
Цитоплазмы нет в клетках вируса вызывающего корь
Обновлено: 26.07.2024
Воспалительные изменения на уровне субклеточных структур при вирусной инфекции.
Вызываемое вирусами развитие клинических проявлений болезни и видимые признаки воспаления есть конечный результат множества событий. Большая часть процессов, связанных с репликацией вируса, а следовательно, с начальными воспалительными реакциями на микроструктурном уровне, недоступна для распознавания на клиническом уровне. Эта особенность, характерная для многих вирусных инфекций и впервые описанная С.А. Mims и А.Н. Sharpe], представляет собой важную концепцию для понимания вирусного воспаления. Зачастую привычные признаки: краснота, припухлость, инфильтрация составляют лишь видимую с большим опозданием и весьма незначительную часть того воспаления, которое захватывает многочисленные структуры пораженного организма.
Основные этапы взаимодействия вирусов и клеток чрезвычайно кратко представлены в модифицированной таблице.
В правом столбце таблицы перечислены события, происходящие внутри клетки, именно они определяют ее конечную судьбу: гибель, трансформацию, резервирование персистентного вируса и т.д. В левом столбце приведены события на уровне организма или ткани, ведущие к заражению клеток и развитию защитных иммунных реакций. Эта схема значительно упрощена, многое зависит от специфики вируса и хозяина.
Респираторные вирусы чаще проникают в организм через верхние дыхательные пути. Первичное прикрепление вируса к эпителиальным клеткам зависит от множества факторов: его устойчивости к неспецифическим защитным факторам (протеолитические ферменты, слизь, движение ресничек, соли желчных кислот), к секреторным антителам, а также от наличия специфических комплементарных рецепторов на поверхности клетки, способных к воссоединению с рецепторами вируса.
Как показали исследования последних лет, комплементарность вирусных и клеточных рецепторов зачастую обусловлена генетическими маркерами организма: группой крови и HLA-принадлежностью. Так, самые тяжелые осложненные формы гриппа чаще возникают у носителей антигенов А-Н и HLA-8, склонность к персистированию превалирует у лиц с группой крови ВШ и HLA-A9. Таким образом, развитие тех или иных форм воспаления отчасти предопределено генетически. Не исключено, что каждому из респираторных вирусов соответствует свой излюбленный генотип.
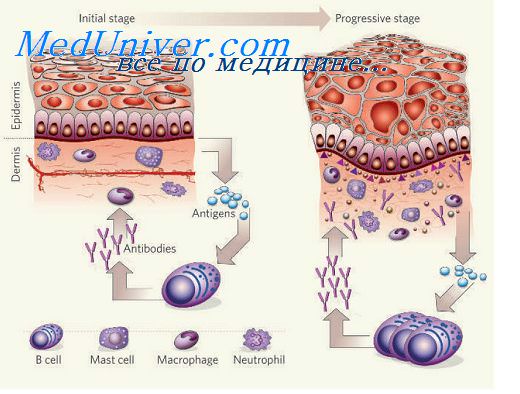
Вторым, не зависящим от специфических рецепторов, механизмом органотропности является протеолитическое расщепление гликопротеиновых оболочек парамиксовирусов (парагрипп, PC-вирус) в присутствии трипсиноподобных ферментов, содержащихся в инфицируемых клетках. Аналогичное свойство присуще вирусу гриппа, у которого ферментативному расщеплению подвергается гемагглютинин. При отсутствии в ткани необходимых ферментов многоцикловая инфекция не воспроизводится.
Прикрепление вируса к клетке является пусковым моментом для развития симптомокомплекса воспаления, который включает в себя альтерацию, экссудацию, трансформацию и пролиферацию в сочетании с нарушениями микроциркуляции и повреждениями функциональной активности. Краткая схема этого сложного и многообразного процесса не может отразить всех возможных вариантов и особенностей, но может облегчить восприятие основных моментов. Большая часть перечисленных в таблице 2 признаков воспаления, прежде всего деструкция и дистрофия, являются общими для всех респираторных вирусов и не могут служить надежными признаками дифференциации. Однако характер вирусных включений: их размеры, локализация, структура в сочетании с определенным типом поражения клеток (образование гигантских структур, пластов, скопления гроздевидных округлых клеток и т.д.) имеют определенное диагностическое значение при рассмотрении воспаления на субклеточном и клеточном уровне. Так, уже в начальной стадии воспаления, через 6-8 часов от момента заражения, при гриппе определяются базофильные включения, содержащие вирусные частицы. Чаще всего они локализуются в апикальной части эпителия; эозинофильные включения реже содержат вирус и чаще рассматриваются в качестве реактивных образований. При парагриппе и PC-инфекции выявляются цитоплазматические фуксинофильные включения, нередко группирующиеся вокруг ядра. При герпетической и цитомегаловирусной инфекции чаще регистрируются четко контурируемые внутриядерные включения, округлые, крупные, эозинофильные и оксифильные, нередко отгороженные от остальной цуклеплазмы светлой зоной. При аденовирусной инфекции также преобладают внутриядерные включения, на ранних стадиях инфекции (через 16 часов) - мелкие, округлые, базофильные или оксифильные; одни из них однородны, другие имеют форму колец с прозрачным центром. В дальнейшем число внутриядерных включений увеличивается, встречаются слоистые и ячеистые формы. На поздних этапах инфекции включения обнаруживаются и в цитоплазме клеток. Наиболее точная дифференцировка специфичности включений осуществляется с помощью прямого и непрямого иммунофлуоресцентного метода с использованием меченых антител.
Формирование вирусных включений интересно не только с точки зрения патогенных возможностей респираторных вирусов, но и с точки зрения анализа самого понятия воспаления. С одной стороны, это несомненный результат непосредственного воздействия вируса на клетку, вероятно это защитная реакция клетки, стремящейся локализовать патогенные частицы, а с другой стороны - это и возможность длительной резервации вируса. Таких двояких по целесообразности моментов в процессе вирусного воспаления достаточно. Так, под действием вирусов может происходить своеобразная активация фагоцитарной способности клеток, что проявляется в усилении цитофагоцитоза, когда инфицированные клетки активно фагоцитируют другие клетки, а также ферменты распавшихся структур. С одной стороны, это полезно с точки зрения утилизации и удаления дегенерировавших элементов, с другой стороны - нередко способствует большему распространению вирусной инфекции по клеточному пласту.
Большинство респираторных вирусов на ранних стадиях инфицирования вызывает ряд изменений клеточных структур, которые можно рассматривать как защитно-компенсаторные: набухание, гиперплазия ряда клеточных элементов, увеличение функционирующих локусов митохондрий, и лишь на более поздних стадиях возникают явления дистрофии, деструкции, либо необратимого перерождения.
Перечисляя изменения клеточных элементов, обусловленных вирусной репродукцией, было бы несправедливым забыть о главной их способности - нарушать синтез клеточных макромолекул. В случае развития вирусного инфицирования в клетке появляется два источника информации: клеточный и вирусный геномы. В подавляющем большинстве случаев ведущим оказывается вирусный геном, под руководством которого осуществляются процессы, направленные на воспроизводство вирусного потомства, в то время как синтез клеточных макромолекул (РНК, ДНК, белка) претерпевает значительное подавление. После длительных экспериментальных поисков было доказано, что в основе ингибирования синтеза клеточных РНК и ДНК в инфицированных клетках лежит феномен блокирования процесса транскрипции, т.е. "считывания" информации, закодированной в клеточном геноме, без нарушения целостности структур ДНК и РНК, а также РНК-полимераз. Ингибитором, ответственным за этот процесс, оказался аргининсодержащий белок - гистон. Большая часть исследователей полагает, что синтез белков-ингибиторов осуществляется самой клеткой в результате реализации информации, закодированной в ее геноме. Вирус лишь стимулирует синтез этих ингибиторов. Помимо подавления синтеза клеточных РНК, гистоны подавляют функцию и синтез ДНК, а также синтез АТФ и перенос аминокислот. Таким образом, появление индуцированных вирусом и воспроизведенных клеткой гистонов является одной из основных причин нарушения синтеза клеточных структур, а, следовательно, и дистрофии, являющейся одним из признаков воспаления.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Что такое корь? Причины возникновения, диагностику и методы лечения разберем в статье доктора Александрова Павла Андреевича, инфекциониста со стажем в 14 лет.
Над статьей доктора Александрова Павла Андреевича работали литературный редактор Маргарита Тихонова , научный редактор Сергей Федосов

Определение болезни. Причины заболевания
Корь (Measles) — острое инфекционное заболевание, вызываемое вирусом кори, который поражает эпителий верхних дыхательных путей и кожные покровы. Клинически характеризуется выраженным синдромом общей инфекционной интоксикации, патогномоничной энантемой, макуло-папулёзной сыпью, конъюнктивитом и синдромом поражения респираторного тракта (дыхательных путей).
Этиология
вид — возбудитель кори (Polinosa morbillarum)
Синдромально заболевание было описано в 1890 году доктором Бельским (Псков), в 1895 году детским доктором Филатовым и в 1898 году доктором Копликом (США).
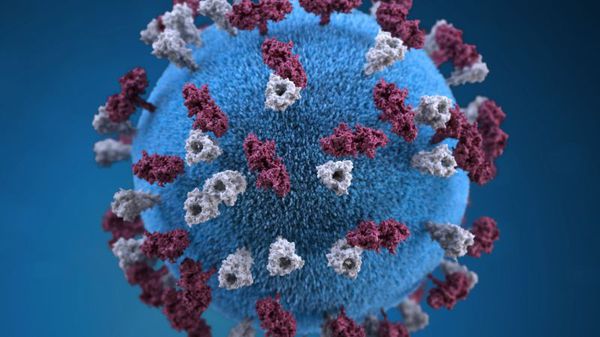
Оболочка вируса имеет 3 слоя: белковую мембрану, липидный слой и наружные гликопротеидные комплексы, образующие своеобразные выступы. Штаммы вирусов идентичны в антигенном отношении, обладают комплементсвязывающими, гемагглютинирующими, гемолизирующими свойствами и симпластообразующей активностью. СД-46 является рецептором у человека к вирусу кори.
Вирус неустойчив во внешней среде – чувствителен к ультрафиолету, в каплях слюны погибает за 30 минут, при высыхании и действии дезинфицирующих средств — мгновенно. Хорошо переносит пониженные температуры. Вирус можно выделить из различных сред организма (кровь, моча, кал, смывы со слизистых ротоглотки, конъюнктив, цереброспинальной жидкости). [2] [5]
Эпидемиология
Антропоноз. Источник заболевания – исключительно заражённый человек, также с атипичной формой заболевания кори. Инфицированный заразен с последних 2-х дней инкубации и до 4-го дня с момента появления высыпаний включительно — с 5 дня заразность пропадает.
Передача заболевания
Механизм передачи: воздушно-капельный (путь аэрозольный), редко трансплацентарный (при заболевании женщины в конце беременности). Теоретически возможно заражение от недавно привитых (но на практике такое случается чрезвычайно редко).
Существует весенне-зимняя сезонность. После перенесённой инфекции, полного курса вакцинации развивается стойкий пожизненный иммунитет.
Корь — одна из основных причин смерти среди детей раннего возраста.
В 2015 году в глобальных масштабах произошло 134 200 случаев смерти от кори. [1] [3]
Кто подвергается риску
Заразиться корью может любой человек, не имеющий иммунитета — тот, кто не был вакцинирован или тот, у кого не выработался иммунитет после вакцинации.
Самому высокому риску заболевания корью и развития осложнений, включая смертельный исход, подвергаются невакцинированные дети раннего возраста. Корь — одна из основных причин смерти среди них.
При обнаружении схожих симптомов проконсультируйтесь у врача. Не занимайтесь самолечением - это опасно для вашего здоровья!
Симптомы кори
Инкубационный период
Инкубационный период при типичной форме — от 9 до 11 суток (в некоторых случая до 13).
Симптомы кори у взрослых
Начало заболевания подострое (т.е. основной синдром появляется на 2-3 сутки от начала заболевания), однако при должной подготовке врача (выявлении патогномоничной энантемы — сыпи на слизистых оболочках) можно определить острое начало (в течении первых суток). Во взрослом возрасте из-за особенностей иммунной системы эти критерии могут не соблюдаться.
- синдром выраженной общей инфекционной интоксикации;
- синдром патогномоничной энантемы (пятна Бельского-Филатова-Коплика);
- синдром макуло-папулёзной экзантемы;
- синдром конъюнктивита (выраженный);
- синдром поражения респираторного тракта (фарингит, трахеобронхит);
- синдром генерализованной лимфаденопатии (ГЛАП);
- гепатолиенальный синдром.
Описана так называемая ранняя диагностика кори, включающая:
- затруднение носового дыхания;
- покраснение правого века;
- субфебрилитет (лихорадка постоянного типа).
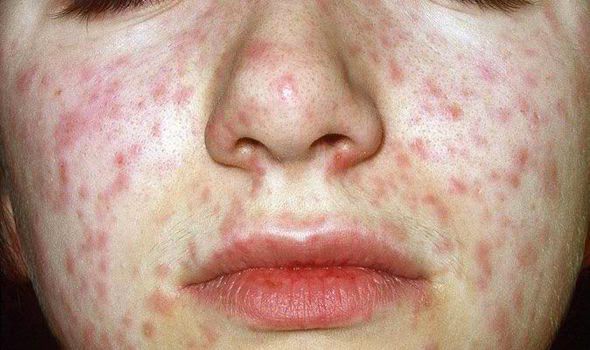
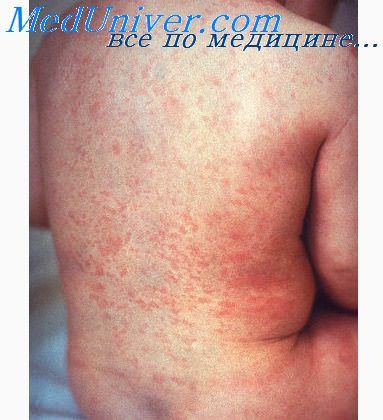
Объективно: можно отметить возникновение макуло-папулёзной сыпи, появляющейся в типичных случаях на 3-4 день болезни, с выраженной этапностью (лицо, шея; после туловище, руки, бёдра; затем олени и стопы, а на лице в этот момент бледнеет). Высыпания представляют собой папулы, окруженные красным пятном, предрасположенные к слиянию (но при наличии чистых участков), иногда появляются петехии. Спустя 3-4 дня сыпь становятся бледной и оставляет после себя бурые пятна и отрубевидное шелушение. Увеличиваются и приобретают чувствительность периферические лимфоузлы (затылочные, заднешейные и подмышечные). Ярко выражен конъюнктивит (конъюнктивы масштабно гиперемированы, отёчны, быстро присоединяется гнойное отделяемое). Характерным является вид больного: одутловатое лицо, красные (как у кролика) глаза, отёчность носа и век, сухие потрескавшиеся губы. При аускультации в лёгких прослушиваются сухие хрипы. Выражена тахикардия, снижение артериального давления. Увеличиваются размеры печени и селезёнки (генерализация инфекции).
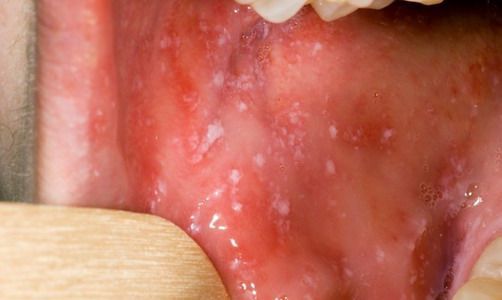
При фарингоскопии в ротоглотке визуализируется гиперемия мягкого нёба, на слизистой поверхности щёк в начале заболевания пятна Бельского — Филатова — Коплика (мелкие белесоватые пятнышки с узкой красноватой каёмкой), которые пропадают при появлении экзантемы. Иногда появляется пятнистая энантема на слизистой мягкого нёба.
Описана митигированная корь (у людей, получивших специфический противокоревой иммуноглобулин в период инкубации), характеризующаяся удлинением инкубационного периода до 21 дня, лёгким течением.
При абортивной форме заболевания характерно типичное начало, но спустя 1-2 суток клинические проявления исчезают.
Существует понятие — реакция на прививку (при первичном введении живой противокоревой вакцины), характеризующееся повышением температуры тела, катаральными явлениями (воспалениями) со стороны верхних дыхательных путей, скудной малозаметной сыпью (чаще развивается у детей и лиц с иммунодефицитом).
У больных с наличием тяжелого иммунодефицита – СПИДа (ВИЧ, онкологические больные, лица, получающие системные цитостатики после трансплантации органов) — течение заболевания крайне тяжёлое (летальность до 80%). [4] [5]
Корь. Вирус кори. Свойства вируса кори. Эпидемиология кори. Антигены вируса кори.
Корь — острая инфекция, проявляющаяся интоксикацией, катаральными явлениями, своеобразной энантемой и папулёзно-пятнистой сыпью.
Вирус кори — типовой вид рода Morbillivirus [от лат. mоrbilli, корь]. В его состав также включены патогенные для человека вирусы под острого склерозирующего панэнцефалита и рассеянного склероза.
Впервые вирус кори выделили Д. Эндерс и Т. Пиблз (1954). Вирус кори имеет сферическую форму, диаметр— 150-250 нм. Геном вируса кори образован несегментированной однонитевой молекулой -РНК.
С ней ассоциированы нуклеокапсидный белок NP и полимеразные белки Р и L, образующие нуклеокапсид со спиральным типом симметрии вируса кори. Снаружи он окружён матриксным М-белком.
Оболочка вируса кори образована двойным слоем липидов, включающим гликопротеипы Н (гемагглютинин) и F (белок слияния). В культуре клеток вирус кори даёт характерный цитопатический эффект с образованием гигантских клеток и/или синцитиев, либо образует зернистые тельца включений в цитоплазме и ядре.
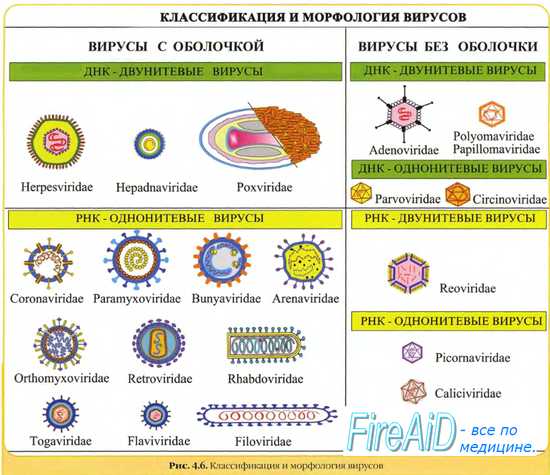
Эпидемиология кори
Корь распространена повсеместно, а в некоторых странах она эндемична. Резервуар кори — больной человек.
Возбудитель кори передаётся воздушно-капельным путём. Наибольшую эпидемическую опасность больной представляет в продромальный период и период кожных высыпаний. Вирус кори нестоек во внешней среде, чувствителен к инсоляции, высоким температурам и быстро разрушается под действием дезинфектантов и детергентов.
Антигенная структура вируса кори
Основные антигены вируса кори — гемагглютинин, белок F и нуклеокапсидный белок NP. AT к гемагглютинину и F-протеину проявляют цитотоксическое действие, направленное против инфицированных клеток.
Вирус кори имеет общие антигенные детерминанты с прочими морбилливирусами. Антигенная структура вируса кори стабильная. Все известные штаммы принадлежат к одному серологическому варианту.
Видео этиология, патогенез кори
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Морфология кори у детей. Экзантема при кори
Некрозы в гортани и трахее при кори неглубоки, поэтому при заживлении рубцы не образуются. В исключительных случаях на их месте отмечается избыточное разрастание грануляционной ткани с последующим развитием множественных полипов или папиллом. При неосложненной кори наблюдается интерстициалыгая пневмония с многоядерными гигантскими клетками в стенке альвеол.
В результате виремии и генерализации процесса развиваются специфичные для кори поражения слизистых оболочек (энантема) и кожи (экзантема). Слизистая оболочка полости рта поражается еще в период продромы кори — появляются участки полнокровия, отека и очагового некроза эпителия с последующим паракератозом, благодаря чему они приобретают характерный беловатый цвет.
Такие участки, располагающиеся на слизистой оболочке щек соответственно малым коренным зубам, патогномоничны для кори и называются пятнами Вельского — Филатова — Коплика. Они появляются раньше высыпаний на коже и .поэтому имеют большое диагностическое значение.
Экзантема при кори в виде крупнопятнистой папулезной сыпи характеризуется этапностью высыпания: в 1-е сутки она появляется за ушами, на переносице, лице, шее и верхней части груди, на 2-е — распространяется на туловище и верхние конечности и на 3-й — на нижние конечности. Микроскопически коревая сыпь представляет собой очажки неспецифического воспалительного процесса в сосочковом слое кожи в виде отека, гиперемии, иногда кровоизлияний с умеренной лимфогистиоцитарной инфильтрацией вокруг сосудов, волосяных каналов и выводных протоков потовых желез.
В эпидермисе отмечаются отек, вакуолизация цитоплазмы и ядер клеток, приводящая местами к гибели целых групп клеток и развитию так называемых фокусных некрозов эпителия. В дальнейшем происходит усиленное и полное ороговение (паракератоз) эпителиальных клеток в пределах пораженных участков.
При стихании воспалительных изменений нарастающий нормальный эпителий вызывает отторжение ороговевших частиц с явлениями паракератоза, что сопровождается очаговым (соответствующим размерам каждого пораженного участка) отрубевидным шелушением. На коже в период выздоровления в течение нескольких дней определяется очаговая бурая пигментация.
Поражение лимфоидного аппарата имеет системный характер, и одним из важнейших признаков является образование многоядерных гигантских клеток в миндалинах, лимфатических узлах, селезенке, аппендиксе. Обнаружение этих клеток в удаленных миндалинах позволяет поставить диагноз кори еще до появления сыпи.
Впервые многоядерные гигантские клетки описали в 1931 г. A. Worthin и W. Finkeldey, обнаружившие их независимо друг от друга в удаленных миндалинах у детей, находящихся в инкубационном или продромальном периодах кори. По поводу генеза этих клеток существуют различные предположения. И. Г. Шройт (1970), изучив сравнительную патологию коревой инфекции и динамику образования гигантских клеток, пришел к выводу, что существуют два основных типа гигантских многоядерных структур (размером от 15 до 100 нм и более), содержащие: 1) ядра ретикулярных клеток и 2) ядра лимфоцитов. Перед высыпанием в гигантских клетках содержатся и ядра плазматических клеток.
Эти данные подтвердили при электронно-микроскопическом изучении гигантских клеток Т. Masanori и соавт. (1976), выявившие у 61 клетки из 76 признаки лимфоцита, у 15 — признаки ретикулярной клетки. Ядра часто соединены мостиками, представляющими собой фрагменты мембраны ядра. Авторы полагают, что возникновение гигантских клеток связано с распадом ядер и слиянием клеток.
По данным А. С. Козлюка (1969), многоядерные гигантские клетки появляются в лимфоидиых органах обезьян в продромальном периоде кори и исчезают с появлением сыпи в течение 24—48 ч. Известно, что гигантские клетки при кори подвергаются дистрофическим изменениям. М. А. Скворцов считает, что гигантоклеточная реакция наиболее выражена в регионарных лимфоидных органах, по отношению к месту вхождения и фиксации вируса. Во всех лимфоидных органах при кори наблюдаются выраженные морфологические сдвиги, свидетельствующие о развитии противоинфекционного иммунитета.
Б. С. Гусман, изучая органы иммуногенеза у детей, умерших от кори (1975), обнаружила в лимфатических узлах увеличение центров размножения фолликулов, набухание эндотелия сосудов и синусов лимфоидных органов, усиление фагоцитарной активности макрофагов, повышение митотической активности ретикулярных клеток и накопление ШИК-положительных веществ в их цитоплазме, пазматизацию В-зависимых зон, повышение активности щелочной фосфатазы и оксидазы, пролиферацию эндотелия с десквамацией в просвет синусов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Цитоплазмы нет в клетках : а) вируса, вызывающего корь ; б) гриба подберёзовика ; в) большого пестрого дятла ; г) подрожника мне нужно правильный ответ на тест.

Цитоплазмы нет у вируса.
Так как это вообще неклеточная форма жизни.
У всех остальных организмов, имеющих клеточной строение цитоплазма в клетке является обязательным компонентом.
Ответ : а) вируса, вызывающего корь.

Какой тип биотических отношений устанавливается между большим пестрым дятлом и малым пестрым дятлом обитающими в одной экосистеме хвойного леса?
Какой тип биотических отношений устанавливается между большим пестрым дятлом и малым пестрым дятлом обитающими в одной экосистеме хвойного леса?
Где находится Цитоплазма в клетке?
Где находится Цитоплазма в клетке!
Срочно ответе правильно!

Цитоплазмы нет в клетках : а) крапивы б) подосиновика в) холерного вибриона г) вируса, вызывающего бешенство?
Цитоплазмы нет в клетках : а) крапивы б) подосиновика в) холерного вибриона г) вируса, вызывающего бешенство.

Цитоплазмы нет в клетках : 1)крапивы 2)подосиновика 3)холерного вибриона 4)вируса, вызывающего бешенство?
Цитоплазмы нет в клетках : 1)крапивы 2)подосиновика 3)холерного вибриона 4)вируса, вызывающего бешенство.

Какой тип биотических отношениях устанавливается между большим пестрым дятлом и малым пестрым дятлом обитающих в одной экосистеме хвойного леса?
Какой тип биотических отношениях устанавливается между большим пестрым дятлом и малым пестрым дятлом обитающих в одной экосистеме хвойного леса?

Особенности раст?
Клтки, вирусов, бактерий и грибов.
При каких условиях вирус проникает в цитоплазму клетки?
При каких условиях вирус проникает в цитоплазму клетки?
Для чего Большому пестрому дятлу острые когти?
Для чего Большому пестрому дятлу острые когти?

Цитоплазма у животных, грибы и бактериальная клетка?
Цитоплазма у животных, грибы и бактериальная клетка.
Из чего состоит.

Какой гриб считается пластинчатым из названных : подосиновик маслёнок лисичка подберёзовик?
Какой гриб считается пластинчатым из названных : подосиновик маслёнок лисичка подберёзовик.
Помогите пожалуйста это тест!
Вы находитесь на странице вопроса Цитоплазмы нет в клетках : а) вируса, вызывающего корь ; б) гриба подберёзовика ; в) большого пестрого дятла ; г) подрожника мне нужно правильный ответ на тест? из категории Биология. Уровень сложности вопроса рассчитан на учащихся 5 - 9 классов. На странице можно узнать правильный ответ, сверить его со своим вариантом и обсудить возможные версии с другими пользователями сайта посредством обратной связи. Если ответ вызывает сомнения или покажется вам неполным, для проверки найдите ответы на аналогичные вопросы по теме в этой же категории, или создайте новый вопрос, используя ключевые слова: введите вопрос в поисковую строку, нажав кнопку в верхней части страницы.
Тополь. Именно это дерево продуцирует наибольшее количество кислорода, вместе с тем перерабатывая углекислый газ, содержащийся в атмосфере.

Потому что на нем появляются спорангии, в которых образуются споры.
Обусловлено строением плазматической мембраны, то есть транспорт осуществляется выборочно, так как мембрана состоит из билипидных слоев, которые в ствою очередь гидрофобны и пропускают только небольшие малекулы и незаряженные частицы, также способны ..
2. Обусловленная введением лечебной сыворотки.
Кількість Ц = кількості Г = 20% А = Т = 100 - 40 = 60 / 2 = 30%.

Сердце стучит. Тихо. Одно. Нам на двоих – только мое… Так начинается синтез белка В клетках, мое - твое ДНК.
Так как черная откраска доминирует, следовательно А - ген черной окраски, а - ген белой окраски. В потомсте получено расщепление, значит носитель доминантного гена - гетерозиготен. Построим схему скрещивания Аа (кот) х аа (кошка) = Аа (черный котен..
1. Если имели в виду корень, то зона деления и происходит быстрое деление клеток, и также на верхушечной почке. 2. Стебель порядковое строение : Кора - Луб - Камбий - Древесина - Сердцевина 3. В зоне деления находятся молодые клетки и постоянно дел..
Размеры печеночного сосальщика взрослых особей достигают около сантиметра в ширину и трех сантиметров в длину. Рот характеризуется свойством всасывания и имеет форму трубки. Гельминт имеет одностороннюю систему переваривания, при которой одно единс..
1) ДНК : ТТГ - ГАА - ААА - ЦГГ - АЦТ тРНК : ААЦ - ЦУУ - УУУ - ГЦЦ - УГА Центральный участок тРНК антикодон УУУпо принципу комплементарности будет соответствовать кодону иРНК ААА По таблице генетического кода на основе кодона иРНК находим аминокислоту..
© 2000-2022. При полном или частичном использовании материалов ссылка обязательна. 16+
Сайт защищён технологией reCAPTCHA, к которой применяются Политика конфиденциальности и Условия использования от Google.
Читайте также: