
Этапы синтеза белка вируса
Обновлено: 11.05.2024
Этап синтеза вирусных компонентов. Синтез вирусных компонентов - эклипс-период.
Вскоре после обнажения вирусного генома происходит уменьшение или исчезновение инфекционности, поскольку вирионов как таковых уже не существует, а инфекционность нуклеиновой кислоты намного меньше инфекционности полных вирионов. Данное явление было названо эклипсом (затмением), а фаза вирусной инфекции с момента адсорбции и до появления в клетке новых инфекционных вирионов — эклипс-фазой, или латентным периодом. При высокой множественности заражения клеток некоторая часть вирионов сохраняет инфекционные свойства в течение всей эклипс-фазы, что зачастую затрудняет определение ее продолжительности.
В течение этого периода происходят все существенные процессы вирусной репродукции, приводящие к образованию вирусных компонентов и формированию нового потомства вирионов. С появлением в клетке первых вновь синтезированных вирионов завершается эклипс-период размножения вируса.
Процессы, происходящие в эклипс-фазе, схематично показаны на рисунке, где в качестве примера представлены основные этапы репликации аденовирусов.
Из представленной схемы видно, что после прикрепления вирион входит в клетку и частично или полностью лишается оболочки, обнажая вирусный геном. Некоторые ранние вирусные гены транскрибируются с образованием мРНК, которые могут затем изменяться различными путями, включая сплайсинг. Ранние мРНК транслируются с образованием трех главных продуктов: белков, которые подавляют или останавливают синтез белков и нуклеиновых кислот; белков, которые регулируют экспрессию вирусного генома; ферментов, необходимых для репликации вирусных нуклеиновых кислот. Вслед за репликацией вирусной нуклеиновой кислоты транскрибируются поздние вирусные гены, продуктами трансляции которых являются в основном структурные вирусные белки, используемые при сборке новых вирионов. Некоторые из них претерпевают посттрансляционные изменения перед включением в структуру вирионов. В каждой инфицированной клетке образуются сотни и тысячи вирионов, которые, освобождаясь, заражают другие клетки.
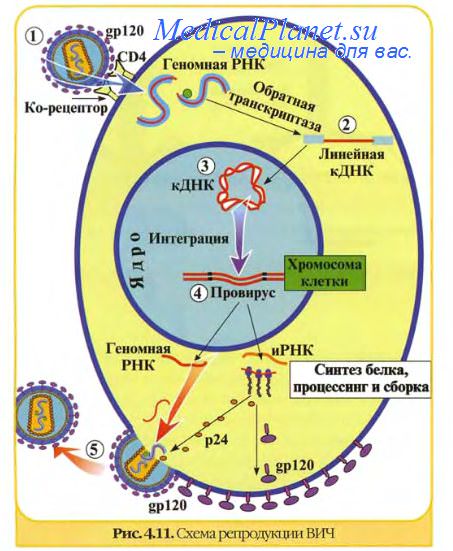
У большинства ДНК-вирусов транскрипция и репликация ДНК происходит в ядре клетки, с использованием клеточной РНК-полимеразы II и других клеточных ферментов. Большинство РНК-вирусов размножается в цитоплазме, где отсутствуют ферменты, копирующие РНК на РНК-матрице, поэтому вирусный геном должен сам по себе функционировать как мРНК или вирус должен нести свою РНК-полимеразу, чтобы транскрибировать РНК на РНК-геноме.
Синтез вирусных белков происходит только в цитоплазме клетки. В инфицированных клетках вирусные нуклеиновые кислоты и вирусспецифические белки синтезируются в значительно большем количестве, чем включаются в вирионы. Избыточный синтез структурных компонентов при репродукции вирусов является своеобразной платой за паразитизм на генетическом уровне.
Потомство вирионов проявляется спустя определенный период после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность периода от момента инфицирования до появления дочерних вирионов довольно вариабельна и отражает глубокое различие между разными вирусами.
Латентный период мелких вирусов, не имеющих оболочки и содержащих односпиральную РНК, длится примерно 2 часа и является наиболее коротким среди всех известных вирусов животных. ДНК-содержащие вирусы имеют более продолжительный латентный период. Длительный латентный период отмечен также у реовирусов и вируса саркомы Рауса. У простых мелких РНК-содержащих вирусов быстрее протекает не только латентный период, но и весь цикл размножения вируса. Вирусы, геном которых выполняет функции мРНК, имеют более короткий репликативный цикл, чем вирусы, у которых функции мРНК выполняет не вирионная РНК, а комплементарная ей минус-нить.
Размножение ДНК-содержащих вирусов может продолжаться даже через 24 часа после заражения. Причины, определяющие длительность цикла размножения разных вирусов, не выяснены. Возможно, что это связано с более сложной последовательностью событий при репродукции крупных, сложно устроенных вирусов.
Укорочение латентного периода при высокой множественности заражения было обнаружено у различных РНК и ДНК вирусов и, возможно, связано с ускорением процесса образования ферментов, участвующих в синтезе нуклеиновой кислоты. При высокой множественности заражения не исключается также роль капсидных белков в ускорении начала цикла репродукции.
Число инфекционных вирусных частиц, образуемых в одной клетке, зависит от типа вируса, и количество их варьирует очень широко. Интересно, что на долю вирусспецифических продуктов приходится от 0,1 до 5% массы клетки животного.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.
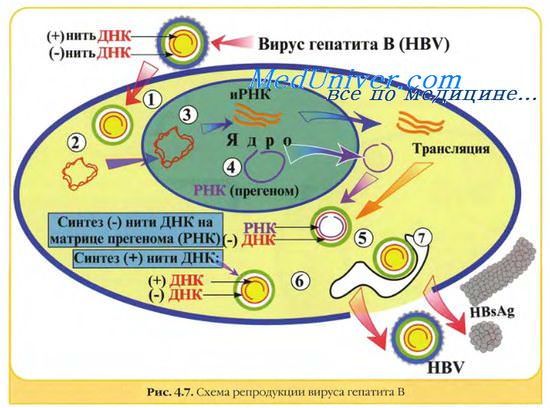
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Этап синтеза вирусных компонентов. Синтез вирусных компонентов - эклипс-период.
Вскоре после обнажения вирусного генома происходит уменьшение или исчезновение инфекционности, поскольку вирионов как таковых уже не существует, а инфекционность нуклеиновой кислоты намного меньше инфекционности полных вирионов. Данное явление было названо эклипсом (затмением), а фаза вирусной инфекции с момента адсорбции и до появления в клетке новых инфекционных вирионов — эклипс-фазой, или латентным периодом. При высокой множественности заражения клеток некоторая часть вирионов сохраняет инфекционные свойства в течение всей эклипс-фазы, что зачастую затрудняет определение ее продолжительности.
В течение этого периода происходят все существенные процессы вирусной репродукции, приводящие к образованию вирусных компонентов и формированию нового потомства вирионов. С появлением в клетке первых вновь синтезированных вирионов завершается эклипс-период размножения вируса.
Процессы, происходящие в эклипс-фазе, схематично показаны на рисунке, где в качестве примера представлены основные этапы репликации аденовирусов.
Из представленной схемы видно, что после прикрепления вирион входит в клетку и частично или полностью лишается оболочки, обнажая вирусный геном. Некоторые ранние вирусные гены транскрибируются с образованием мРНК, которые могут затем изменяться различными путями, включая сплайсинг. Ранние мРНК транслируются с образованием трех главных продуктов: белков, которые подавляют или останавливают синтез белков и нуклеиновых кислот; белков, которые регулируют экспрессию вирусного генома; ферментов, необходимых для репликации вирусных нуклеиновых кислот. Вслед за репликацией вирусной нуклеиновой кислоты транскрибируются поздние вирусные гены, продуктами трансляции которых являются в основном структурные вирусные белки, используемые при сборке новых вирионов. Некоторые из них претерпевают посттрансляционные изменения перед включением в структуру вирионов. В каждой инфицированной клетке образуются сотни и тысячи вирионов, которые, освобождаясь, заражают другие клетки.
У большинства ДНК-вирусов транскрипция и репликация ДНК происходит в ядре клетки, с использованием клеточной РНК-полимеразы II и других клеточных ферментов. Большинство РНК-вирусов размножается в цитоплазме, где отсутствуют ферменты, копирующие РНК на РНК-матрице, поэтому вирусный геном должен сам по себе функционировать как мРНК или вирус должен нести свою РНК-полимеразу, чтобы транскрибировать РНК на РНК-геноме.
Синтез вирусных белков происходит только в цитоплазме клетки. В инфицированных клетках вирусные нуклеиновые кислоты и вирусспецифические белки синтезируются в значительно большем количестве, чем включаются в вирионы. Избыточный синтез структурных компонентов при репродукции вирусов является своеобразной платой за паразитизм на генетическом уровне.
Потомство вирионов проявляется спустя определенный период после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность периода от момента инфицирования до появления дочерних вирионов довольно вариабельна и отражает глубокое различие между разными вирусами.
Латентный период мелких вирусов, не имеющих оболочки и содержащих односпиральную РНК, длится примерно 2 часа и является наиболее коротким среди всех известных вирусов животных. ДНК-содержащие вирусы имеют более продолжительный латентный период. Длительный латентный период отмечен также у реовирусов и вируса саркомы Рауса. У простых мелких РНК-содержащих вирусов быстрее протекает не только латентный период, но и весь цикл размножения вируса. Вирусы, геном которых выполняет функции мРНК, имеют более короткий репликативный цикл, чем вирусы, у которых функции мРНК выполняет не вирионная РНК, а комплементарная ей минус-нить.
Размножение ДНК-содержащих вирусов может продолжаться даже через 24 часа после заражения. Причины, определяющие длительность цикла размножения разных вирусов, не выяснены. Возможно, что это связано с более сложной последовательностью событий при репродукции крупных, сложно устроенных вирусов.
Укорочение латентного периода при высокой множественности заражения было обнаружено у различных РНК и ДНК вирусов и, возможно, связано с ускорением процесса образования ферментов, участвующих в синтезе нуклеиновой кислоты. При высокой множественности заражения не исключается также роль капсидных белков в ускорении начала цикла репродукции.
Число инфекционных вирусных частиц, образуемых в одной клетке, зависит от типа вируса, и количество их варьирует очень широко. Интересно, что на долю вирусспецифических продуктов приходится от 0,1 до 5% массы клетки животного.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Этапы трансляции мРНК при синтезе белка
На протяжении нескольких последних десятилетий контроль над экспрессией генов на уровне трансляции стал способом регулирования роста, пролиферации, злокачественной трансформации и апоптоза. Регулирование трансляции мРНК может изменить общую трансляционную или рибосомную емкость клетки (т.е. число рибосом) либо изменить трансляционную или рибосомную эффективность (т.е. количество белка, синтезированного на одной рибосоме).
Кроме того, изменения рибосомной эффективности могут ограничиваться регулированием экспрессии конкретного белка (например, мРНК ферритина) или влиять на скорость трансляции специфических белков, которые затем участвуют в трансляции мРНК и, следовательно, оказывают глобальное всеобъемлющее влияние на производительность и эффективность трансляции мРНК (например, терминального олигопиримидина мРНК, который описан далее).
Изменение рибосомной емкости указывает на изменение содержания рибосомных белков и рибосомной РНК (рРНК) в клетке. Время, необходимое для модификации этих популяций, показывает, что изменить общий синтез белка на уровне рибосомной емкости могут лишь хронические заболевания или такие продолжительные состояния, как длительное голодание или неконтролируемый диабет.
На уровень РНК также влияет возраст. В частности, скорость общего оборота рРНК и транспортной РНК (тРНК) во всем организме у недоношенных младенцев в 3-4 раза выше, чем у взрослых, а оборот мРНК у новорожденных в 6 раз выше, чем у взрослых. Это подчеркивает тот факт, что высокий темп роста у новорожденных возможен благодаря способности организма создавать и использовать трансляционный аппарат.
Каждая рибосома состоит из 80 различных белков и 4 видов РНК (5S, 5.8S, 18S, 28S). Три вида РНК (5.8S, 18S, 28S) транскрибируются с одного гена (45S рибосомная ДНК (рДНК)) РНК-полимеразой типа I. Транскрипция РНК-полимеразы типа I усиливается фактором транскрипции UBF (фактором регуляции последующих звеньев сигнальной цепи). Состояния, усиливающие процессы анаболизма, могут сопровождаться повышенной экспрессией и/или активностью белка UBF, в результате увеличивается транскрипция 45S рДНК. Например, повышенная активность UBF способствует гипертрофии кардиомиоцитов новорожденных.
На рибосомную емкость также может оказывать влияние синтез рибосомного белка. Регуляция его синтеза происходит за счет наличия последовательности пиримидинов в 5-нетранслируемом участке гена. Этот фрагмент терминального олигопиримидина (TOP) преимущественно обнаруживается в мРНК, кодирующей рибосомные белки, факторы трансляции мРНК, такие как факторы элонгации eEFIA и eEF2, и поли-(А)-связывающий белок, участвующие в образовании рибосом.
Сокращение трансляции TOP мРНК снижает общую белковосинтетическую емкость клетки, поскольку TOP мРНК кодирует трансляционный аппарат. Ряд исследований связал воедино фосфатидилинозитол-3-киназу и сигнальный путь mTOR в трансляционной активации TOP мРНК, однако на сегодняшний день точная роль каждого пути в этом процессе полностью не определена. Питание продуктами, содержащими белок, повышает уровень трансляции TOP мРНК, в то время как диета, обедненная аминокислотами, его снижает.
Сигнальный путь, важный при регулировании трансляции ТОР мРНК и, следовательно, при биогенезе рибосом, включает протеинкиназу mTOR. mTOR образует отдельные сигнальные комплексы с дополнительными взаимодействующими белками, которые затем побуждают трансляционный аппарат адекватно реагировать на питательные вещества, инсулин/факторы роста и энергетический статус. Таким образом, путь, преобразующий сигнал mTOR, является важным механизмом, посредством которого эукариотические клетки настраивают свой потенциал биосинтеза белка в ответ на условия среды, окружающей клетку.
Нутриент-зависимый комплекс 1 mTOR, реагирующий как на гормональный, так и на аминокислотный статус, активирует сигнальный путь киназы рибосомного белка S6 (S6K1), контролирующий клеточную гипертрофию и гомеостаз глюкозы. Нутриент-зависимый комплекс 2 mTOR является важным фактором регуляции синтеза актинового цитоскелета и развития и ремоделирования тканей. Кроме того, mTOR контролирует клеточную пролиферацию, синтез рРНК, процессинг пре-рРНК и аутофагию. Таким образом, сигнальный путь mTOR контролирует широкий спектр различных событий, связанных с ростом и развитием.
Надо отметить, что mTOR и его сигнальные комплексы занимают центральное место во всех процессах, касающихся роста, и, следовательно, необходимы для роста новорожденных и анаболической реакции на прием нутриентов.
Регулирование рибосомной емкости дает организму возможность адаптироваться к условиям длительных изменений. С другой стороны, при необходимости изменений рибосомной эффективности возможно достичь без промедления, поскольку весь белковосинтетический аппарат уже присутствует. В перечень условий, которые изменяют рибосомную эффективность, входят питание, гипоксия и гормональные колебания. Изменения рибосомной эффективности предполагают регулирование на уровне трансляции мРНК.
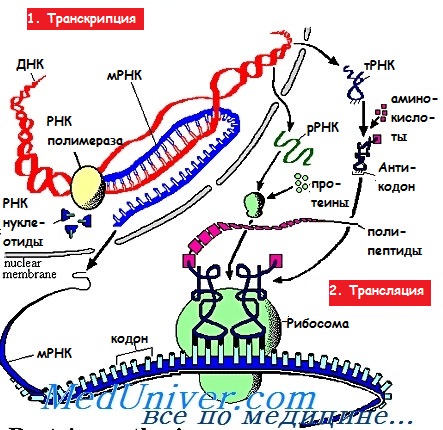
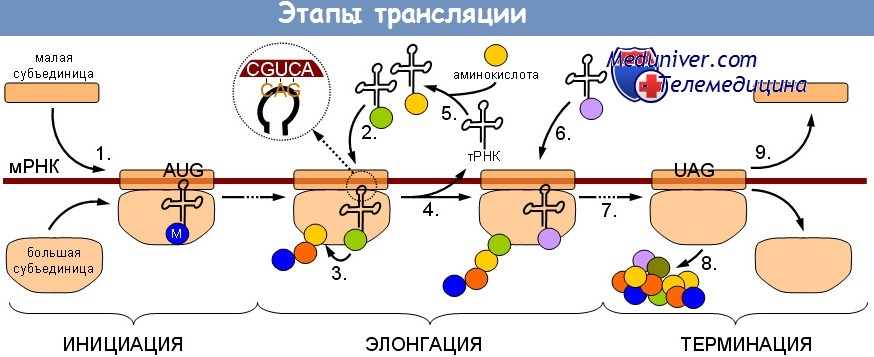
Трансляция мРНК является высокоорганизованным и многокомпонентным сигнальным путем, который можно разделить на три стадии:
(1) инициацию;
(2) элонгацию;
(3) терминацию.
На первоначальной стадии небольшие (40S) рибосомные субъединицы вовлекаются в отобранную мРНК и вместе с большой (60S) рибосомной субъединицей формируют рибосому 80S, которая способна распознать начало трансляционного кодона и начать процесс элонгации. Процесс элонгации включает энергоемкую процедуру присоединения аминокислот, одной за другой, к удлиняющейся цепочке пептида. Стадия терминации состоит из идентификации стоп-кодона и отделения рибосомных субъединиц от мРНК.
Каждая из этих стадий регулируется отдельными категориями белковых факторов, называемых соответственно стадиям эукариотическими факторами инициации (eIF), эукариотическими факторами элонгации (eEF) и эукариотическими факторами высвобождения (eRF). Большинство этих белковых факторов, регулирующих трансляцию, имеют несколько субъединиц и содержат связывающие участки для взаимодействия с другими факторами трансляции, а также для объединения с рибосомой. Кроме того, некоторые из них способны проявлять каталитическую активность, которая может быть использована для стимулирования или ингибирования трансляции.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Репродукция вируса – это процесс размножения вирусных частиц в чувствительных к ним клетках. Репродуцируются только вирулентные вирусы, обладающие высокой степенью патогенности [3] .
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций [3] .
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина [2] .
- Все вирусы, содержащие молекулу ДНК, кроме вирусаоспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирусоспы размножается только в цитоплазме клетки [2] .
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами [2] .
- Белки вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению) [2] .
- Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование (присоединении к полипептиду углеводных остатков) [2] .

Этапы репродукции (жизненного цикла) вируса

1. Адсорбция вируса на мембране клетки.; 2. Проникновение вируса в клетку.; 3. Депротеинизация.; 4. Синтез компонентов вирусов.; 5. Формирование дочерних вирионов.; 6. Выход вирионов [2] .
Этапы репродукции вируса
Репродукцию или жизненный цикл вируса делят на шесть последовательных этапов:
- Адсорбция на мембране клетки [2] .
- Проникновение в клетку [2] .
- Депротеинизация [2] .
- Синтез компонентов вирусов[2] .
- Формирование дочерних вирионов[2] .
- Выход вирионов[2] .

Процесс адсорбции вириона на поверхности клетки

1. Вирион.; 2. Клеточный рецептор.; 3. Прикрепительный белок; 4. Клеточная мембрана [2] .
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона [2] .
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур [2] .
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно [2] .
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 [2] .

Проникновение вируса в клетку (Путь I)

Слияние вирусной оболочки с клеточной мембраной
1. Вирион.; 2. Инфицируемая клетка.; 3. Ядро [2] .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона [2] . Существует два пути:
- Путь I – слиянии вирусной оболочки с клеточной мембраной [2] .
- Путь II – рецептор-опосредованный эндоцитоз [2] .
Путем слияния суперкапсида с клеточной мембраной (путь I) в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки [2] .
Путем рецептор-опосредованного эндоцитоза (путь II) в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении, вирусный геном в цитоплазме клетки освобождается из эндосомы [2] .

Проникновение вируса в клетку (Путь II)

1. Вирион.; 2. Клетка.; 3. Эндосома.; 4. Ядро [2] .
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы) [2] .
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона [2] .
Синтез компонентов вируса
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [1] .
Место синтеза компонентов дочерних вирионов зависит от типа генома:
- реализация генетической информации у ДНК-содержащих вирусов идет по пути: ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом идет по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок;
- РНК-содержащие ретровирусы идут по следующему пути передачи информации: РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок [2] .
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона [2] .
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках [2] .
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК [2] .
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается [2] .
РНК-содержащие ретровирусы отличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза [2] [4] .
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков, и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры [2] .
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более [2] .
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования[2] .
Путь лизиса клетки тесно связан ее деструкцией. Он характерен для безоболочечных вирусов, не имеющих суперкапсидной оболочки (суперкапсида) [2] .
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы, высвобождаются в течении 2–6 часов. В начале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида [2] .
Читайте также: