
Днк содержащие вирусов классификация днк-содержащих
Обновлено: 24.04.2024
ДНК-вирусы как причина развития рака - вирусный канцерогенез
В канцерогенезе также могут участвовать ДНК-содержащие вирусы. Наиболее крупные ДНК-содержащие вирусы принадлежат к группе герпесвирусов. Первым вирусом, выделенным из культуры опухолевых клеток, был вирус Эпштейна-Баррра (ЭБВ).
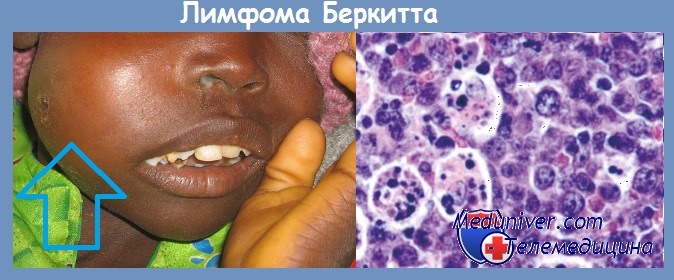
Вирус был получен из культуры опухолевых клеток больных лимфомой Беркита. В культуре in vitro ЭБВ вызывает бесконечную пролиферацию человеческих В-лимфоцитов. Вирусные гены, встраиваясь в геном этих клеток, инициируют пролиферацию путем активации клеточных генов-регуляторов роста (онкогенов).
В крови больных лимфомой Беркита обнаруживается более высокий, чем у здоровых людей, титр антител к антигенам вирусной оболочки и к мембранным антигенам. В то же время в африканских странах среди множества детей, инфицированных этим вирусом, лимфома развивается только в единичных случаях.
Механизм онкогенеза этого заболевания не может быть объяснен только вирусным заражением. К тому же, за пределами африканского континента регистрируются случаи заболевания лимфомой Беркита, абсолютно не связанные с инфицированием вирусом ЭБВ. По-видимому, ЭБВ также может принимать участие в патогенезе карциномы носоглотки, так как клетки этой опухоли экспрессируют вирусные антигены EBNA-1.
При исследованиях саркомы Капоши, развивающейся у больных СПИДом, в 1994 г. из ее клеток был выделен новый герпесвирус. Первоначально вирус получил название герпес-вируса саркомы Капоши (ГСК), впоследствии он был переименован в герпесвирус человека серотип 8 (ГВЧ 8). Вирус обнаруживается в веретеновидных клетках саркомы Капоши практически при всех случаях заболевания.
ГВЧ 8 был также найден в опухолевых клетках первичной эффузионной лимфомы и в клетках многоочаговой болезни Кастлемана. Данный вирус кодирует синтез циклина, который является промотором клеточного деления, чем и объясняется высокая пролиферативная активность инфицированных клеток.
Человеческие папилломавирусы (ПВЧ) вызывают кожные бородавки и ряд доброкачественных папиллом. Это весьма обширный класс вирусов, и некоторые его представители могут в той или иной степени участвовать в развитии злокачественных опухолей. Большинство вирусов вызывает лишь ограниченную пролиферацию клеток, но ряд серотипов может вызвать и развитие злокачественных новообразований.
Наибольшую опасность с точки зрения онкологии представляют серотипы 16, 18, 31 и 33. Более 90% опухолей карциномы шейки матки in situ обнаруживают в своих клетках генные последовательности вирусов ПВЧ. Эти вирусы также играют значительную роль в возникновении инвазивных и многоочаговых плоскоклеточных раков кожи, которые часто развиваются у больных после долгосрочных курсов иммуносупрессивной терапии.
Механизмы вирусного канцерогенеза в этом случае изучены недостаточно. В исследованиях доказано, что существует вероятность включения вирусной ДНК в клетки базального эпителия. Обычно в клеточную ДНК внедряются восемь ранних и два поздних гена. Ранние гены Е6 и Е7 вызывают пролиферацию клеток.
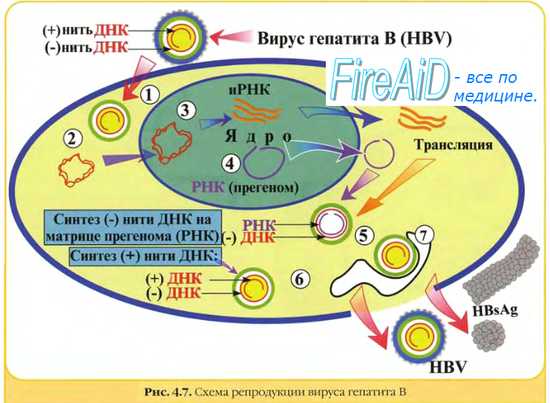
Вирус гепатита В четко связывают с развитием печеночно-клеточного рака. Риск развития этого заболевания у инфицированных вирусом в 200 раз превышает таковой у неинфицированных людей. Механизмы зарождения рака не до конца понятны: считается, что вирус вызывает бесконечную пролиферацию клеток, хотя зараженные вирусом клетки уничтожаются иммунной системой организма. Возможно, что пролиферирующие клетки становятся чувствительными к другим канцерогенам, например к афлатоксину.
Подытоживая вышесказанное, можно заключить, что основным механизмом канцерогенного воздействия является повреждение молекул ДНК и нарушения процессов ее репарации. Если не происходит исправления первичных повреждений ДНК, ошибки со временем начинают накапливаться, что ведет к дестабилизации структуры хромосом и к соматическим мутациям. Большинство таких мутаций летальны для клетки.
Однако, если часть клеток выживает, происходит постепенное накопление клеточной массы с опухолевым фенотипом, что в итоге приводит к клиническим проявлениям. Большой вклад в понимание этого процесса внесли современные исследования механизмов регулировки клеточного цикла и недавнее открытие регуляторных белков, которые запрещают клетке делиться, если ее геном сильно поврежден.
Для развития опухоли может быть недостаточно единичных мутаций в генах-регуляторах деления, но накопление массы ошибок в регуляторных генах может привести к фатальным последствиям. Особенно критичны мутации в генах, регулирующих клеточное деление, так называемых онкогенах. Очень опасны также мутационная инактивация генов, подавляющих клеточное деление — генов-подавителей опухоли, и мутации в генах, кодирующих белки, запрещающие деление при повреждении клеточного генотипа.
Накопление числа жизнеспособных мутаций по одному или нескольким из этих генов приводит к общей клеточной дезорганизации и к потере контроля над клеточным делением. В следующем разделе мы рассмотрим механизмы действия онкогенов и наиболее важные моменты контроля над клеточным циклом.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
История открытия вирусов. Обнаружение в 1901 году первого вирусного заболевания человека — жёлтой лихорадки. Спиральные и икосаэдрические капсиды. Строение Т-фага кишечной палочки со смешанным типом симметрии. Классы ДНК-содержащих опухолеродных вирусов.
| Рубрика | Медицина |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 07.06.2016 |
| Размер файла | 525,8 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
КАЗАХСКИЙ НАЦИОНАЛЬНЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ
ДНК СОДЕРЖАЩИЕ ВИРУСЫ. КЛАССИФИКАЦИЯ ДНК СОДЕРЖАЩИХ ВИРУСОВ
Выполнила: Канафина Т.М.
История открытия вирусов
ДНК - содержащие вирусы
Список использованной литературы
Вирусы от живых организмов - внутриклеточных паразитов отличаются полным отсутствием основного и энергетического обмена, и отсутствием сложнейшего элемента живых систем -- аппарата трансляции (синтеза белка), степень сложности которого превышает таковую самих вирусов.
История открытия вирусов
В 1852 г. русский ботаник Д.И. Ивановский впервые получил инфекционный экстракт из растений табака, пораженных мозаичной болезнью. Когда такой экстракт пропустили через фильтр, способный задерживать бактерии, отфильтрованная жидкость все еще сохраняла инфекционные свойства. В 1898 г. голландец Бейеринк придумал новое слово вирус, чтобы обозначить этим термином инфекционную природу некоторых профильтрованных растительных жидкостей. Хотя удалось достигнуть значительных успехов в получении, высокоочищенных проб вирусов и было установлено, что по химической природе это нуклеопротеины, сами частицы все еще оставались неуловимыми и загадочными, потому что они были слишком малы, чтобы их можно было увидеть с помощью светового микроскопа. Поэтому-то вирусы и оказались в числе первых биологических структур, которые были исследованы в электронном микроскопе сразу же после его изобретения в 30-е годы нашего столетия.
В 1901 году было обнаружено первое вирусное заболевание человека -- жёлтая лихорадка. Это открытие было сделано американским военным хирургом У. Ридом и его коллегами.
В 1911 году Фрэнсис Раус доказал вирусную природу рака -- саркомы Рауса (лишь в 1966 г, спустя 55 лет, ему была вручена за это открытие Нобелевская премия по физиологии и медицине).
Эксперимент Херши. Эксперимент проводился на бактериофаге T2, структура которого к тому времени была выяснена с помощью электронной микроскопии. Оказалось, что бактериофаг состоит из белковой оболочки, внутри которой находится ДНК. Эксперимент был спланирован таким образом, чтобы выяснить, что же -- белок или ДНК -- является носителем наследственной информации.
Херши и Чейз выращивали две группы бактерий: одну в среде, содержащей радиоактивный фосфор-32 в составе фосфат-Иона, другую -- в среде с радиоактивной серой-35 в составе сульфат-Иона. Бактериофаги, добавленные в среду с бактериями и размножавшиеся в них, поглощали эти радиоактивные изотопы, которые служили маркёрами, при построении своей ДНК и белков. Фосфор содержится в ДНК, но отсутствует в белках, а сера, наоборот, содержится в белках (точнее в двух аминокислотах: цистеин и метионин), но её нет в ДНК. Таким образом, одни бактериофаги содержали меченые серой белки, а другие -- меченую фосфором ДНК.
После выделения радиоактивно-меченых бактериофагов их добавляли к культуре свежих (не содержащих изотопов) бактерий и позволяли бактериофагам инфицировать эти бактерии. После этого среду с бактериями подвергали энергичному встряхиванию в специальном смесителе (было показано, что при этом оболочки фага отделяются от поверхности бактериальных клеток), а затем инфицированных бактерий отделяли от среды. Когда в первом опыте к бактериям добавлялись меченые фосфором-32 бактериофаги, оказалось, что радиоактивная метка находилась в бактериальных клетках. Когда же во втором опыте к бактериям добавлялись бактериофаги, меченые серой-35, то метка была обнаружена во фракции среды с белковыми оболочками, но её не было в бактериальных клетках. Это подтвердило, что материалом, которым инфицировались бактерии, является ДНК. Поскольку внутри инфицированных бактерий формируются полные вирусные частицы, содержащие белки вируса, данный опыт был признан одним из решающих доказательств того факта, что генетическая информация (информация о структуре белков) содержится в ДНК.
В 1969 году Алфред Херши получил Нобелевскую премию за открытия генетической структуры вирусов.
В 2002 году, в университете Нью-Йорка был создан первый синтетический вирус
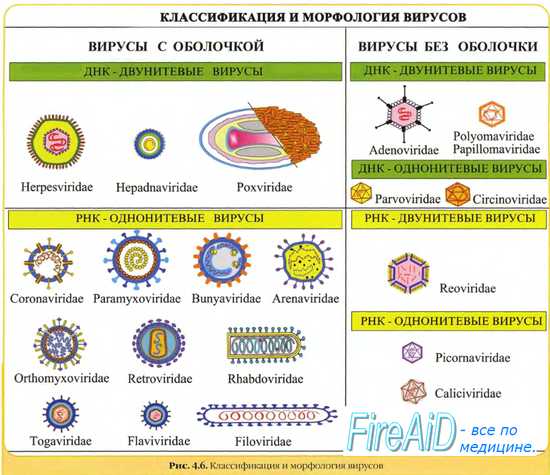
ДНК-содержащие вирусы несут в качестве генетического материала либо одно -, либо двухцепочечную ДНК, которая может быть как линейной, так и кольцевой. В ДНК закодирована информация о всех белках вируса. Вирусы, заражающие бактерии, называются бактериофагами. К ДНК-содержащим вирусам относятся вирусы гепатита В, герпес, вирусы оспы, паповавирусы, гепаднавирусы, парвовирусы.
По виду цепи ДНК вирусы делятся на 3 группы:
Первая группа -- вирусы с двуцепочечной ДНК. Репликация ДНК-генома этих вирусов осуществляется при посредстве промежуточных молекул РНК: Молекулы РНК образуются в результате транскрипции вирусных ДНК в клеточном ядре хозяйским ферментом ДНК-зависимой РНК-полимеразой. Транскрибируется только одна из нитей вирусной ДНК. Синтез ДНК на мРНК происходит в результате реакции, катализируемой обратной транскриптазой: сначала синтезируется (-) нить ДНК, а затем на вновь синтезированной (-) нити ДНК тот же фермент строит (+) нить.
Вторая группа -- вирусы с двуцепочечной ДНК.В одних случаях производством как мРНК, так и ДНК занимаются клеточные ферменты; в других случаях вирусы используют собственные ферменты. Бывает, что те и другие ферменты обслуживают процесс репликации и транскрипции. К этой группе относятся вирусы герпеса, оспы и др.
Тип нуклеиновой кислоты
Симметрия капсидов
Капсид с оболочкой или без нее
Примеры вирусов данной группы:
- обычно встречаются у нитевидных вирусов.
- образуются путем самосборки асимметричных белковых субъединиц (капсомеров), объединяющихся в трубчатую структуру со спиральной симметрией.
- Субъединицы в большинстве случаев гомогенны, так, что поверхность вириона состоит из множества копий одного и того же белка, хотя под наружным капсидом могут находиться и другие белки.
- ДНК в таких вирусах либо вытянута, либо может быть туго скручена в комплексе со специальными связывающими белками.
Спиральный тип симметрии имеет вирус гриппа - а.
Икосаэдрические капсиды
- свойственны большинству сферических ДНК-содержащих вирусов
- икосаэдр - это многогранник с двадцатью треугольными гранями, имеющий кубическую симметрию и приблизительно сферическую форму.
- Вершины треугольников соединяясь образуют двенадцать вершин икосаэдра;
- в местах соединения располагаются обычно пентамерные белковые структуры - пентоны; там же могут находится участки, на которых формируются белковые нити, нередко ассоциированные с вершинами
- Грани икосаэдра заполнены другими белковыми субъединицами, сгруппированными обычно в гексамерные структуры - гексоны (пример, у аденовируса прозрачка 1). вирус капсид симметрия опухолеродный
- Количество субъединиц, необходимое для заполнения граней, определяется размерами вириона в целом, и разные икосаэдрические вирусы содержат поэтому разное число гексонов - обычно при неизменном числе пентонов.
- ДНК обычно плотно свернута внутри капсида;
- иногда она связана с белками или полипептидами, способными стабилизировать ее структуру.
Кубический тип симметрии у вирусов:
герпеса - б
полиомиелита - г.
аденовируса - в
Сложные капсиды без оболочки
- типичны для бактериофагов:
- они состоят из частей с разными типами симметрии.
Фотография бактериофага (увеличение 500000 раз).
строение Т-фага кишечной палочки со смешанным типом симметрии. 1 - кубоидальная капсидная головка, 2 - двухнитчатая ДНК, 3 - стержень, 4 - спиралеобразный сокращающийся капсид (чехол), 5- базальная пластинка, 6 - хвостовые фибриллы.
Сложные капсиды с оболочкой
- есть только у вирусов эукариотических клеток.
- В них ДНК-белковые комплексы окружены одним или несколькими белковыми слоями и наружной мембраной, почти все белковые компоненты которой являются вирусными по своему происхождению, а липидные структуры - клеточными.
ДНК-содержащие опухолеродные вирусы разделяются на следующие 5 классов.
1. Полиомавирусы - обезьяний вирус SV40, вирус полиомы мышей и вирусы человека ВК и JC.
2. Папилломавирусы - 16 вирусов папилломы человека и множество папилломовирусов животных.
3. Аденовирусы - 37 вирусов человека, множество аденовирусов животных (например, 24 вируса обезьян и 9 вирусов крупного рогатого скота).
4. Герповирусы - вирусы простого герпеса человека, цитомегаловирус человека, вирус Эпштейна-Барр и онкогенные вирусы приматов, лошадей, кур, кроликов, лягушек.
5. Вирусы, подобные вирусу гепатита В, - вирус гепатита В человека, гепатита североамериканского сурка, гепатита земляных белок и гепатита уток.
Систематику и таксономию вирусов кодифицирует и поддерживает Международный Комитет по Таксономии Вирусов (International Committee on Taxonomy of Viruses, ICTV), поддерживающий также и таксономическую базу The Universal Virus Database ICTVdB.
Классификация ICTV
Международным Комитетом по Таксономии Вирусов в 1966 году была принята система классификации вирусов основанная на различии типа (РНК и ДНК), количества молекул нуклеотических кислот (одно- и двух-цепочечные) и на наличии или отсутствии оболочки ядра. Система классификации представляет собой серию иерархичных таксонов:
Отряд (-virales)
Семейство (-viridae)
Подсемейство (-virinae)
Классификация Балтимора
Нобелевский лауреат, биолог Дэвид Балтимор, предложил свою схему классификации вирусов, основываясь на различиях в механизме продукции мРНК. Эта система включает в себя семь основных групп:
(I) Вирусы, содержащие двуцепочечную ДНК и не имеющие РНК-стадии (например, герпесвирусы, поксвирусы, паповавирусы, мимивирус).
(II) Вирусы, содержащие двуцепочечную РНК (например, ротавирусы).
(III) Вирусы, содержащие одноцепочечную молекулу ДНК (например, парвовирусы).
(IV) Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности (например, пикорнавирусы, флавивирусы).
(V) Вирусы, содержащие одноцепочечную молекулу РНК негативной или двойной полярности (например, ортомиксовирусы, филовирусы).
(VI) Вирусы, содержащие одноцепочечную молекулу РНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретровирусы (например, ВИЧ).
(VII) Вирусы, содержащие двуцепочечную ДНК и имеющие в своём жизненном цикле стадию синтеза ДНК на матрице РНК, ретроидные вирусы (например, вирус гепатита B).
В настоящее время, для классификации вирусов используются обе системы одновременно, как дополняющие друг друга.
Дальнейшее деление производится на основе таких признаков как структура генома (наличие сегментов, кольцевая или линейная молекула), генетическое сходство с другими вирусами, наличие липидной оболочки, таксономическая принадлежность организма-хозяина и так далее.
В результате проведенной работы, я изучила историю открытия, виды, строение, размножение вирусов, вирусные заболевания.
На основе моего изучения можно сделать вывод, что вирусы весьма разнообразны и многие из них остаются нераспознанными. Для того чтобы знать лечение и причины возникновения различных заболеваний, мы должны тщательно изучать все особенности вирусов и вирусных инфекций.
Список использованной литературы
Подобные документы
Проблемы борьбы с вирусами - возбудителями заболеваний. История открытия вирусов, их формы. Многообразие строения вирусов. Особенности вирусов, их классификация и этапы жизнедеятельности. Анализ строения бактериофага. Вирусные заболевания человека.
презентация [576,5 K], добавлен 12.05.2013
Гипотезы происхождения, история открытия вирусов, их строение и химические свойства. Классификация вирусов, их взаимодействие с клеткой. Способы передачи вирусных заболеваний: оспа, гепатит, грипп, полиомиелит, СПИД. Эволюция вирусов на современном этапе.
реферат [46,4 K], добавлен 20.12.2009
Свойства вирусов и плазмид, по которым они отличаются от остального живого мира. Морфология вирусов. Исходы взаимодействия вирусов с клеткой хозяина. Методы культивирования вирусов. Вирусы бактерий (бактериофаги). Этапы взаимодействия фагов и бактерий.
реферат [25,6 K], добавлен 21.01.2010
История возникновения вирусов, простые и сложные вирусы. Содержание теории регрессивного происхождения вирусов. Основания для выдвижения эндогенного происхождения вирусов. Основные недостатки теории происхождения вирусов из доклеточных форм жизни.
презентация [5,7 M], добавлен 10.10.2019
Сущность понятия "вирус", история изучения. Схематическое строение вируса. Классификация вирусов: дезоксивирусы, рибовирусы. Схематичное изображение расположения капсомеров в капсиде вирусов. Вирус иммунодефицита человека, трехмерное изображение.
презентация [496,1 K], добавлен 19.10.2011
Классификация и типы ретровирусов как носителей и активаторов онкогенов: высокоонкогенные, низкоонкогенные, механизм действия. Строение, элементы и цикл развития данных вирусов. T-лимфотропные вирусы человека: эпидемиология, описание, профилактика.
курсовая работа [1,3 M], добавлен 27.06.2011
Открытие первого вируса, поражающего человека, его проникновение в клетку. Этапы развития вирусологии. Использование лабораторных мышей и куриных эмбрионов для культивирования вирусов. Строение и химический состав вириона. Выход вирионов из клетки.
Общая характеристика и классификация ДНК-геномных вирусов как вирусов, геном которых представлен дезоксирибонуклеиновой кислотой. Характеристика, виды, онкогенность, репликация и лабораторная диагностика вируса папилломы и вируса полиомы человека.
| Рубрика | Медицина |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 10.12.2010 |
| Размер файла | 295,0 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
ПЛАН:
1. Классификация и характеристика ДНК-геномных вирусов
2. Вирус папилломы человека. Характеристика. Онкогенность. Лабораторная диагностика
3. Вирус полиомы человека. Характеристика. Онкогенность
1. Классификация и характеристика ДНК-геномных вирусов
ДНК-содержащие вирусы - вирусы, геном которых представлен дезоксирибонуклеиновой кислотой и репликация идет посредством ДНК-зависимой ДНК-полимеразы, без использования промежуточного звена-посредника РНК.
Геномная ДНК этих вирусов может быть двуцепочечной или одноцепочечной и иметь линейную или кольцевую форму.
Размер вириона в нанометрах
Аденовирусы человека 42 типов
Вирус гепатита В
Вирусы простого герпеса, цитомегалии, Эпстайна-Барр
Вирусы папилломы, полиомы
Геномы большинства ДНК-содержащих вирусов (за исключением поксвирусов) транскрибируются в ядре клетки-хозяина с помощью клеточной РНК-полимеразы II . Синтез и процессинг мРНК у этих вирусов практически полностью осуществляется за счет клеточных механизмов (несколько вирусных белков участвуют только в регуляции транскрипции).
У многих ДНК-содержащих вирусов транскрипция происходит в три этапа, во время которых избирательно считываются сверхранние, ранние и поздние гены.
Нуклеокапсиды герпесвирусов проникают в цитоплазму вместе с белками матрикса, затем этот комплекс транспортируется вдоль микротрубочек к порам ядерной оболочки и далее в ядро. Белки матрикса вместе с клеточными факторами транскрипции запускают транскрипцию сверхранних генов. У герпесвирусов продукты сверхранних генов, так называемые сверхранние белки, играют роль трансактиваторов и необходимы для начала транскрипции ранних генов. Однако у других ДНК-содержащих вирусов транскрипция ранних генов может происходить и без участия сверхранних белков.
Большинство ранних генов кодируют белки, осуществляющие репликацию вирусной ДНК и запуск транскрипции поздних генов. Поздние гены кодируют главным образом структурные белки, необходимые для сборки вирусов и их выхода из зараженной клетки. Транскрипция поздних генов зависит от репликации ДНК и прекращается в присутствии ингибиторов репликации.
Для каждого семейства ДНК-содержащих вирусов характерен уникальный механизм репликации ДНК.
Геном герпесвирусов представлен линейной ДНК, которая в зараженной клетке замыкается в кольцо. Кольцевые молекулы вирусной ДНК реплицируются по механизму "катящегося кольца" с образованием длинных линейных конкатемерных молекул, состоящих из многих копий генома. Затем конкатемерные молекулы расщепляются на полноценные геномы. В репродукции герпесвирусов участвуют вирусные ДНК-полимераза и ферменты, увеличивающие внутриклеточную концентрацию дезоксинуклеозидтрифосфатов - структурных элементов ДНК.
Репликация линейных геномов аденовирусов происходит при участии вирусной ДНК-полимеразы и нуклеотид-белкового комплекса, выполняющего роль праймера.
Двухцепочечные кольцевые геномы паповавирусов реплицируются с сохранением кольцевой структуры. Репликация осуществляется клеточными ферментами, а для запуска репликации необходим ранний вирусный белок - Т-антиген.
Изредка геномы паповавирусов (например, папилломавирусов) встраиваются в хромосому зараженной клетки, что приводит к усиленной экспрессии вирусных белков и неконтролируемому делению клетки. Интеграция паповавирусов в клеточный геном может стать причиной злокачественного новообразования, например рака шейки матки.
В отличие от репродукции других ДНК-содержащих вирусов, репродукция поксвирусов полностью происходит в цитоплазме. Эти вирусы содержат факторы транскрипции, РНК-полимеразу, а также набор ферментов, обеспечивающих процессинг мРНК (присоединение 7-метилгуанозина, полиаденилирование). Геном поксвирусов имеет уникальное строение: комплементарные цепи двухцепочечной ДНК на обоих концах ковалентно соединены между собой, так что в действительности геном состоит из одноцепочечной кольцевой молекулы ДНК. Кроме того, в геноме имеются инвертированные концевые повторы. Репликация вирусной ДНК начинается с расщепления одной из цепей в области инвертированных концевых повторов; затем одновременно на обоих концах генома начинается синтез комплементарной цепи, который осуществляет вирусная ДНК-полимераза. Праймером служит прилегающий к месту расщепления короткий участок расщепленной цепи (самозатраванный механизм). Поксвирусы, как и герпесвирусы, кодируют несколько ферментов, способствующих репликации ДНК путем увеличения концентрации дезоксирибонуклеозидтрифосфатов.
2. Вирус папилломы человека. Характеристика. Онкогенность. Лабораторная диагностика
Папилломавирус человека (HPV - Human Papillomavirus) - вирус из рода папилломавирусов, семейства паповавирусов. Передаётся только от человека к человеку и приводит к изменению характера роста тканей. Известно более 100 видов ВПЧ. Из них более 40 - могут вызвать поражение аногенитального тракта (половые органы и задний проход) мужчин и женщин и появление остроконечных кондилом. Некоторые из них безвредны, другие вызывают бородавки, некоторые вызывают рак.
HPV - одна из наиболее распространённых болезней, передаваемых половым путём.
Жизненный цикл вируса:
Характерные проявления инфекции - простые бородавки.
В настоящее время известно уже более 100 типов (штаммов) папилломавируса, обнаруженных у человека. Большинство представителей обладает онкогенными свойствами и вызывает доброкачественные или злокачественные опухоли (папилломы). Доказана этиологическая роль папилломавируса в развитии рака шейки матки. Папилломавирусы имеют видоспецифические и некоторые типоспецифические антигены. Не культивируются в клеточных системах.
Вирус папилломы человека выявляется в кератоцитах в виде эписомы, 50 - 200 копий на пораженную клетку. Процессы продуктивной инфекции и интеграции не наблюдаются.
Пути инфицирования:
Папилломавирусы являются антропонозными возбудителями - то есть передача их возможна только от человека к человеку.
Есть вероятность сохранения вирусов в отшелушивающих клетках кожи определенное время - поэтому для некоторых заболеваний, вызываемых вирусом, вероятен контактно-бытовой путь заражения (бородавки) при условии микроповреждений кожи.
Основным путем заражения аногенитальными бородавками (остроконечными кондилломами) является половой путь заражения (включая орально - генитальные контакты (крайне редко) и анальный секс).
Возможно заражение папилломавирусом новорожденных при родах, что является причиной возникновения ларингеального папилломатоза у детей и аногенитальных бородавок у младенцев.
Попав в организм, вирусы папиллом инфицируют базальный слой эпителия, причем наиболее пораженным участком является зона перехода многослойного плоского эпителия в цилиндрический эпителий. В зараженной клетке вирус существует в двух формах: эписомальной (вне хромосом клетки), которая считается доброкачественной формой, и интросомальной - интегрированной (встраиваясь в геном клетки) - которую определяют как злокачественную форму паразитирования вируса.
Типы папилломавирусов:
Условно все известные папилломавирусы можно разбить на три основные группы:
1) Неонкогенные папилломавирусы (HPV 1,2,3,5)
2) Онкогенные папилломавирусы низкого онкогенного риска (в основном HPV 6,11,42,43,44)
3) Онкогенные папилломавирусы высокого онкогенного риска (HPV 16,18,31,33,35,39,45,51,52,56,58,59 и 68)
Диагностика папилломавирусной инфекции
1. Клинический осмотр.
По характерной клинической картине выявляются все виды бородавок, остроконечные кондиломы. При наличии аногенитальных бородавок обязателен осмотр шейки матки, по показаниям - для исключения эндоуретральных кондилом - уретроскопия
2. Кольпоскопия. Кольпоскопия и биопсия показаны всем женщинам с цервикальной интраэпителиальной неоплазией класса II(CIN II) или класса III(CIN III), независимо от подтверждения у них наличия ВПЧ-инфекции Тест с уксусной кислотой (Acetic Acid Test). В настоящее время специфическим кольпоскопическим признаком ПВИ шейки матки считается неравномерное поглощение йодного раствора Люголя беловатым после уксуса участком эпителия (в виде йодпозитивных пунктации и мозаики). Признаками ПВИ шейки матки могут также быть ацетобелый эпителий, лейкоплакия, пунктация, белые выросты и мозаика, атипичная зона трансформации, жемчужная поверхность после обработки уксусом.
3. Цитологическое исследование шеечных мазков по Папаниколау.
Цитологическое исследование шеечных мазков по Папаниколау (PAP - smear test) выделяет следующие результаты:
1-й класс - атипические клетки отсутствуют, нормальная цитологическая картина.
2-й класс - изменение клеточных элементов обусловлено воспалительным процессом во влагалище и (или) шейки матки.
3-й класс - имеются единичные клетки с изменениями соотношения ядра и цитоплазмы, диагноз недостаточно ясен, требуется повторение цитологического исследования или необходимо гистологическое исследование биоптированной ткани для изучения состояния шейки матки.
4-й класс - обнаруживаются отдельные клетки с признаками злокачественности, а именно с увеличенными ядрами и базофильной цитоплазмой, неравномерным распределением хроматина.
5-й класс - в мазке имеются многочисленные атипические клетки.
4. Гистологическое исследование.
Обнаруживается умеренное утолщение рогового слоя с папилломатозом, паракератозом и акантозом; могут присутствовать фигуры митоза. Диагностически важным считается наличие в глубоких участках мальпигиева слоя койлоцитов - больших эпителиальных клеток с круглыми гиперхромными ядрами и выраженной перинуклеарной вакуолизацией.
5. Типирования папилломавирусов с помощью полимеразной цепной реакции (ПЦР).
Метод ПЦР имеет большую диагностическую значимость и позволяет идентифицировать отдельные типы ВПЧ. Однако использование этого метода как диагностического критерия для неопластических процессов шейки матки приводит к значительной гипердиагностике, так как примерно в 80 % случаев инфицирование имеет кратковременный характер и заканчивается спонтанным выздоровлением и элиминацией вируса. Таким образом, положительный результат при лабораторном исследовании на ДНК ВПЧ не позволяет в большинстве случаев прогнозировать развитие цервикального рака. Однако он имеет большую прогностическую значимость, особенно если на фоне ВПЧ -- инфекции уже имеется картина дисплазии эпителия шейки матки, и позволяет говорить о степени канцерогенного риска.
6. Исследование на заболевания, передающиеся половым путем
Так как аногенитальные бородавки в 90 % случаев ассоциируются с другими урогенитальными инфекциями, целесообразно проводить исследования на их наличие методом ПЦР.
3. Вирус полиомы человека. Характеристика. Онкогенность
Полиомавирус относится к семейству Роlyomaviridae. У человека патологию вызывают два вида: ВК и JC. ВК-вирус встречается в 60-90% случаев. В основном болеют дети. Развивается респираторная инфекция без признаков почечной патологии. Поражение почек возможно при воздействии полиомавируса ВК. Другими представителями группы полиомавирусов являются вирус JС, вызывающий прогрессивную мультифокальную энцефалопатию у человека, и вирус SV-40, являющийся лимфотропным вирусом приматов.
Широкое распространение полиомавирусной инфекции (60-100% населения является серопозитивным) с локализацией ее в В-клетках, в переходном эпителии, а также в тубулярном эпителии почек не сопровождается возникновением воспалительных изменений.
Активация вируса ВК наблюдается при подавлении Т-клеточного иммунитета. В связи с этим полиомавирусная инфекция ассоциируется с поражением почечного трансплантата, нефропатией при ВИЧ-инфекции. Описаны также случаи интерстициального нефрита у лиц с другими причинами иммунодепрессивного синдрома. Однако появились данные об ассоциации гематурии с инфекцией вирусом ВК в отсутствие иммунодефицитного состояния. Известны также случаи активации инфекции у больных, страдающих сахарным диабетом при диабетической нефропатии.
У иммунодефицитных пациентов в нефробиоптате Полиомавирус выявляется в 17,9% случаев, в моче - в 16,2% случаев.
В большинстве случаев патология почек описывается как полиомавирус-ассоциированная нефропатия. Вирусные включения обнаруживаются в канальцевом эпителии. Полиомавирус ВК обнаруживается в почках в 10% аутопсий, нередко обнаруживается в моче пожилых лиц.
Частая персистенция вируса в почке (до 65% при иммуногистохимическом исследовании) ассоциируется с развитием нефропатии лишь в 2-6% случаев.
При этом основным отличием инфекции вирусом ВК у реципиентов почечного аллотрансплантата от цитомегаловирусной и Эпштейна-Барр вирусной инфекции служит отсутствие системных проявлений. Нефропатия проявляется прогрессивным снижением функции трансплантата. Морфологическими проявлениями полиомавирусной нефропатии могут быть интерстициальный нефрит с мононуклеарной инфильтрацией, тубулит, наличие интрануклеарных включений.
Онкогенные вирусы могут оказывать либо прямое трансформирующее, либо непрямое промоторное действие.
Онкогенную активность вирусов определяет особый трансформирующий ген (онкоген). Принципиальное отличие вирусных онкогенов от клеточных онкогенов - отсутствие гомолога в геноме нормальной клетки.
Вирусы, содержащие онкоген в своём геноме, обозначают как оnс+-вирусы. Большинство из них ДНК-геномные.
Однако довольно часто эти вирусы утрачивают онкоген и, соответственно, трансформирующую активность; в этом случае их обозначают как onс-вирусы.
Онкоген не кодирует репродукцию жизненно важных для вируса белков, и его утрата не лишает вирус способности репродуцироваться в чувствительных клетках. В зависимости от особенностей структуры вирусного генома выделяют независимые и слитые онкогены.
Независимые онкогены транслируются в виде отдельного белка, не содержащего продуктов экспрессии соседнего гена. При этом независимый онкоген может быть локализован как на конце вирусного генома (например, ген sre у вируса саркомы Рауса), так и в его середине (например, ген ras у вируса саркомы мышей).
Слитые онкогены транскрибируются в виде РНК, кодирующей опухолевый белок в составе единой молекулы, несущей аминокислотную последовательность структурных белков вирусов.
Литература:
1. Коротяев А. И. Медицинская микробиология, иммунология и вирусология, СПб.: СпецЛит, 2002;
Подобные документы
Понятие вируса папилломы как эпителиотропного вируса, способного поражать клетки эпителия эктодермального происхождения, его типы и формы. Этапы и клиническая картина протекания заболевания, его диагностика и подходы к лечению, этиология и патогенез.
реферат [359,5 K], добавлен 01.06.2015
Строение вируса папилломы человека. Процесс проникновения вируса в клетку. Множественные кондиломы шейки матки. Причины, которые способствуют развитию заболевания при попадании папилломовирусной инфекции в организм. Возникновение необратимой неоплазии.
презентация [2,2 M], добавлен 08.12.2011
Папилломовирусная инфекция - хроническое инфекционное заболевание: возбудители и симптомы. Пути передачи вируса папилломы, типы заболевания. Специфика бовеноидного папулеза. Проявление вируса папилломы у мужчин и у женщин. Диагностика и лечение болезни.
презентация [5,0 M], добавлен 13.11.2012
Сущность понятия "вирус", история изучения. Схематическое строение вируса. Классификация вирусов: дезоксивирусы, рибовирусы. Схематичное изображение расположения капсомеров в капсиде вирусов. Вирус иммунодефицита человека, трехмерное изображение.
презентация [496,1 K], добавлен 19.10.2011
Теории происхождения, история изучения и открытия вирусов. Их жизненный цикл, роль в заболеваниях человека, биосфере и эволюции. Морфологические типы капсидов. Формирование липидной оболочки вируса. Виды вирусных инфекций человека, растений, бактерий.
курсовая работа [2,3 M], добавлен 18.05.2016
Биология вируса иммунодефицита человека. Группы высокого риска инфицирования ВИЧ. Стадии, лабораторная диагностика и клиническая классификация ВИЧ-инфекции. Поражение нервной системы человека. Факторы, влияющие на распространение вируса половым путем.
реферат [33,3 K], добавлен 10.05.2009
Характеристика методики выявления дезоксирибонуклеиновой кислоты вируса Эпштейна-Барр у больных с различной инфекционной патологией. Определение чувствительности и специфичности выявления ДНК вируса Эпштейна-Барр у больных инфекционным мононуклеозом.
Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных.
Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
Морфология вирусов. Размеры вирусов.
Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки.
Нуклеиновые кислоты вирусов
Вирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине.
Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180") повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
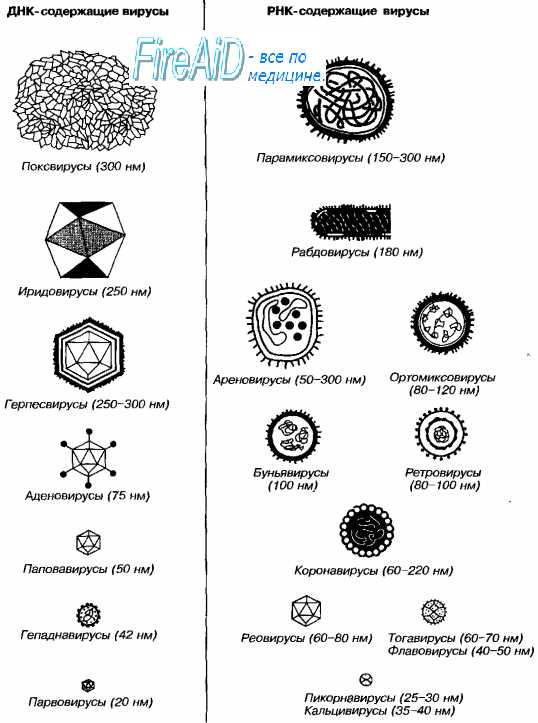
Рис. 2-1. Размеры и морфология основных возбудителей вирусных инфекций человека.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК.
Инфекционность нуклеиновых кислот вирусов
Многие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: