
Что такое трансляция вирусов
Обновлено: 19.04.2024
Вирусные геномы кодируют синтезы двух классов белков: структурные белки входят в состав дочерних популяций, а неструктурные белки обслуживают процессы репродукции, но не входят в состав дочерних популяций (ингибиторы синтеза клеточных РНК и белков, протеазы и др.).
Трансляция РНК-содержащих вирусов. Поскольку вирусный геном кодирует несколько белков, то возможно два варианта трансляции:
каждый полипептид синтезируется отдельно от других (тога- и ретровирусы);
Некоторые вирусы используют оба этих механизма. Полипептиды, образующиеся при обоих вариантах трансляции, могут подвергаться посттрансляционной модификации (гликозилирование, фосфорилирование или сульфатирование).
Трансляция ДНК-содержащих вирусов. В трансляционных процессах доминирует трансляция отдельных мРНК, кодирующих индивидуальные полипептиды. В отдельных случаях (например, у аденовирусов) не менее трёх белков образуются путём нарезания общего полипептида-предшественника.
В зависимости от типа генетического материала (днк или рнк), образование дочерних копий геномов протекает по-разному.
У ДНК-геномных вирусов репликация вирусных ДНК принципиально сходна с репликацией клеточных ДНК.
Репликацию РНК-геномных вирусов осуществляют вирусные РНК-зависимые РНК-полимеразы (репликазы). Исключение составляют ретровирусы, их +РНК служит матрицей для синтеза ДНК Синтез ДНК на матрице РНК осуществляет вирусная РНК-зависимая ДНК-полимераза (обратная транскриптаза), необходимая для переписывания информации с РНК на ДНК. Синтезируемая вирусная ДНК интегрируется в клеточный геном в форме ДНК-провируса.
Репликация однонитевых РНК вирусов. Репликация протекает в два этапа: первый включает образование матрицы, комплементарной геному; второй — образование копий РНК с этой матрицы. При репликации +РНК-вирусов количество копий -РНК (на матрице родительской нити +РНК) строго контролируется, а количество копий +РНК (с матрицы синтезированной нити -РНК) не контролируется.
Репликация двухнитевых РНК вирусов. В качестве матрицы для синтеза +РНК вирусные репликазы используют минус-нить РНК и наоборот. Часть молекул -РНК соединяется с +РНК и образует двухнитевую молекулу РНК, а другая часть молекул -РНК функционирует как матрица для синтеза мРНК.
Сборка вирусов
У просто устроенных вирусов, состоящих из нуклеиновой кислоты и нескольких белков, сборка состоит из упорядоченного взаимодействия этих молекул. У сложно устроенных вирусов сборка дочерних популяций протекает многоступенчато.
Взаимодействие нуклеиновых кислот с внутренними и оболочечными белками приводит к образованию нуклеокапсидов, или сердцевин. В процессе образования «одетых - вирусов полные нуклеокапсиды упорядочение выстраиваются с внутренней стороны клеточной мембраны под участками, модифицированными оболочечными вирусными белками (М-белками). При нарушениях процесса самосборки могут образовываться пустые капсиды либо комплексы нуклеиновых кислот с внутренними белками.
Высвобождение дочерних вирионов из клетки
Высвобождение дочерних вирионов — конечная стадия репродуктивного цикла. Вирусы, лишённые суперкапсида, и поксвирусы обычно высвобождаются быстро; выход дочерних популяций сопровождается разрушением цитоплазматической мембраны (ЦПМ) и лизисом клетки. Вирусы, содержащие суперкапсид, высвобождаются медленнее. Модифицированные участки мембраны с заключёнными в них вирионами выпячиваются наружу и затем отпочковываются. Принцип высвобождения дочерних популяций почкованием во многом сходен с процессами, направленными на отторжение непригодного для клетки материала или обновление клеточных мембран. При высвобождении почкованием изменённая клетка иногда может сохранять жизнеспособность.
Интегративное взаимодействие, или вирогения, не приводит к гибели клетки. Нуклеиновая кислота вируса встраивается в геном клетки-хозяина и в последующем функционирует как его составная часть. Наиболее яркие примеры подобного взаимодействия — лизогения бактерий и вирусная трансформация клеток.
Интерференция вирусов происходит при инфицировании клетки двумя вирусами. Различают гомологичную (при инфицировании клетки родственными вирусами) и гетерологичную (если интерферируют неродственные виды) интерференцию. Это явление возникает не при всякой комбинации возбудителей, иногда два разных вируса могут репродуцироваться одновременно (например, вирусы кори и полиомиелита). Интерференция реализуется либо за счёт индукции одним вирусом клеточных ингибиторов (например, ИФН), подавляющих репродукцию другого, либо за счёт повреждения рецепторного аппарата или метаболизма клетки первым вирусом, что исключает возможность репродукции второго.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Синтез вирусных белков
Субстратом для синтеза вирусных белков являются аминокислоты, тождественные аминокислотам белков клетки. Матрицей при синтезе белков у ДНК содержащих вирусов является информационная РНК, которая формируется на вирусной ДНК. Информационные РНК синтезируются из субстратов зараженной клетки. Синтез матричной м РНК осуществляется ДНК зависимой РНК полимеразой, которая присутствует в зараженной клетке или приносится извне с вирусной частицей.
Процесс синтеза белков у РНК содержащих вирусов происходит без участия ДНК. Функцию вирусно формационной РНК выполняет вирусно цифическая РНК.
Как известно, важнейшим компонентом системы синтеза белка являются рибосомы. При изучении роли рибосом в синтезе вирусного белка было показано, что при заражении клеток вирусом приостанавливается процесс синтеза новых рибосом при этом высвобождаются старые рибосомы, имеющиеся в клетках, и на них осуществляется синтез белка.
Считается, что синтез белков у большинства вирусов происходит в цитоплазме.
Формирование (морфогенез) зрелых вирусных частиц и их выход из клетки. Зрелая вирусная частица (вирион) формируется из молекул нуклеиновой кислоты и белковых субъединиц. В состав вируса могут входить липопротеидные мембраны и другие компоненты.
Учитывая, что РНК- и ДНК-содержащие вирусы являются облигатными внутриклеточными паразитами, создание противовирусных препаратов является сложной задачей в химиотерапии вирусных инфекций. В процессе размножения вирусы используют аппарат биосинтеза клеток макроорганизма, изменяя его определенным образом. В связи с этим трудно избирательно воздействовать на вирус, не повредив при этом клетки макроорганизма.
Транскрипция вируса в клетке. Трансляция вирусов.
Транскрипция +РНК-содержащих вирусов. Функции мРНК выполняет геном (+РНК), поэтому у таких вирусов для синтеза вирусных белков (трансляция) нет необходимости в процессе транскрипции. Другими словами, у +РНК-содержащих вирусов транскрипция отсутствует.
Транскрипция -РНК-содержащих вирусов и вирусов, имеющие две нити РНК. Функции мРНК выполняют транскрипты, комплементарные -РНК вириона. Поэтому у таких вирусов транскрипция существует как самостоятельный этап репродуктивного цикла. Для образования транскриптов в составе вирионов имеется собственная РНК-полимераза (транскриптаза).
Транскрипция ДНК-вирусов. Транскрипция — самостоятельный этап репродуктивного цикла, так как геном ДНК-вирусов должен транскрибироваться для образования мРНК. Вирусы, репродуцирующиеся в ядре (например, герпес- и аденовирусы) для этой цели используют клеточную ДНК-зависимую РНК-полимеразу (транскриптазу). Вирусы, репродуцирующиеся в цитоплазме (например, поксвирусы) лишены такой способности и содержат (как и вирусы с -РНК) собственную транскриптазу.
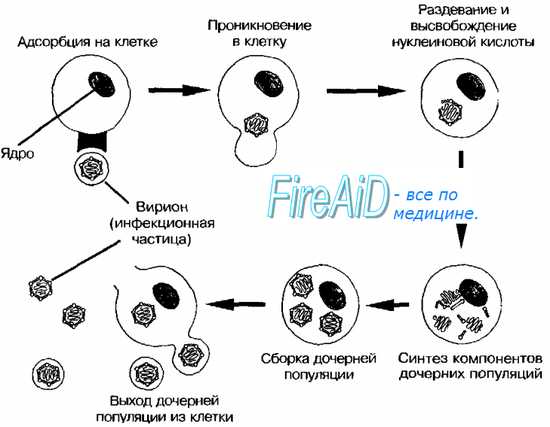
Рис. 2-3. Основные этапы репродукции вирусов.
Трансляция вирусов
Вирусные геномы кодируют синтезы двух классов белков: структурные белки входят в состав дочерних популяций, а неструктурные белки обслуживают процессы репродукции, но не входят в состав дочерних популяций (ингибиторы синтеза клеточных РНК и белков, протеазы и др.).
Трансляция РНК-содержащих вирусов. Поскольку вирусный геном кодирует несколько белков, то возможно два варианта трансляции:
1) каждый полипептид синтезируется отдельно от других (тога- и ретровирусы);
Некоторые вирусы используют оба этих механизма. Полипептиды, образующиеся при обоих вариантах трансляции, могут подвергаться посттрансляционной модификации (гликозилирование, фосфорилирование или сульфатирование).
Трансляция ДНК-содержащих вирусов. В трансляционных процессах доминирует трансляция отдельных мРНК, кодирующих индивидуальные полипептиды. В отдельных случаях (например, у аденовирусов) не менее трёх белков образуются путём нарезания общего полипептида-предшественника.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Транскрипция вирусов. Характеристика транскрипции вирусов.
У оц(+)РНК-вирусов репликация начинается с полной или частичной трансляции генома без промежуточных этапов. У всех других классов вирусов процесс экспрессии вирусного генома начинается с транскрипции мРНК.
В случае с ДНК-вирусами, реплицирующимися в ядре, эту функцию выполняет клеточная ДНК-зависимая РНК-полимераза II. Все другие вирусы нуждаются в уникальной и специфичной транскриптазе, которую кодирует вирус, и она является структурным компонентом вириона. Двуцепочечные ДНК-вирусы, которые размножаются в цитоплазме, имеют ДНК-зависимую РНК-полимеразу, тогда как двуцепочечные РНК-вирусы имеют РНК-зависимую РНК-полимеразу, специфичную для двуцепочечных РНК, а негативно полярные односпиральные РНК-вирусы носят РНК-зависимую РНК-полимеразу, специфичную для одноцепочечных РНК.
Транскрипция вирусных ДНК и РНК контролируется сложными регуляторными механизмами и, прежде всего, продуктами экспрессии регуляторных генов.
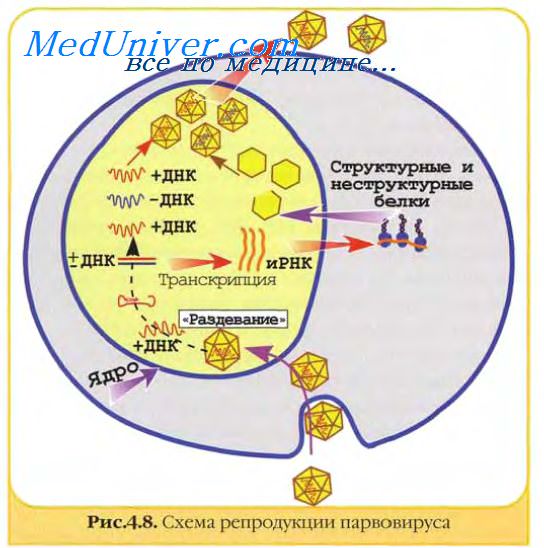
Транскрипты, которые образовались особенно в ранней стадии инфекции, считываются полностью на поздней стадии инфекции с образованием ряда длинных транскриптов с различными функциями.
Общими закономерностями транскрипции некоторых вирусов с дцДНК-геномом являются: транскрипция ранних и поздних генов с разных цепей ДНК, наличие перекрывающихся генов, разных рамок считывания и интронов.
У РНК-вирусов регуляция транскрипции в общем происходит менее сложно, чем у ДНК-вирусов. Временные различия транскрипции разных генов выражены не так отчетливо. У большинства семейств вирусов с оц(+)РНК геномная РНК служит мРНК, и для транскрипции-репликации РНК требуется только образование негативной цепи. Однако для ретровирусов и вирусов с несегментированным негативным РНК-геномом существут другие механизмы регуляции транскрипции.
Различные виды мРНК имеют различный период полураспада, что может также служить одним из возможных уровней регуляции вирусной репликации.
Первичные РНК-транскрипты, образующиеся на ДНК-геномах в ядре перед выходом в цитоплазму, претерпевают серию посттрансляционных изменений (кэпирование, аденилирование, метилирование, делетирование и сплайсинг).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Стратегия вирусного генома, использующего клеточный аппарат трансляции, должна быть направлена на создание механизма для подавления трансляции собственных клеточных иРНК и для избирательной трансляции вирусных иРНК, которые всегда находятся в значительно меньшем количестве, чем клеточные матрицы. Этот механизм реализуется на уровне специфического узнавания малой рибосомальной субъединицей вирусных иРНК, т. е. на уровне формирования инициирующего комплекса. Поскольку многие вирусы не подавляют синтез клеточных иРНК, в зараженных клетках возникает парадоксальная ситуация: прекращается трансляция огромного фонда функционально активных клеточных иРНК, и на освободившихся рибосомах начинается трансляция одиночных молекул вирусных иРНК. Специфическое узнавание рибосомой вирусных иРНК осуществляется за счет вирусспецифических инициаторных факторов.
Два способа формирования вирусных белков:
1) иРНК транслируется в гигантский полипептид-предшественник, который после синтеза последовательно нарезается на зрелые функционально активные белки; 2) иРНК транслируется с образованием зрелых белков, или белков, которые лишь незначительно модифицируются после синтеза.
2) иРНК транслируется с образованием зрелых белков, или белков, которые лишь незначительно модифицируются после синтеза.
Второй способ формирования белков характерен для ДНК-содержащих вирусов и большинства РНК-содержащих вирусов. При этом способе синтезируются короткие моноцистронные иРНК в результате избирательной транскрипции одного участка генома (гена).
Вирусные белки в процессе инфекции синтезируются в избыточном количестве. Так, только 35% всех вирусиндуцированных белков герпесвирусов включается в вирионы. Вирусные белки накапливаются и в ядре и в цитоплазме, однако это не говорит о месте их синтеза.
СБОРКА ВИРУСНЫХ ЧАСТИЦ
Синтез компонентов вирусных частиц в клетке разобщен и может протекать в разных структурах ядра и цитоплазмы. При таком дисъюнктивном способе репродукции образование вирусной частицы возможно лишь в том случае, если вирусные нуклеиновые кислоты и белки обладают способностью при достаточной концентрации узнавать друг друга в многообразии клеточных белков и нуклеиновых кислот и самопроизвольно соединяться друг с другом, т. е. способны к самосборке. В основе самосборки лежит специфическое белокнук-леиновое и белок-белковое узнавание, которое может происходить в результате гидрофобных, солевых и водородных связей, а также стерического соответствия. Белокнуклеиновое узнавание ограничено небольшим участком молекулы нуклеиновой кислоты и определяется уникальными последовательностями нуклеотидов в неко дирующей части вирусного генома. С этого узнавания участка генома вирусными капсидными белками начинается процесс сборки вирусной частицы. Присоединение остальных белковых молекул осуществляется за счет специфиче-ческих белок-белковых взаимодействий или неспецифических белокнуклеиновых взаимодействий.
В связи с разнообразием структуры вирусов животных разнообразны и способы формирования вирионов, однако можно сформулировать следующие общие принципы сборки.
1) У просто устроенных вирусов формируются прови-рионы, которые затем в результате модификаций белков превращаются в вирионы. У сложно устроенных вирусов сборка осуществляется многоступенчато. Сначала формируются нуклеокапсиды или сердцевины, с которыми взаимодействуют белки наружных оболочек.
2) Сборка сложно устроенных вирусов (за исключением сборки вирусов оспы и реовирусов) осуществляется на клеточных мембранах. Сборка ядерных вирусов происходит с участием ядерных мембран, сборка цитоплазмати-ческих вирусов — с участием мембран эндоплазматическои сети или плазматической мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы.

5) Сложно устроенные вирусы для построения своих частиц используют ряд элементов клетки-хозяина, например липиды, некоторые ферменты, у ДНК-геномного SV40 — гистоны, у оболочечных РНК-геномных вирусов — актин, а в составе ареновирусов обнаружены даже рибосомы. Клеточные молекулы несут определенные функции в вирусной частице, однако включение их в вирион может явиться и следствием случайной контаминации, как, например, включение ряда ферментов клеточных оболочек или клеточных нуклеиновых кислот.
Читайте также: