
Действие биологических веществ на вирусы
Обновлено: 19.04.2024
Для терапии таких заболеваний, как астма, атеросклероз, системная красная волчанка, псориаз, ревматоидный артрит, рассеянный склероз и другие, такие как вирус натуральной оспы, используются человеческие антитела, блокирующие белковые факторы воспаления. Оказалось, что для этих целей можно использовать и белки инфекционных микроорганизмов, которые эволюционно адаптированы к преодолению защитных систем организма.
Особый интерес для медицинской биотехнологии представляют высоко патогенные вирусы, для которых человек является единственным хозяином. И сегодня на основе белков вируса натуральной оспы, столетиями являвшегося настоящим бичом цивилизаций, создаются препараты для коррекции тяжелейших патологических состояний неинфекционной, в том числе аутоиммунной, природы
Микроорганизмы обитают везде: в воздухе и в воде, в почве и в наших собственных телах… По сравнению с человеком и животными размножаются они чрезвычайно быстро, и их сообщества неизмеримо многочисленнее. Так, если человечество перевести в масштаб микромира, то все 7 млрд человек легко уместились бы в стандартной микробиологической колбе.
Специфическая защита организма против конкретного инфекционного агента – адаптивный или приобретенный иммунитет, развивается медленнее и представляет собой сложное взаимодействие иммунных клеток разного типа, регулируемое специальными белками.
ИММУННАЯ ОБОРОНА
Задача иммунной системы организма – защищать его от любых внешних и внутренних угроз.
Врожденный иммунитет неспецифичен – он обеспечивает защиту от любых инфицирующих агентов: вирусов, бактерий, а также макромолекул.
Одной из первых (возможно, и одной из самых древних) линий такой неспецифической защиты является апоптоз – программируемое самоубийство клетки. В случае вирусной инфекции апоптоз, индуцируемый в результате молекулярного распознавания специфичных молекул патогена, предотвращает размножение вируса и инфицирование других клеток организма хозяина.
Важную роль в ранней неспецифической защите организма от вирусной инфекции играет воспаление, возникающее в первые часы и дни после инфицирования и направленное на ограничение распространения вируса. Клетки врожденной иммунной системы млекопитающих – макрофаги, дендритные клетки и натуральные киллеры, в ответ на инфекцию продуцируют так называемые провоспалительные цитокины, такие как интерлейкин-1β(IL-1β), интерлейкин-18 (IL-18), фактор некроза опухолей (TNF) и гамма-интерферон (γ-IFN). В развитии воспалительной реакции участвуют белки и другого класса – хемокины, которые регулируют перемещение и функции лейкоцитов.
К дополнительным механизмам врожденного иммунитета можно отнести комплемент – многокомпонентный набор сложных белков сыворотки крови, которые являются протеолитическими ферментами, участвующими в инактивации инфекционных агентов и зараженных ими клеток.
Специфическая защита организма против конкретного инфекционного агента – адаптивный или приобретенный иммунитет, развивается медленнее реакций врожденного иммунитета. Он осуществляется в результате сложного взаимодействия клеток разного типа, контролируемого цитокинами TNF, IL-1β и γ-IFN. Его результат – появление B-лимфоцитов, синтезирующих специфичные противовирусные антитела, и вирусоспецифических цитолитических T-лимфоцитов. Специфичные антитела могут взаимодействовать с вирусными частицами и их компонентами индивидуально либо в комплексе с комплементом, инактивируя их.
Таким образом, интерлейкин-1β, фактор некроза опухолей и гамма-интерферон являются важнейшими цитокинами, которые, наряду с регулированием воспалительных реакций, контролируют развитие адаптивного иммунного ответа организма на инфекцию.
Но что если использовать для этих целей белки патогенных микроорганизмов, таких как вирусы, эволюционно-адаптированных к преодолению защитных систем организма, обернув, образно говоря, зло во благо?
Патогенный и антропонозный
Царство вирусов было открыто более ста лет назад выдающимся русским ученым Д. И. Ивановским, однако изучить эти мельчайшие организмы удалось лишь в последние десятилетия благодаря бурному развитию инструментальных методов исследований.
Разные вирусы млекопитающих отличаются не только по размеру генома и самих вирусных частиц, но и по стратегии своего развития в организме хозяина. Вирусы различных семейств демонстрируют удивительное разнообразие в механизмах преодоления систем врожденного и адаптивного иммунитета млекопитающих. Поэтому изучение этих особенностей вирусов позволяет выявить новые закономерности организации и функционирования защитных систем животных и человека, обеспечивающих выздоровление после инфицирования болезнетворным агентом.
Удивительный пример высокой патогенности для человека и строгой антропонозности представляет собой вирус натуральной оспы. Этот вирус, предположительно, исходно имел широкий круг хозяев, однако в процессе эволюции утратил способность размножаться в организме других млекопитающих, сохраняясь в эндемичном (т. е. свойственном только данной местности) состоянии в течение многих столетий в густонаселенных районах, прежде всего, Индийского субконтинента (Щелкунов, 2012).
Оружие против воспаления
Как уже упоминалось, главной причиной аутоиммунных заболеваний является разбалансировка иммунитета, в том числе сверхсинтез веществ, провоцирующих воспалительные реакции.
Одним из ключевых цитокинов иммунного воспалительного ответа является фактор некроза опухолей (TNF) – именно его повышенная продукция приводит к таким болезням аутоиммунной природы, как псориаз, болезнь Крона, ревматоидный артрит и т. п. Высокий уровень продукции TNF обусловливает и тяжелую патологию, часто завершающуюся летальным исходом – септический или эндотоксический шок.
Воспрепятствовать связыванию TNF с его белковыми рецепторами, зафиксированными на клеточных мембранах, можно с помощью соответствующих моноклональных антител либо так называемых растворимых форм его рецепторов, которые представляют собой внеклеточные домены (участки) рецепторных белков и обычно обнаруживаются в сыворотке и других биологических жидкостях. Присоединение подобных молекул к TNF должно инактивировать этот цитокин и уменьшить вызванный им воспалительный процесс.
Действительно, в модельных лабораторных экспериментах было доказано терапевтическое действие анти-TNF антител при лечении ряда патологий, обусловленных повышенной продукцией TNF. Однако попытки использовать для этих целей напрямую растворимые клеточные TNF-рецепторы не дали положительных результатов. Успеха удалось добиться лишь с помощью методов генетической инженерии, когда были созданы так называемые химерные белки, состоящие из TNF-связывающей части клеточных рецепторов и фрагмента иммуноглобулина человека.
Однако клинические исследования показали, что пациенты, страдающие ревматоидным артритом или другим воспалительным или аутоиммунным заболеванием, оказываются избирательно чувствительны лишь к одному из этих анти-TNF препаратов. Кроме того, поскольку все эти препараты имеют белковую природу, они сами по себе являются мишенью для иммунной системы больных, поэтому при длительной терапии их эффективность может снижаться. Это означает, что при потере чувствительности к одному препарату его требуется заменить на другой.
РЕВМАТОИДНЫЙ АРТРИТ — системное аутоиммунное заболевание соединительной ткани, проявляющееся главным образом хроническим воспалением суставов: голеностопных, лодыжек, коленей и кистей рук.
Сегодня от этой болезни страдает каждый сотый житель Земли, т. е. более 70 млн человек. Женщины болеют в несколько раз чаще, чем мужчины. Болезнь обычно развивается после 30 лет. В 70 % случаев ревматоидный артрит приводит к инвалидности, которая наступает довольно рано.
Причины возникновения этой болезни точно не установлены, но известно, что толчком к ее развитию служит сбой в иммунной системе в результате переохлаждения, стрессов, травм суставов и инфекции, в том числе ОРЗ, ангины и гриппа. При ревматоидном артрите наблюдается избыточный синтез таких провоспалительных цитокинов, как фактор некроза опухолей и гамма-интерферон
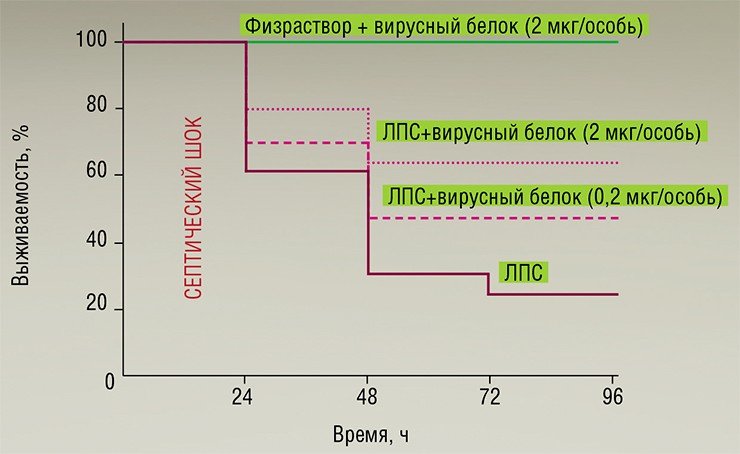
Методами генетической инженерии здесь были созданы рекомбинантные бакуловирусы, способные продуцировать в клеточной культуре TNF-связывающий белок (CrmB), характерный для вирусов оспы коров, оспы обезьян и натуральной оспы. Однако на экспериментальной модели эндотоксического шока было показано, что существенным терапевтическим эффектом обладает лишь белок CrmB вируса натуральной оспы (Gileva et al., 2006). Именно этот белок или его реконструированные варианты могут стать действующим началом новых средств анти-TNF терапии (Гилева и др., 2009).
На сегодня имеется много данных о том, что воспалительный процесс и гистопатологические изменения при ряде воспалительных и аутоиммунных заболеваний нервной системы, артрите, гломерулонефрите, системной красной волчанке и других болезнях во многих случаях обусловлены активацией системы комплемента крови, представляющей собой еще одну из систем неспецифического иммунитета. Комплемент играет важную роль и в реакции отторжения трансплантата.
Поксвирусы кодируют особый белок (КСБ), способный ингибировать активацию комплемента, причем наиболее эффективно с белками комплемента человека взаимодействует белок того же вируса натуральной оспы. Сегодня считается, что именно этот вирусный белок может стать перспективным препаратом для лечения болезни Альцгеймера, синдрома мультиорганной дисфункции и отторжения ксенотрансплантатов (Jha, Kotwal, 2003). Так, на лабораторных животных было показано, что рекомбинантный КСБ способствует восстановлению функций мозга после средней и тяжелой черепно-мозговой травмы. Этот белок оказался эффективен и при травмах спинного мозга: при его применении значительно уменьшались гистопатологические изменения, вызванные воспалительными реакциями.
В патогенезе воспалительных и аутоиммунных заболеваний важную роль играют и хемокины – обширное семейство небольших белков, имеющих очень сходную третичную структуру. Поксвирусы кодируют хемокинсвязывающие белки, по аминокислотной последовательности не имеющие гомологов среди известных белков позвоночных. Их высокий терапевтический потенциал был показан на ряде лабораторных моделей воспалительных и аутоиммунных заболеваний (Непомнящих, Щелкунов, 2008).
Таким образом, предположение ученых, что вирусные белки, являющиеся антагонистами иммунных белков-регуляторов, можно использовать в терапевтических целях, полностью подтвердилось. Сегодня на их основе с помощью методов генетической инженерии уже разрабатываются препараты нового поколения для коррекции патологических состояний человека, связанных с избыточной активацией белков-медиаторов защитных систем организма.
И с этой точки зрения, мы можем по-новому взглянуть на смертельно опасный для человека вирус натуральной оспы: результаты лабораторных исследований и доклинических испытаний свидетельствуют о большой перспективности препаратов для лечения тяжелейших воспалительных и аутоиммунных заболеваний человека, созданных на основе белков этого вируса, на протяжении столетий бывшего настоящим бичом цивилизаций.
Непомнящих Т. С., Щелкунов С. Н. Иммуномодулирующие белки поксвирусов как новые средства иммунокорректирующей терапии // Молекуляр. биология. 2008. Т. 42, №. 5. С. 904–912.
Щелкунов С. Н. Вирус натуральной оспы – источник новых медицинских препаратов // Соросовский образовательный журнал. 1995. № 1. С. 28—31.
Щелкунов С. Н. Генетическая инженерия: Учеб.-справ. пособие. 3-е изд., испр. и доп. Новосибирск: Сиб. универ. изд-во, 2008. 514 с.
Щелкунов С. Н. Преодоление ортопоксвирусами защитных систем организма млекопитающих // Молекуляр. биология. 2011. Т. 45, № 1. С. 30—43.
Щелкунов С. Н. Оспа – дамоклов меч цивилизаций // Наука из первых рук. 2012. № 6 (48). С. 96—109.
Shchelkunov S. N. Orthopoxvirus genes that mediate disease virulence and host tropism // Advances in Virology. 2012. Vol. 2012, Article ID 524743, 17 p. doi:10.1155/2012/524743.
Работа выполнена при финансовой поддержке РФФИ (грант 12-04-00110а)
Биологическим оружием (БО) называют боевые средства, поражающее действие которых основано на использовании болезнетворных свойств микроорганизмов (возбудителей) или микробов, вызывающих болезни людей, животных и растений. Цель применения биологического оружия — снижение боеспособности противника. Это может быть достигнуто путём непосредственного поражения людей, а также уничтожением животных и сельскохозяйственных растений, в результате чего человек лишается средств существования (продовольствия), а в некоторых случаях порчи материалов вооружения, военной техники и снаряжения.
Биологическое оружие обладает рядом особенностей, главная из которых способность вызывать массовые заболевания людей (эпидемии), животных (эпизоотии) и растении (эпифитотии). Для заражения достаточно незначительного числа микробов. Попав в организм, микробы быстро размножаются, вызывают его заболевание, а затем вследствие контакта людей друг с другом, через выделения больных, воздух, воду, пищевые продукты, а также через различных переносчиков, обычно насекомых, заболевание при благоприятных условиях может приобрести весьма широкие масштабы.
При этом могут использоваться микробы (вирусы, бактерии, грибки) – возбудители бруцеллеза, туляремии, сибирской язвы, чумы, холеры, сапа, дифтерии, брюшного тифа, лихорадки, энцефалита, оспы, гриппа и многих других заболеваний.
Поражающее действие БО проявляется не сразу, а спустя определенное время (инкубационный период), зависящее как от вида и количества попавших в организм болезнетворных микробов или их токсинов, так и от физического состояния организма. Наиболее часто инкубационный период продолжается от 2 до 5 суток. В течение почти всего этого периода личный состав сохраняет боеспособность, иногда даже не подозревая о состоявшемся заражении. Некоторые из возникающих в результате заражения заболевания, называемые контагиозными (чума, натуральная оспа и др.), могут затем передаваться от пораженных к окружающим здоровым людям через воздух, укусы кровососущих насекомых и другими путями. Заболевания, называемые неконтагиозными (сибирская язва, туляремия и др.), от больных людей к здоровым практически не передаются. Классификация заболеваний приведена на рис.5.

Рис. 5. Классификация заболеваний
Особо следует подчеркнуть сильное психологическое воздействие, оказываемое БО на человека. Наличие реальной угрозы внезапного применения противником БО, как и появление в войсках и среди гражданского населения крупных вспышек и эпидемий опасных инфекционных заболеваний, способны повсеместно вызвать страх, панические настроения, снизить боеспособность войск, дезорганизовать работу тыла.
Основу поражающего действия биологического оружия составляют биологические средства (БС) – специально отобранные для боевого применения биологические агенты, способные в случае проникновения в организм людей, животных (растений) вызывать тяжелые инфекционные заболевания. К ним относят: отдельные виды болезнетворных микробов и вирусов – возбудителей наиболее опасных инфекционных заболеваний, а также токсичные продукты их жизнедеятельности; генетический материал – молекулы инфекционных нуклеиновых кислот, полученный из микробов (вирусов). Для уничтожения посевов зерновых, технических и других сельскохозяйственных культур можно ожидать помимо использования микробов – возбудителей болезней культурных растений преднамеренное применение насекомых – наиболее опасных вредителей сельскохозяйственных культур.
Патогенные микроорганизмы – возбудители инфекционных болезней чрезвычайно малы по размерам, не имеют цвета, запаха, вкуса и поэтому не определяются органами чувств человека. В зависимости от размеров, строения и биологических свойств они подразделяются на классы (рис.6), из которых помимо вирусов наибольшее значение имеют бактерии, риккетсии и грибки.

Рис.6. Классификация биологических агентов
Риккетсии – это небольшие (размером от 0.4 до 1 мкм) клетки-палочки. Размножаются поперечным бинарным делением только внутри клеток живых тканей. Они не образуют спор, но достаточно устойчивы к высушиванию, замораживанию действию относительно высоких температур (до 5600С). Риккетсии являются причиной таких тяжелых заболеваний человека, как сыпной тиф, пятнистая лихорадка Скалистых гор и др.
Грибки – одно- или многоклеточные микроорганизмы растительного происхождения, отличающиеся от бактерий более сложным строением и способом размножения. Споры грибков высокоустойчивы к высушиванию, воздействию солнечных лучей и дезинфицирующих веществ. Заболевания, вызываемые патогенными грибками, характеризуются поражением внутренних органов с тяжелым и длительным течением.
Вирусы – обширная группа биологических агентов, не имеющих клеточной структуры, способных развиваться и размножаться только в живых клетках, используя для этого их биосинтетический аппарат. Размеры внеклеточных форм вирусов колеблются от 0,02 до 0,4 мкм. Большинство из них недостаточно устойчивы к различным факторам внешней среды: плохо переносят высушивания, солнечный свет, особенно ультрафиолетовые лучи, а также температуру 6000С и действия дезинфицирующих средств. Патогенные вирусы являются причиной многих тяжелых заболеваний человека, таких как натуральная оспа, тропические геморрагические лихорадки, ящур и др.
Эффективность действия БО зависит не только от поражающих способностей биологических средств, но в значительной степени от правильно выбора способов и средств их применения.
Способы боевого применения БС основываются на способности патогенных микробов в естественных условиях проникаться в организм человека следующими путями:
- с воздухом через органы дыхания (аэрогенный, воздушно-капельный путь);
- с пищей и водой через пищеварительный тракт (алиментарный путь);
- через неповрежденную кожу в результате укусов зараженных кровососущих членистоногих (трансмиссивный путь);
- через слизистые оболочки рта, носа, глаза, а также через поврежденные кожные покровы (контактный путь).
Способы боевого применения БС:
- распыление биологических рецептур для заражения приземного слоя воздуха частицами аэрозоля – аэрозольный способ;
- рассеивание в районе цели искусственно зараженных биологическими средствами кровососущих переносчиков – трансмиссионный способ;
- заражение биологическими средствами воздуха и воды в замкнутых пространствах (объема) при помощи диверсионного снаряжения – диверсионный способ.
Аэрозольный способ является основным способом боевого применения БС. Он позволяет внезапно и скрытно заражать биологическими средствами на больших пространствах приземные массы воздуха, местность и находящиеся на ней живую силу, вооружение и военную технику. При этом заражению биологическим аэрозолем одновременно подвергается живая сила, не только открыто расположенная на местности, но и находящаяся в негерметизированных вооружении, военной технике и сооружениях.
Перевод биологических рецептур в аэрозоль осуществляется двумя основными методами: силой взрыва ВВ биологического боеприпаса и с помощью распылительных устройств.
К достоинствам первого метода (взрыва) относят простоту, надежность, высокую экономичность. Однако в результате образования в момент взрыва высокой температуры и ударной волны наблюдается значительная гибель биологических средств.
В распылительных устройствах перевод рецептуры в аэрозоль осуществляется либо под воздействием сжатого инертного газа (в механических генераторах аэрозолей), либо набегающим воздушным потоком (в выливных авиационных приборах). Распылительные устройства, устанавливаемые на пилотируемых и беспилотных летательных аппаратах, позволяют создавать на определенных высотах облако зараженной атмосферы, которое, дрейфуя и постепенно оседая, способно заражать приземные воздушные массы над значительной по площади территорией.
Трансмиссионный способ заключается в преднамеренном рассеивании в заданном районе искусственно зараженных биологическими средствами кровососущих переносчик с помощью энтомологических боеприпасов (авиационных бомб и контейнеров специальной конструкции).
Трансмиссионный способ основан на том, что многие из существующих в природе кровососущих членистоногих легко воспринимают, длительной сохраняют, а затем через укусы передают возбудителей ряда опасных для человека и животных заболеваний. Так, отдельные виды комаров способны передавать желтую лихорадку, лихорадку денге, Венесуэльский энцефаломиелит лошадей, блохи – чуму, вши – сыпной тиф, москиты – лихорадку паппатачи.
Применение искусственно зараженных переносчиков наиболее вероятно в теплое время года и природных условиях, близких к естественному обитанию переносчиков.
Диверсионный способ применения БС заключается в преднамеренном скрытном заражении биологическими средствами замкнутых пространств (объектов) воздуха и воды, а также продовольствия (фуража), используемых непосредственно, без дополнительной очистки (обработки).
С помощью малогабаритного диверсионного снаряжения (портативных генераторов аэрозолей, распыляющих пеналов и т.п.) возможно в определенный момент осуществить заражение воздуха в местах массового скопления людей. Возможно также заражение воды в городских водонапорных системах, для чего могут быть использованы возбудители чумы, холеры, брюшного тифа и особенно ботулинический токсин. Путем диверсий, кроме того, могут быть распространены искусственно зараженные кровососущие переносчики и насекомые.
Основным методом применения биологических рецептур является распыление их в воздухе и создание, таким образом, облака биологического аэрозоля. Заболевания личного состава будут при этом возникать в результате вдыхания аэрозольных частиц, содержащих возбудителей болезней. Таким путём можно заразиться многими заболеваниями, включая и такие, возбудители которых в естественных условиях воздушно капельным путём не передаются. БО способно вызывать поражения на больших площадях, чем другие средства поражения. Это связано с высокой инфекционностью биологических аэрозолей. Непосредственная защита личного состава в период биологического нападения противника обеспечивается использованием средств индивидуальной и коллективной защиты, а также применением средств экстренной профилактики, имеющихся в индивидуальных аптечках.
Личный состав, находящийся в очаге биологического заражения, должен не только своевременно и правильно использовать средства защиты, но и строго выполнять правила личной гигиены и следующие правила:

У РНК-содержащих ретровирусов сначала происходит обратная транскрипция генома в ДНК, затем ее интеграция в клеточные хромосомы и лишь после этого транскрипция генов.
Цитопатические эффекты при вирусных инфекциях разнообразны, они определяются как вирусом, так и клеткой и сводятся к разрушению клетки (цитолитический эффект), сосуществованию вируса и клетки без гибели последней (латентная и персистирующая инфекция) и трансформации клетки.
Вовлеченность организма в инфекционный процесс зависит от ряда обстоятельств - количества погибших клеток, токсичности вирусов и продуктов распада клеток, от реакций организма, начиная от рефлекторных и заканчивая иммунными. Количество погибших клеток влияет на тяжесть инфекционного процесса. Например, будут ли поражены при гриппе только клетки носа и трахеи или вирус поразит клетки эпителия альвеол, зависит тяжесть и исход болезни.
Хотя вирусы и не образуют типичных токсинов, однако и вирионы, и вирусные компоненты, накапливающиеся в пораженных тканях, выходя в кровоток, оказывают токсическое действие. Неменьшее токсическое действие оказывают и продукты распада клеток. В этом случае действие вирусной инфекции столь же неспецифично, как и действие патогенных организмов, убивающих клетки и вызывающих их аутолиз. Поступление токсинов в кровь вызывает ответную реакцию - лихорадку, воспаление, иммунный ответ. Лихорадка является преимущественно рефлекторным ответом на поступление в кровь и воздействие на ЦНС токсичных веществ.
Если лихорадка - общий ответ организма на вирусную инфекцию, то воспаление - это местная многокомпонентная реакция. При воспалении происходят инфильтрация пораженных тканей макрофагами, утилизация продуктов распада, репарация и регенерация. Одновременно развиваются реакции клеточного и гуморального иммунитета. На ранних стадиях инфекции действуют неспецифические киллеры и антитела класса IgM. Затем вступают в действие основные факторы гуморального и клеточного иммунитета. Однако гораздо раньше, уже в первые часы после заражения, начинает действовать система интерферона, представляющая семейство секреторных белков, вырабатываемых клетками организма в ответ на вирусы и другие стимулы. Описанные явления относятся к так называемой острой репродуктивной вирусной инфекции. Взаимодействие вируса и клеток может происходить, как отмечалось выше, без гибели последних. В этом случае говорят о латентной, т.е. бессимптомной или персистирующей хронической вирусной инфекции. Дальнейшая экспрессия вируса, образование вирусспецифических белков и вирионов вызывает синтез антител, на этой стадии латентная инфекция переходит в персистирующую и появляются первые признаки болезни.
Репродукция вируса в клетках сопровождается развитием цитопатических процессов, специфичных для разных вирусов и для разных типов инфекционных процессов. Цитопатические процессы при вирусных инфекциях разнообразны, они определяются как вирусом, так и клетками, причем специфика их больше "задается" клеткой, нежели вирусом, и сводится в основном к разрушению клеток, сосуществованию вируса и клеток без гибели последних и трансформация клеток. Несмотря на значительные различия цитоцидного действия разных вирусов, в общем, они сходны. Подавление синтеза клеточных макромолекул - нуклеиновых кислот и белков, а также истощение энергетических ресурсов клетки ведут к необратимым процессам, заканчивающимся гибелью пораженной клетки. Повреждение клеток вирусами, их отмирание и распад переносят вирусную инфекцию с клеточного уровня на уровень организма в целом.
При встрече организма с вирусной инфекцией продукция интерферона (растворимого фактора, вырабатываемого вирус-инфицированными клетками, способного индуцировать антивирусный статус в неинфицированных клетках) становится наиболее быстрой реакцией на заражение, формируя защитный барьер на пути вирусов намного раньше специфических защитных реакций иммунитета, стимулируя клеточную резистентность, - делая клетки непригодными для размножения вирусов.
Продукция и секреция цитокинов относятся к самым ранним событиям, сопутствующим взаимодействию микроорганизмов с макрофагами. Этот ранний неспецифический ответ на инфекцию важен по нескольким причинам: он развивается очень быстро, поскольку не связан с необходимостью накопления клона клеток, отвечающих на конкретный антиген; ранний цитокиновый ответ влияет на последующий специфический иммунный ответ.
Интерферон активирует макрофаги, которые затем синтезируют интерферон-гамма, ИЛ-1, 2, 4, 6, ФНО, в результате макрофаги приобретают способность лизировать вирус-инфицированные клетки.
Интерферон-гамма является специализированным индуктором активации макрофагов, который способен индуцировать экспрессию более 100 разных генов в геноме макрофага.
Продуцентами этой молекулы являются активированные Т-лимфоциты (Тh1-тип) и естественные киллеры (NK-клетки). Интерферон-гамма индуцирует и стимулирует продукцию провоспалительных цитокинов (ФНО, ИЛ-1, 6), экспрессию на мембранах макрофагов, антигенов МНС II; гамма-интерферон резко усиливает антимикробную и противовоспалительную активность путем повышения продукции клетками супероксидных радикалов, а усиление иммунного фагоцитоза и антителоопосредованной цитотоксичности макрофагов под влиянием гамма-интерферона связано с усилением экспрессии Fc-рецепторов для JgG. Активирующее действие интерферона-гамма на макрофаги опосредовано индукцией секреции этими клетками ФНО -альфа. Этот пик наблюдается совместно с ФНО-альфа. Максимум продукции ИЛ-4 наступает через 24-48 ч с момента активации клеток. При этом ИЛ-4 рассматривается как цитокин, ограничивающий иммуновоспалительные реакции и снижающий ответ организма на инфекцию, угнетая при этом экспрессию гамма-интерферона. Интерферон-гамма ин витро усиливает фагоцитарную активность нейтрофилов, что обусловлено усилением экспрессии Fc-рецепторов и поверхностных белков семейства интегринов на нейтрофилы. Это позволяет нейтрофилам осуществлять цитотоксические функции и фагоцитоз. В качестве основных эффекторных клеток воспалительного процесса, они обеспечивают элиминацию инфекта из организма.
Взаимодействие цитокина с клеткой определяется универсальной биологической системой, специфическим механизмом которой является рецепторный аппарат, связанный с восприятием метаболического кода. Для проявления биологической активности цитокина необходимо присутствие на поверхности чувствительных клеток специфических рецепторов, которые могут экспрессироваться параллельно с синтезом цитокина. Рецепторы цитокинов представляют собой комплексы, состоящие из двух и более рецепторных молекул, которые объединяются на мембране клетки-мишени и образуют высокоаффинный рецепторный комплекс. Большинство рецепторов состоит из отдельных молекул, связывающих цитокины, которые ассоциируются после связывания лиганда с сигналпередающим рецепторным компонентом; часть рецепторов существует как растворимые изоформы, способные связывать и растворять цитокины, а часть функционирует как многокомпонентные блоки; механизм комплексирования субъединиц рецепторов объясняет плейотропные и дублирующие эффекты цитокинов, имеющих большое структурное сходство. Рецепторы ИЛ-10 имеют гомологию рецепторов интерферона, и подобно ИЛ-10 индуцирует экспрессию в моноцитах гена Fc- рецептора. Для полного функционирования цитокиновой системы необходимы повышение уровня цитокина в ответ на инфект и экспрессия нормального количества рецепторов к ним на клетках. Изменение рецепторов после их связывания с цитокином заключается в интернализации комплексов цитокин - рецептор внутрь клетки. На поверхности клеток рецептор появляется заново, постепенно синтезируясь в течение 24-36 ч (время появления рецепторов интерферон-альфа). В этот период клетки остаются чувствительными к последующим дозам цитокина, чем объясняется эффективность введения препаратов интерферона и их индукторов три раза в неделю.
При антигенной стимуляции клеток трансдукция сигнала с активированного рецептора на генетический аппарат осуществляется с помощью внутриклеточных регуляторных систем, компоненты которых (белки мембран, ферментов, хроматина) связываются с чувствительными к ним последовательностями ДНК. После связывания цитокина (интерферон) с поверхностными клеточными мембранными рецепторами происходит активация ферментов протеинкиназы-С (ПКС), тирозинкиназы, ц-АМФзависимой протеинкиназы, серин-треонинкиназы. Интерферон-альфа активирует tyk 2 и jak 1-киназы, а интерферон-гамма активирует jak 1 и 2-киназы. Далее факторы транскрипции перемещаются в ядро клетки и связывают гены раннего ответа.
Первый ответ клеток на цитокин - это быстрая индукция генов раннего ответа ("immediate early" генов), в число которых и входит ген интерферон-гамма. Стимуляция экспрессии этих генов важна для выхода клеток из Go-стадии и перехода в Gi-стадию и дальнейшей прогрессии клеточного цикла. Их индукция происходит после активации рецепторов роста на клеточной мембране и активации протеин-киназной системы. Гены раннего ответа являются ключевыми регуляторами клеточной пролиферации и дифференцировки, кодируют белки, регулирующие репликацию ДНК.
Таким образом, при активации клеток происходит стимуляция генов раннего ответа, что ассоциируется с изменением фаз клеточного цикла. Основная протективная роль в иммунном ответе, направленном против внутриклеточных паразитов (грибы, простейшие, вирусы, микобактерии туберкулеза), принадлежит клеточным механизмам. Способность перечисленных возбудителей переживать и размножаться внутри клеток делает их защищенными от действия антител и системы комплемента. Резистентность к антимикробным факторам макрофагов позволяет им длительно переживать внутри этих клеток. Для элиминации возбудителя необходим специфический клеточно-опосредованный ответ. Его специфичность определяется антигенраспознающими СД8+-Т-лимфоцитами, которые пролиферируют, активируются и формируют клон эффекторных цитотоксических лимфоцитов. Решающий момент специфического иммунного ответа - это ответ СД4+Т-лимфоцитов с хелперной направленностью на распознавание антигена. На этом этапе определяется форма иммунного ответа: либо с преобладанием гуморального иммунитета, либо с преобладанием клеточных реакций (ГЗТ). Направление дифференцировки СД4 + -лимфоцитов, от которого зависит форма специфического иммунного ответа, контролируется цитокинами, образующимися в ходе воспалительной реакции. Так, в присутствии ИЛ-12 и интерферон-гамма СД4 + -лимфоциты дифференцируются в воспалительные Тh1-клетки, начинают продуцировать и секретировать интерлейкин-2, интерферон-гамма, ФНО и определяют клеточный характер специфического иммунного ответа. Присутствие ИЛ-12 обеспечивается его продукцией макрофагами, а интерферон-гамма - естественными киллерами, активированными в раннюю фазу ответа на внутриклеточно паразитирующие бактерии и вирусы. В отличие от этого, в присутствии ИЛ-4 СД4 + -лимфоциты дифференцируются в хелперы Тh 2, которые начинают продуцировать и секретировать ИЛ-4, ИЛ-5, ИЛ-6 и запускают гуморальный иммунный ответ, т.е. синтез специфических антител - иммуноглобулинов. Воспалительные Тh 1-лимфоциты нужны для борьбы с внутриклеточными паразитами, а Тh 2 хелперы нужны для элективной защиты от внеклеточных паразитов.
Вирусная инфекция может вызывать быстрое подавление экспрессии ряда клеточных генов (из которых наиболее изучены интерфероновые гены и гены, кодирующие дс-РНК-зависимые ферменты -2,5-ОАС и ПК-дс), принимающих участие в антивирусном действии. Специальные исследования механизма антивирусного действия интерферонов и дс-РНК в клеточных и бесклеточных системах показали ключевую роль в этом процессе вышеуказанных ферментов. ПК-дс, взаимодействуя с дс-РНК, фосфорилируется и в активной форме фосфорилирует регуляторные факторы транскрипции и трансляции, из которых наиболее изучен инициирующий фактор трансляции (eIF2).
ПК-дс выполняет регуляторную роль в системе клеточной пролиферации на уровне факторов трансляции и активации ряда генов цитокинов. Вероятно, существует связь между подавлением транскрипции мРНК и ПК-дс, угнетением общего синтеза клеточного белка при вирусных инфекциях и накоплением в ядрах клеток белка нуклеокапсида и белка NSP2. Фрагментация клеточных хромосом, наблюдающаяся на ранних сроках вирусной инфекции, может быть одной из причин подавления экспрессии генов, участвующих в противовирусном ответе.
Есть основания предполагать участие белков NSP2 в регуляции активности генов цитокинов - низкомолекулярных белковых регуляторных веществ, продуцируемых клетками и способных модулировать их функциональную активность. Нарушения в системе цитокинов приводят к нарушению кооперативных взаимодействий иммунокомпетентных клеток и нарушению иммунного гомеостаза.
В последние годы показано, что ИЛ- 12, относящийся к провоспалительным цитокинам, является ключевым для усиления клеточно-опосредованного иммунного ответа и инициации эффективной защиты против вирусов.
Средства терапии гриппа и ОРЗ можно разделить на этиотропные, иммунокорригирующие, патогенетические и симптоматические. Приоритет принадлежит этиотропным препаратам, действие которых направлено непосредственно на возбудитель инфекции. Все препараты этиотропного действия целесообразно рассматривать с учетом их точек приложения в цикле репродукции вирусов гриппа и других ОРЗ.
Применение химиопрепаратов для профилактики и лечения гриппа и ОРЗ относится к базовой терапии и является общепризнанным мировым стандартом. Многолетние клинические исследования достоверно выявили их высокую лечебно-профилактическую значимость. Химиотерапевтические средства представлены тремя основными группами: это блокаторы М2-каналов (амантадин, ремантадин); ингибиторы нейраминидазы (занамивир, озельтамивир) и ингибиторы протеаз (амбен, аминокапроновая кислота, трасилол). Препараты оказывают прямое антивирусное действие, нарушая различные фазы репликативного цикла вирусов. Несколько особняком стоит группа вирулицидных препаратов, применяемых местно для предотвращения адсорбции и проникновения вирионов в клетки.
Бактериофаги – это вирусы, которые поражают только бактерий. В ходе инфекции они влияют на все процессы жизнедеятельности бактериальной клетки, фактически превращая ее в фабрику по производству вирусного потомства. В конце концов клетка разрушается, а вновь образованные вирусные частицы выходят наружу и могут заражать новые бактерии.
Несмотря на огромное число и разнообразие природных фагов, встречаемся мы с ними редко. Однако бывают ситуации, когда деятельность этих вирусов не остается незамеченной. Например, на предприятиях, где производят сыры, йогурты и другие молочно-кислые продукты, часто приходится сталкиваться с вирусной атакой на бактерии, сбраживающие молоко. В большинстве таких случаев фаговая инфекция распространяется молниеносно, и полезные бактерии гибнут, что приводит к значительным экономическим потерям (Neve et al., 1994).
Именно благодаря прикладным исследованиям в интересах молочной промышленности, направленным на получение устойчивых к бактериофагам штаммов молочно-кислых бактерий, был открыт ряд механизмов, с помощью которых бактерии избегают инфекции. Параллельно были изучены способы, с помощью которых вирусы, в свою очередь, преодолевают бактериальные системы защиты (Moineau et al., 1993).
Кто защищен – тот вооружен
На сегодня известно пять основных, весьма хитроумных механизмов защиты, которые бактерии выработали в непрестанной борьбе с вирусами: изменение рецептора на поверхности клетки; исключение суперинфекции; системы абортивной инфекции; системы рестрикции-модификации и, наконец, системы CRISPR-Cas.
К средствам противовирусной защиты бактерий относятся и системы рестрикции-модификации, в которые входят гены, кодирующие два белка-фермента – рестриктазу и метилазу. Рестриктаза узнает определенные последовательности ДНК длиной 4—6 нуклеотидов и вносит в них двуцепочечные разрывы. Метилаза, напротив, ковалентно модифицирует эти последовательности, добавляя к отдельным нуклеотидным основаниям метильные группы, что предотвращает их узнавание рестриктазой.
Врага нужно знать в лицо
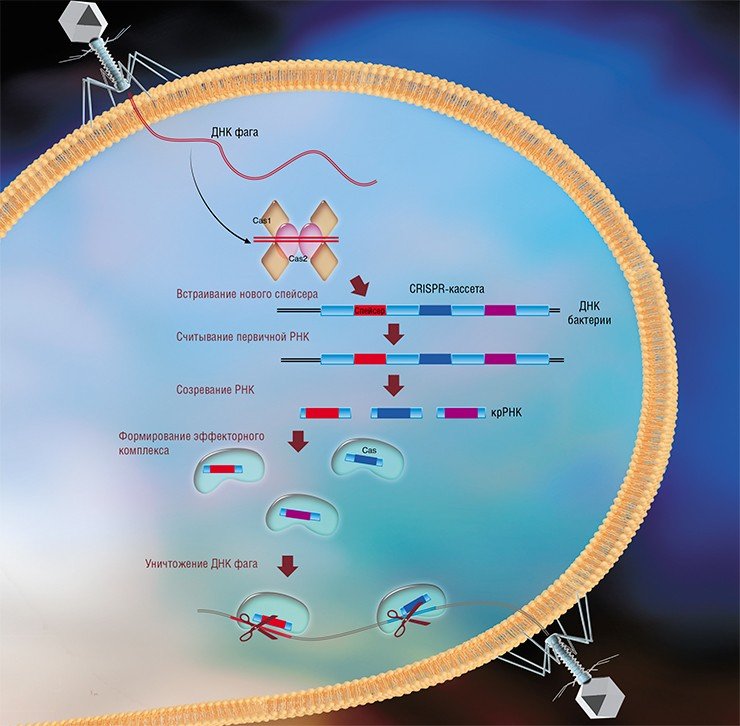
Системы CRISPR-Cas являются уникальным примером адаптивного иммунитета бактерий. При проникновении в клетку ДНК фага специальные белки Cas встраивают фрагменты вирусной ДНК длиной 25—40 нуклеотидов в определенный участок генома бактерии (Barrangou et al., 2007). Такие фрагменты называются спейсерами (от англ. spacer – промежуток), участок, где происходит встраивание, – CRISPR-кассета (от англ. Clustered Regularly Interspaced Short Palindromic Repeats), а сам процесс приобретения спейсеров – адаптацией.
Чтобы использовать спейсеры в борьбе с фаговой инфекцией, в клетке должен происходить еще один процесс, управляемый белками Cas, названный интерференцией. Суть его в том, что в ходе транскрипции CRISPR-кассеты образуется длинная молекула РНК, которая разрезается белками Cas на короткие фрагменты – защитные криспрРНК (крРНК), каждая из которых содержит один спейсер. Белки Cas вместе с молекулой крРНК образуют эффекторный комплекс, который сканирует всю ДНК клетки на наличие последовательностей, идентичных спейсеру (протоспейсеров). Найденные протоспейсеры расщепляются белками Cas (Westra et al., 2012; Jinek et al., 2012).
Системы CRISPR-Cas обнаружены у большинства прокариот – бактерий и архей. Хотя общий принцип действия всех известных систем CRISPR-Cas одинаков, механизмы их работы могут существенно отличаться в деталях. Наибольшие различия проявляются в строении и функционировании эффекторного комплекса, в связи с чем системы CRISPR-Cas делят на несколько типов. На сегодняшний день описаны шесть типов таких неродственных друг другу систем (Makarova et al., 2015; Shmakov et al., 2015).
Наиболее изученной является система CRISPR-Cas I типа, которой обладает излюбленный объект молекулярно-биологических исследований – бактерия кишечная палочка (Esсherichia coli). Эффекторный комплекс в этой системе состоит из нескольких небольших белков Cas, каждый из которых отвечает за разные функции: разрезание длинной некодирующей CRISPR РНК, связывание коротких крРНК, поиск, а затем разрезание ДНК-мишени.
Гонка вооружений
Бактериофаги, как факторы среды, вызывают направленные изменения в геноме бактерий, которые наследуются и дают бактериям явное преимущество, спасая от повторных инфекций. Поэтому системы CRISPR-Cas можно считать примером ламарковской эволюции, при которой происходит наследование благоприобретенных признаков (Koonin et al., 2009)
Некоторые бактериофаги реагируют на наличие в бактериальной клетке систем CRISPR-Cas выработкой особых анти CRISPR-белков, способных связываться с белками Cas и блокировать их функции (Bondy-Denomy et al., 2015). Еще одно ухищрение — обмен участков генома вируса, на которые нацелена система CRISPR-Cas, на участки геномов родственных вирусов, отличающихся по составу нуклеотидной последовательности (Paez-Espino et al., 2015).
Благодаря постоянному совершенствованию биоинформатических алгоритмов поиска, а также включению в анализ все большего количества прокариотических геномов, открытие новых типов CRISPR-Cas систем является делом недалекого будущего. Предстоит также выяснить и детальные механизмы работы многих недавно открытых систем. Так, в статье, опубликованной в 2016 г. в журнале Science и посвященной анализу системы CRISPR-Cas VI типа, описан белок С2с2, образующий эффекторный комплекс с крРНК, который нацелен на деградацию не ДНК, а РНК (Abudayyeh et al., 2016). В будущем такое необычное свойство может быть использовано в медицине для регулирования активности генов путем изменения количества кодируемых ими РНК.
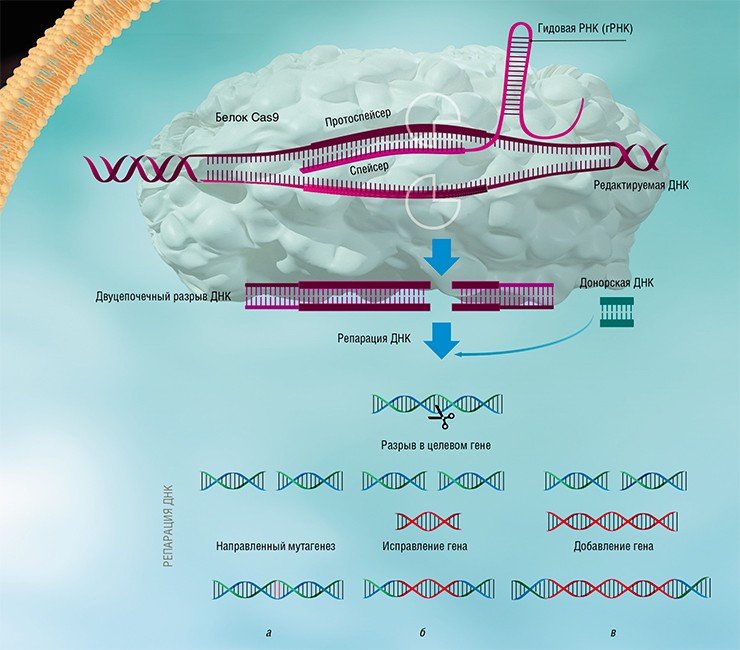
Изучение стратегий борьбы бактерий с бактериофагами, несмотря на свою кажущуюся фундаментальность и отвлеченность от задач практической медицины, принесло неоценимую пользу человечеству. Примерами этого могут служить методы молекулярного клонирования и редактирования геномов – направленного внесения или удаления мутаций и изменения уровня транскрипции определенных генов.
Благодаря быстрому развитию методов молекулярной биологии всего лишь через несколько лет после открытия механизма действия систем CRISPR-Cas была создана работающая технология геномного редактирования, способная бороться с болезнями, ранее считавшимися неизлечимыми. Доступность и простота этой технологии позволяют рассматривать ее как основу для медицины, ветеринарии, сельского хозяйства и биотехнологий будущего, которые будут базироваться на направленных и безопасных генных модификациях.
Нет никаких сомнений, что дальнейшее изучение взаимодействия бактерий и их вирусов может открыть перед нами такие возможности, о которых мы сейчас даже не подозреваем.
Abudayyeh O. O., Gootenberg J. S., Konermann S. et al. C 2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector // Science. 2016. V. 353: aaf5573.
Barrangou R., Fremaux C., Deveau H. et al. CRISPR provides acquired resistance against viruses in prokaryotes // Science. 2007. V. 315. P. 1709–1712.
Bikard D., Marraffini L. A. Innate and adaptive immunity in bacteria: mechanisms of programmed genetic variation to fight bacteriophages // Curr. Opin. Immunol. 2012. V. 1 P. 15–20.
Bondy-Denomy J., Garcia B., Strum S. et al. Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins // Nature. 2015. V. 526. P. 136–139.
Calendar R., Abedon S. T. The Bacteriophages // 2nd Ed., Oxford University Press. 2006.
Datsenko K. A., Pougach K., Tikhonov A. et al. Molecular memory of prior infections activates the CRISPR/Cas adaptive bacterial immunity system // Nat. Commun. 2012. V. 3. P. 945
Jiang W., Marraffini L. A. CRISPR-Cas: New Tools for Genetic Manipulations from Bacterial Immunity Systems // Annu. Rev. Microbiol. 2015. V. 69. P. 209–28.
Jinek M., Chylinski K., Fonfara I., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity // Science. 2012. V. 337. P. 816–821.
Koonin E. V., Wolf Y. I. Is evolution Darwinian or/and Lamarckian? // Biol. Direct. 2009. V. 4. P. 42.
Lopez-Pascua L., Buckling A. Increasing productivity accelerates host-parasite coevolution // J. Evol. Biol. 2008. V. 3. P. 853–860.
Makarova K. S., Wolf Y. I., et al. An updated evolutionary classification of CRISPR-Cas systems // Nat. Rev. Microbiol. 2015. V. 11. P. 722–736.
Moineau, S., Pandian S., Klaenhammer T. R. Restriction/modification systems and restriction endonucleases are more effective on lactococcal bacteriophages that have emerged recently in the dairy industry // Appl. Envir. Microbiol. 1993. V. 59. P. 197–202.
Neve H., Kemper U., et al. Monitoring and characterization of lactococcal bacteriophage in a dairy plant // Kiel. Milckwirtsch. Forschungsber. 1994. V. 46. P. 167–178.
Nuñez J. K., Harrington L. B., et al. Foreign DNA capture during CRISPR-Cas adaptive immunity // Nature. 2015a. V. 527. P. 535–538.
Nuñez J. K., Kranzusch P. J., et al. Cas1-Cas2 complex formation mediates spacer acquisition during CRISPR-Cas adaptive immunity // Nat. Struct. Mol. Biol. 2014. V. 21. P. 528–534.
Nuñez J. K., Lee A. S., Engelman A., Doudna J. A. Integrase-mediated spacer acquisition during CRISPR-Cas adaptive immunity // Nature. 2015b. V. 519. P. 193–198.
Paez-Espino D., Sharon I., et al. CRISPR Immunity Drives Rapid Phage Genome Evolution in Streptococcus thermophilus // MBio. 2015. V. 6: e00262–15.
Shmakov S., Abudayyeh O. O., Makarova K. S., et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. // Mol. Cell. 2015. V. 60. P. 385–397
Tan D., Svenningsen S. L., Middelboe M. Quorum sensing determines the choice of antiphage defense strategy in Vibrio anguillarum. // mBio 2015. V. 6: e00627–15.
Westra E. R., van Erp P. B., Künne T., et al. CRISPR immunity relies on the consecutive binding and degradation of negatively supercoiled invader DNA by Cascade and Cas3 // Mol. Cell. 2012. V. 46. P. 595–605.
Читайте также: