с использованием разных механизмов. Известно, что клеточные ДНК-полимеразы могут начинать синтез новой цепи ДНК только с короткого праймера.
Для решения этой проблемы ДНК-вирусы разных семейств пользуются различной стратегией. Одни из них имеют циркулярный ДНК-геном, другие —линейный геном с комплементарными концами, у третьих имеется белковый праймер на 5'-конце.
Для репликации вирусной ДНК необходимы некоторые кодируемые вирусом ферменты: геликаза (с АТФазной активностью), чтобы расплести двойную спираль; белок, дестабилизирующий двойную спираль до тех пор, пока каждая из нитей не будет скопирована; ДНК-полимераза для копирования каждой цепи в направлении от 5'-конца к З'-концу; РНКаза для разрушения РНК-праймера после его функционирования; ДНК-лигазы для соединения ДНК-фрагментов, если она синтезируется не целиком. Часто один большой фермент обладает двумя и более активностями.
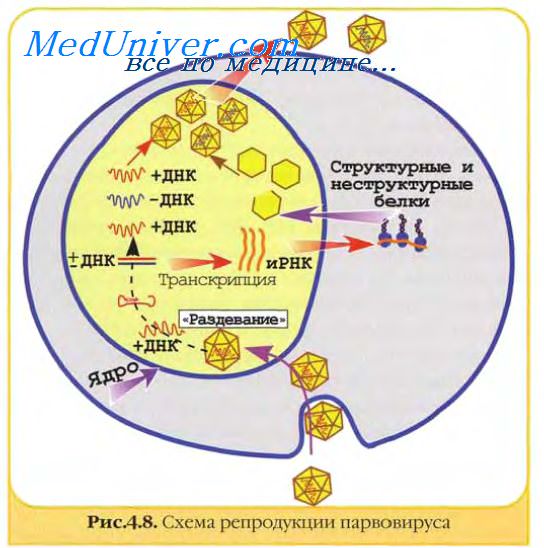
![репликация парвовируса]()
Геном парвовирусов использует для своей репликации клеточные ферменты, включая ДНК-полимеразу А. Ранний вирусный белок присоединяется к месту регуляторной последовательности вирусного генома и начинается репликация ДНК.
Происходит непрерывный и прерывистый синтез, соответственно, двух растущих цепей ДНК: главной и ответвляющейся. Прерывистый синтез ответвляющейся цепи связан с повторяющимися короткими олигонуклеотидными праймерами, поочередно рождающими короткие цепи ДНК (фрагменты Оказаки), которые затем соединяются ковалентно индуцированной вирусом ДНК-лигазой в одну растущую цепь.
Инициирующими единицами репликации папиллома- и полиомавирусов являются ковалентно связанные циркулярные (±)ДНК и линейные (±)ДНК адено- и герпесвирусов. Праймером синтеза аденовирусной ДНК служит предшественник белка, ковалентно связанный с вирионной ДНК. Репликация ДНК происходит непрерывно от 5'-конца к 3'-концу с использованием вирионной ДНК полимеразы. Синтез фрагментов Оказаки не происходит. Герпесвирусы кодируют большинство (если не все) белков, необходимых для репликации ДНК, включая ДНК-полимеразу, геликазу, праймазу, белок, соединяющий одноцепочечные ДНК, и белок, различающий начало репликации.
Вирусы оспы и асфаравирусы, которые размножаются целиком в цитоплазме, являются самодостаточными по механизму репликации. Гепаднавирусы, подобно ретровирусам, используют положительно полярные одноцепочечные РНК транскрипты в качестве посредников в синтезе ДНК с помощью обратной транскрипции. Синтез вирусной ДНК может происходить как в цитоплазме, так и в ядре зараженной клетки. Например, ДНК вирусов оспы синтезируется в цитоплазме, а ДНК герпес-, адено-, папиллома-, полиома- и парвовирусов — в ядре.
Делеция значительного количества генов (~ 40%) у оспо- и герпесвирусов не оказывает существенного влияния на их размножение в культуре клеток, возможно, что они важны для выживания этих вирусов в природе.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.В зависимости от типа генетического материала (ДНК или РНК), образование дочерних копий геномов протекает по-разному.У ДНК-содержащих вирусов, как в прокариотических, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК- содержащих вирусов сначала образуется комплементарная нить, - так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК.
Репликация вирусных РНК, как минус, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза. Это геномный белок, который есть у всех РНК-овых вирусов. Исключение составляют ретровирусы, их +РНК служит матрицей для синтеза ДНК. Синтез ДНК на матрице РНК осуществляет вирусная РНК-зависимая ДНК-полимераза (обратная транскриптаза), необходимая для переписывания информации с РНК на ДНК. Синтезируемая вирусная ДНК интегрируется в клеточный геном в форме ДНК-провируса.
Репликативная форма РНК минус-нитевых вирусов (плюс нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус нитей), но и выполняет функции и-РНК, т.е. идет на рибосомы и обеспечивает синтез вирусных белков (трансляция).
Репликация однонитевых РНК вирусов. Репликация протекает в два этапа: первый включает образование матрицы, комплементарной геному; второй — образование копий РНК с этой матрицы. При репликации +РНК-вирусов количество копий -РНК (на матрице родительской нити +РНК) строго контролируется, а количество копий +РНК (с матрицы синтезированной нити -РНК) не контролируется.
Репликация двухнитевых РНК вирусов. В качестве матрицы для синтеза +РНК вирусные репликазы используют минус-нить РНК и наоборот. Часть молекул -РНК соединяется с +РНК и образует двухнитевую молекулу РНК, а другая часть молекул -РНК функционирует как матрица для синтеза мРНК.
_________________________________________
добавка от Заикиной. Я не сравнивала содержание
3) У ДНК-содержащих вирусов, как в прокариотических, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК-содержащих вирусов сначала образуется комплементарная нить, так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК. При участии ДНК-зависимой РНК-полимеразы синтезируются и-РНК, которые поступают на рибосомы клетки, где и синтезируются вирусспецифические белки. У двунитевых ДНК-содержащих вирусов, геном которых транскрибируется в цитоплазме клетки хозяин. Вирусы, геномы которых транскрибируются в ядре клетки, используют содержащуюся там клеточную ДНК-зависимую РНК-полимеразу. Репликация вирусных РНК, как минус, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза. Репликативная форма РНК минус-нитевых вирусов (плюс нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус нитей), но и выполняет функции и-РНК. У плюс-нитевых РНК-содержащих вирусов функцию трансляции выполняют ее копии, синтез которых осуществляется через репликативную форму (минус нить) при участии вирусных РНК-зависимых РНК-полимераз. У некоторых РНК-содержащих вирусов имеется совершенно уникальный механизм транскрипции. Он обеспечивается специфическим вирусным ферментом - ревертазой (обратной транскриптазой), и называется обратной транскрипцией. Суть ее состоит в том, что в начале на матрице вирусной РНК при участии обратной транскриптазы образуется транскрипт, представляющий собой одну нить ДНК. На нем с помощью клеточной ДНК-зависимой ДНК-полимеразы синтезируется вторая нить и формируется двунитевой ДНК-транскрипт. С него обычным путем через образование и-РНК происходит реализация информации вирусного генома. Результатом описанных процессов репликации, транскрипции и трансляции является образование дочерних молекул вирусной нуклеиновой кислоты и вирусных белков, закодированных в геноме вируса. После этого наступает третий, заключительный период взаимодействия вируса и клетки. Из структурных компонентов на мембранах цитоплазматического ретикулюма клетки собираются новые вирионы. Вновь сформировавшиеся вирионы пассивно или активно покидают клетку и оказываются в окружающей ее среде. Таким образом, синтез вирусных нуклеиновых кислот и белков и сборка новых вирионов происходят в определенной последовательности и в разных структурах клетки в связи с чем способ репродукции вирусов и был назван дисъюнктивным.![]()
Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Читайте также: