
Этапы взаимодействия вирусов с чувствительными клетками и факторы способные их нарушить
Обновлено: 25.04.2024
Вирусные частицы, или вирионы являются инертными, статическими формами вируса. Когда вирионы находятся вне клетки, они не репродуцируются. В настоящее время известно три типа взаимодействия вирусов с клетками хозяина.
Продуктивный тип взаимодействия заканчивается образованием вирусного потомства.
Абортивный тип не завершается образованием вирусных частиц, поскольку инфекционный процесс прерывается и не сопровождается формированием инфекционного потомства.
Интегративный тип взаимодействия вирусов с клеткой характерен для онкогенных вирусов, нуклеиновая кислота которых способна встраиваться (интегрироваться) в клеточную хромосому, вызывая трансформацию клеток. Границы между вирусами с автономной репликацией геномов и интеграционными вирусами довольно условны, и один и тот же вирус, в зависимости от вида клеток, может вести себя либо как инфекционный, либо как интеграционный геном. Результатом такого взаимодействия вируса и клетки является изменение наследственных свойств клетки. Данный тип взаимодействия вируса и клетки называют вироге-
нией, подобно лизогении при взаимодействии фагов с бактериями. Вирусы, способные вызывать вирогению, относят к группе умеренных.
Продуктивный тип взаимодействия вируса и клетки получил название репродукции вирусов (от англ. rерroduсе — воспроизводить). Репродукция вирусов — это образование по принципу комплементарности и путем репликации копий вирусных нуклеиновых кислот и индуцирование молекулами последних биосинтеза вирусных белков с последующей самоорганизацией этих компонентов в вирусные частицы.
Синтез и репликацию нуклеиновых кислот вирусов осуществляют ферменты. Используя нуклеотиды клетки, ферменты создают из них полинуклеотидные цепи новых молекул нуклеиновых. кислот вирусов. В зависимости от типа синтезируемых нуклеиновых кислот они называются ДНК-полимеразами или РНК-синтетазами. В некоторых случаях нуклеиновые кислоты вирусов реплицируются клеточными полимеразами — ферментами, присутствующими в клетке до ее заражения вирусом, хотя чаще репликацию вирусных нуклеиновых кислот осуществляют полимеразы, появляющиеся после заражения клетки вирусом. Такие полимеразы называют вирусспецифическими, так как биосинтез их закодирован в структуре нуклеиновых кислот самих вирусов. И, наконец, репликация нуклеиновых кислот некоторых вирусов происходит за счет предсуществующих в вирионе полимераз. Синтез вирусспецифических полимераз и вирусных структурных белков осуществляется на рибосомах клетки. Вирусные полимеразы, например РНК-репликазы, являются строго специфичными.
При большом разнообразии механизмов репродукции вирусов общим для них является то, что источником мономеров для синтеза и репликации нуклеиновых кислот служат нуклеотиды клетки. Источником мономеров для синтеза и построения белков всех вирусов являются аминокислоты, и синтез белков всех вирусов независимо от ультраструктуры их нуклеиновых кислот осуществляется в клеточных рибосомах. Источником энергии для биосинтетических процессов при репродукции вирусов является АТФ, которая вырабатывается в митохондриях клетки-хозяина.
Процесс репродукции вирусов включает шесть этапов: адсорбцию вируса на клетке; проникновение в клетку; депротеинизацию и освобождение вирусного генома; синтез вирусных компонентов в клетке-хозяине; сборку и формирование вирусов; выход зрелых вирусов из клетки.
Адсорбция, то есть прикрепление ви руса к клетке, осуществляется специфическими и неспецифическими механизмами. Неспецифический — определяется силами электростатического
взаимодействия. В этом процессе участвуют положительно заряженные аминные группы вирусного белка и кислые фосфатные группы клеточной поверхности, имеющие отрицательный заряд. Специфический механизм взаимодействия вируса и клетки обусловлен комплементарными клеточными и вирусными рецепторами. Вирусные рецепторы подразделяют на липопротеиновые (у арбовирусов) и мукопротеиновые (у миксовирусов и аденовирусов). Спектр чувствительности клеток к вирусам часто определяется наличием соответствующих рецепторов.

Резистентность клеток можно преодолеть путем разрушения клеточной мембраны, для чего используют инактивированный вирус Сендай. Специфические противовирусные антитела и антитела к нормальным клеткам препятствуют адсорбции вирусов. Процесс адсорбции состоит из двух периодов — обратимого и необратимого. Период обратимой адсорбции может закончиться десорбцией вируса. При длительном контакте клеток и вируса наступает стадия необратимой адсорбции. Адсорбированные вирионы таким образом могут частично элюироваться с поверхности клеток, небольшая часть остается интактной, а основная масса вирусных частиц проникает в клетку.
Проникновение вирусов в клетку осуществляется путем ви-ропексиса, или пиноцитоза. При этом в месте адсорбции вириона происходит сначала инвагинация наружной мембраны клетки и образование внутриклеточной вакуоли с вирусной частицей. Через некоторое время вирусная и клеточная мембраны вакуоли лизируются и высвобождается нуклеокапсид вируса. Другие вирусы, например вирус Неrреs simр1ех, проникают в клетку не путем виропексиса, а расплавления мембран вируса и клетки. После расплавления вирусный нуклеопротеид оказывается в цитоплазме. Большинство вирусов проникают в клетку путем виропексиса, меньшинство видов — путем сплавления. У фагов этот процесс, то есть освобождение ДНК и ее последующая инъекция сквозь оболочку бактериальной клетки, происходит непосредственно на поверхности этой клетки сразу же после прикрепления к ней фага.
Как только вирусный геном освобождается от белка, вирусная нуклеиновая кислота дезорганизует работу клеточных систем. Вирус стимулирует синтез ингибитора клеточных РНК. Ингиби-
Реализация генетической информации вирусов осуществляется в соответствии с процессами транскрипции — синтеза информационных РНК, комплементарных матричным ДНК или РНК; трансляции — синтеза белков на рибосомах клетки с участием и-РНК; репликации — синтеза молекул нуклеиновой кислоты, гомологичных геному (рис. 7).
| Синтез вирусных ДНК у ДНК-содержащих вирусов осуществляется с помощью ДНК-полимераз. Благодаря участию этого фермента из нуклеотидов клетки синтезируется и строится вторая комплементарная нить ДНК, в результате чего образуются новые двухцепочечные молекулы ДНК. Процесс репликации молекул ДНК продолжается до тех пор, пока в клетке не накопится определенное их количество, необходимое для потомства вирусной частицы, проникшей в клетку.
Поскольку типы и формы нуклеиновых кислот разнообразны — кроме двухцепочечной ДНК могут быть РНК, одно- и двухцепочечные, линейные и кольцевые молекулы — то и механизмы их репликации различны.
| Вирусные белки в т. ч. ферменты |
| ОАО |
| Новые молекулы вирусной ДНК |
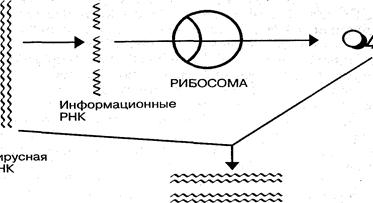
Рис. 7. Схема репликациодвухцепочечных молекул вирусных ДНК и передачи генетической информации в рибосомы
| И-РНК(+) |
| Вирусные белки в т. ч. РНК-репликаза |
| Транскриптаза ———————^ Вирусная РНК |
 |
| ОАО |
| Новые молекулы вирусных(-) РНК |
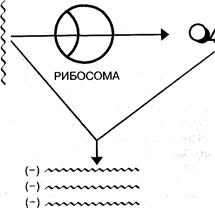
Рис. 8. Схема репликации вирусных минус-РНК и передачи генетической информации в рибосомы
у которых РНК обладает информационными свойствами и направляет синтез специфических белков (пикорна- и тогавирусы);
вирусы, у которыхРНК не является информационной, а служит только матрицей для синтезаи-РНК (ортомиксо-, парамиксо-, рабдовирусы) (рис. 8).
Особым способом репликации отличаются РНК-содержащие двухцепочечные ретровирусы. Существенное отличие состоит в том, что у них имеется фермент — РНК-зависимая-ДНК-полимераза, с помощью которой на матрице вирионной РНК синтезируется ДНК-провирус кольцевой формы. Этот провирус встраивается в клеточную хромосому и транскрибируется клеточной РНК-полимеразой точно так же, как и обычные клеточные гены. Образующаяся РНК направляет синтез вирусных белков и одновременно является вирионной РНК. Вирусная нуклеиновая кислота кодирует синтез двух классов белков: неструктурных белков — ферментов, которые обеспечивают процесс репродукции вирусов на разных его этапах, и структурных белков, которые войдут в состав вирионов потомства. Наряду с синтезом белков в клетке при репродукции вирусов, имеющих суперкапсид, например, вируса гриппа, происходит синтез углеводных цепей, входящих в состав гликопротеидов. Присоединение углеводов осуществляется с помощью клеточных ферментов трансфераз, синтез липидов суперкапсида — клеткой-хозяином. Вирусный суперкапсид формируется при включении липидов из плазматической мембраны клетки-хозяина. Синтез вирусных нуклеиновых кислот и вирусспецифических белков происходит почти одновре-
менно и не менее чем на час опережает созревание вирусных частиц.
При таком механизмевыхода вирионов из клетки последняя сохраняет в определенной мере свои функции. Так, например, процесс выхода вирусов гриппа может продолжаться более 30 часов. Герпесвирусы могут выходить из клетки через цитоплазматические трубочки, соединяющие ядерную оболочку с наружной мембраной клетки. Благодаря такому механизму выделения эти вирусы передаются от клетки к клетке, не выходя во внешнюю среду. Образовавшиеся в процессе репродукции вирионы могут инфицировать новые клетки и проходить в них новый цикл репродукции.
Интегративный тип взаимодействия характеризуется встраиванием нуклеиновой кислоты вируса в хромосому клетки-хозяина. При этом вирусный геном функционирует как составная часть клеточного генома. Интегративный тип взаимодействия присутствует у бактериофагов, онкогенных вирусов, вируса гепатита В, вирусов герпеса и вируса иммунодефицита человека. В частности,
умеренные фаги вступают в симбиоз с частью бактерий, при этом ДНК фага встраивается в хромосому бактерии. В данном случае геномом фага называют профаг, ставший частью хромосомы бактерии и не вызывающий ее лизиса.
Симбиоз микробной клетки с умеренным фагом называют лизогенией. Самопроизвольно или под действием ультрафиолета, или химических факторов профаг может из хромосомы переходить в цитоплазму и вести себя как вирулентный фаг, лизирующий бактерии.
У онкогенных вирусов и вируса иммунодефицита человека процесс интеграции является обязательным в цикле их репродукции. У этих вирусов на матрице РНК с помощью фермента РНК-зависимой ДНК-полимеразы синтезируется ДНК-копия, которая встраивается в хромосому клетки. ДНК вируса, находящаяся в составе хромосомы клетки, называется ДНК-провирусом. Клетка сохраняет свои функции и при делении передает ДНК-провирус дочерним клеткам.
Таким образом, состояние вирогении наследуется. ДНК-про-вирус несет дополнительную генетическую информацию, поэтому интеграция является причиной некоторых аутоиммунных и хронических заболеваний, опухолей. Под действием некоторых физических и химических факторов ДНК-провирус может вырезаться из клеточной хромосомы и переходить в автономное состояние, включаясь в цикл репродукции.
Вирусы- мельчайшие неклеточные формы жизни; царство Vira. Вирусная частица – вирион, которая включает в себя генетический материал (ДНК/РНК) и нуклеокапсид, защищающий его, не имеют собственной белоксинтезирующей системы, поэтому являются облигатными внутриклеточными паразитами, размножаются в ядре и ЦП клетки.
Особенности размножения (этапы репродукции):
1. Избирательная адсорбция на клеточных рецепторах (прикрепление)
2. Рецептор-опосредованный эндоцитоз:
Взаимодействие с рецепторами – деполимеризация клатрина – образование пиноцитозного пузырька – вход в ЦП
Ранняя эндосома – поздняя эндосома
3. Раздевание (депротеинизация) (у оболочечных+слияние ЦПМ клетки и суперкапсида): ЭКЛИПС
4. Синтез макромолекул:
· Синтез ранних мРНКнеструктурных белков (полимеразы, вирусные протеазы, регуляторы скорости репликации);
Ø -РНК: достройка до +РНК;
Ø Ретротранскрипция: -РНК –ДНК.
· Синтез структурных белков (белки капсида и суперкапсида, ферменты) и поздних мРНК;
· Посттрансляционная модификация белка.
**Возможна интеграция генетического материала вируса в геном хозяина (гепатит В, герпес, ВИЧ)
Провирус – интегрированная в геном клетки вирусная ДНК
5. Сборка вирионов:
· ДНК-вирусы: в ядре;
· РНК-вирусы: в ЦПМ.
Гликопротеины сложных вирусов транспортируются к ЦПМ или накапливаются в ЭПР
6. Выход вирионов:
· Литический тип репродукции: полиомиелит
· Нелитический тип: путем почкования – гепатит В
Классификация
1. По строению:
| Простые вирусы | Сложные вирусы (оболочечные) |
| НК+капсид (белковый футляр)+ферменты и НК-связанные белки = нуклеокапсид | Нуклеокапсид+суперкапсид(мембрана клетки-хозяина с глиекпротеинами вируса) |
| Вирус полиомиелита, аденовирус | Вирус гриппа А, вирус бешенства |
2. По типу НК:
· РНК (80% всех вирусов) (Ретровирусы, Рабдовирусы, Пикорновирусы, Коронавирусы):
Ø +РНК цепь: используется в качестве мРНК и генома (полиовирус);
Ø -РНК цепь: служит геномом, в качестве мРНК – достраивается комплиментраная +РНК цепь (парагрипп);
· ДНК (у многих бактериофагов) (Аденовирусы, Герпесвирусы, Парвовирусы):
Ø Двуцепочечная (герпес);
Ø Одноцепочечная (првовирусы);
Ø Двунитчатая кольцевая (папилломавирус);
Ø Двунитчатая кольцевая с дефектом одной цепи (геатит В)
3. По размеру вирионов:
4. По форме вирионов:
· Спиральная симметрия (Вирус бешенства);
· Икосаэдрическая симметрия (Аденовирус человека).
Тропность к тканям и клеткам.
6. По механизму передачи:
· Аэрогенный (грипп, ротавирус);
· Фекально – оральный (энтеровирусы, ротавирус);
· Трансмиссивный (лихорадка Западного Нила);
· Контактный (оспа, бешенство, ВИЧ, ЦМВ, гепатит).
7. По классу поражаемых хозяев:
Ø Бруцеллез – свиньи;
Ø Сибирская язва – КРС;
Ø Энцефалит - клещи;
Ø Болезнь Лайма;
Ø Чума (легочная);
Вирусологический метод, основные этапы.
Способ обнаружения и идентификации вирусов посредством культивирования; являет собой выделение вируса из патологического материала на восприимчивой живой системе.
Идентификация: противовирусные препараты или сыворотки.
Этапы
1. Подготовка материала (фекалии, носоглоточные смывы, спинномозговая жидкость, кровь, моча, мазок с конъюнктивы):
· Фильтрация через бактериальные фильтры;
· Обработка антимикробными и антигрибковыми препаратами.
2. Подбор модели:
Живые системы.
· Культура клеток (специально культивированные среды):
*Матрас: сосуд, покрытый соединением (коллаген) для заякоривания клеток на стенке, используются высокочистые марки агара.
v Первичные/неперевиваемые: культура каждый раз получается из исходной ткани.
v Полуперевиваемые (эмбриональные клетки – для производства вакцин), не способы к злокачественному перерождению; способны совершить до 50 делений (лимит Хейфлика); образуют монослой клеток.
· Эмбрионы птиц (малоинформативные);
· Лабораторные животные (наблюдение клинических проявлений).
4. Оценка феноменов присутствия вирусов:
· Цветная проба Солка (для суспензионных культур):
· Феномен Дюльбенко (культура на агаре)
Ø Образование синцития – корь;
Ø Окружности вирионов с полостями – полиомиелит;
Ø Группы вирионов между фибробластами– аденовирус;
Ø Гемадсорбция – грипп
5. Титрование вируса
· Прямое обнаружение АГ вируса: РИФ, ИФА, РНГА
Ø РНЦПД (в монослое)
Ø ТельцпБабеша-Негри (гиппокамп): бешенство;
Ø Совиный глаз: ЦМВ;
Ø Пустое ядрышко: аденовирус.
Этапы взаимодействия вирусов с чувствительными клетками и факторы, способные их нарушить.
1. Адсорбция: взаимодействие специфических рецепторов ЦПМ клетки и адгезинов на поверхности вириона.
Поврежденные клетки могут не иметь специфических рецепторов
2. Пенетрация: слияние суперкапсида с мембраной клетки (для сложных вирусов) или клатрин/кавиолин-зависимый эндоцитоз (для простых)
3. Раздевание (освобождение от нуклеокапсида) и активация НК.
4. Синтез НК и вирусных белков
5. Сборка вириона: ассоциация ген. материала и капсидного белка
6. Выход из клетки и приобретение суперкапсида (для сложных)
Формы вирусной инфекции.
По локализации.
1. Очаговая: короткий инкубационный период, вирусы размножаются в органах, не попадая в кровь и лимфу. Характерен короткий и нестойкий иммунитет. В основном – респираторные инфекции.
2. Генерализованная: длительный инкубационный период; возбудители с кровью и лимфой разносятся по всему организму, поражая восприимчивые ткани. Иммунитет – длительный и стойкий.
По длительности.
1. Острое заболевание (часто): ярко выраженные клинические проявления; как правило, завершается освобождение организма от вирусов и выздоровлением, но возможна хронизация при условии некорректного лечения.
2. Инаппарантная форма: атипичная острая инфекция (отсутствуют характерные для острого процесса симптомокомплексы).
3. Медленные инфекции: многомесячный инкубационный период, после которого проявляются симптомы, всегда заканчивается летально (ВИЧ - инфекция, бешенство, проказа).
4. Хроническая форма (персистенция): протекают в течении нескольких месяцев (бруцеллез);
· Ремиссия: улучшение состояния, незначительное проявление либо отсутствие симптомов;
· Рецидив: обострение патологического процесса, ясное проявление клиники.
По числу возбудителей.
1.Моноинфекция: вызвана патогеном одного вида.
2. Смешанная форма: инфицирование происходит несколькими вирусами; при этом возможны их взаимовлияния, проявляющиеся в подавлении или усилении действия возбудителей на организм.
© 2014-2022 — Студопедия.Нет — Информационный студенческий ресурс. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав (0.014)
Каждый конкретный вирус может поражать только конкретный тип (один или несколько) клеток. Это свойство вирусов лежит в основе их строго цитотропизма – способности к репликации в строго определенных клетках и органах макроорганизма.
Это явление обусловлено тем, что только та клетка является чувствительной к определенному вирусу, которая имеет на внешней мембране рецепторы для его адсорбции и, кроме того, обладает ферментами, осуществляющими его депротеинизацию.
Существуют три основных типа взаимодействия вирусов с клеткой.
Продуктивный тип (продуктивная инфекция) заканчивается образованием новых вирионов. При абортивном (абортивная инфекция) происходит прерывание инфекционного процесса в клетке и новые вирионы не образуются. Интегративный тип (интегративная инфекция) характеризуется встраиванием вирусной ДНК в геном клетки и совместной репликацией генома вируса (в виде провируса) с геномом клетки. В последнем случае происходит как бы совместное сосуществование вируса и клетки, такой процесс называется еще вирогенией.
При продуктивной инфекции размножение вирусов в чувствительной клетке осуществляется путем последовательной смены пяти основных этапов: прикрепления вириона к поверхности клетки, проникновения в нее нуклеиновой кислоты вируса (процесс ее освобождения от белоксодержащих оболочек называется депротеинизацией), синтеза компонентов вируса, сборки вирионов (при этом места сборки вирионов могут проявляться в виде внутриклеточных включений), выхода вирионов из клетки.
При этом за прикрепление (адсорбцию) вирусной частицы на клетке отвечают прикрепительные белки, взаимодействующие с рецепторами адсорбции на ее поверхности. Роль таких прикрепительных белков у сложных вирусов играют шипы суперкапсида, а у простых – белки капсида.
В клетку вирус проникает двумя основными путями: виропексисом (рецепторным эндоцитозом) или путем слияния мембран. Причем, один и тот же вирус может использовать для проникновения в клетку обе способа.
При виропексисе место адсорбции вириона впячивается внутрь, пока не сформируется эндоцитозный пузырек, в котором и происходит депротеинизация.
При слиянии мембран внешняя оболочка вируса сливается в клеточной мембраной и сердцевина вируса оказывается непосредственно в цитоплазме клетки, в которой нуклеиновая кислота уже окончательно освобождается от капсидных белков.
Компонентами вируса, синтезируемыми в клетке, являются вирусные белки и нуклеиновая кислота (т.е. вирусный геном).
В процессе синтеза вирусных белков различают синтез ранних белков (до момента репликации вирусной нуклеиновой кислоты) и поздних (синтезируемых после репликации вирусной нуклеиновой кислоты).
О вариантах репликации нуклеиновой кислоты вируса, т.е. множественного репликации его генома, будет сказано ниже.
В местах сборки вирусные частицы могут скапливаться в больших количествах, что, вкупе с реакцией клетки на скопление вирионов, может внешне проявляться в виде внутриклеточных включений.
Готовые вирионы выходят из клетки или путем ее лизиса или посредством почкования от ее мембраны (как внешней, так и, например, мембраны эндоплазматического ретикулума).
У РНК-геномных вирусов возможны три варианта транскрипции вирусного генома.
У РНК-геномных вирусов с негативным геномом вирионная минус-нить служит матрицей для синтеза информационной РНК, на матрице которой собираются вирусные белки.
У РНК-геномных вирусов с позитивным геномом вирионная плюс-нить, после множественного копирования, используется как матрица для синтеза вирусных белков.
У ретровирусов на матрице вирионной РНК собирается ДНК, которая используется как матрица для синтеза информационной РНК, служащей, в свою очередь, матрицей для синтеза вирусных белков.
ДНК-геномные вирусы собираются, как правило, в ядре. В цитоплазме собираются лишь Hepadna- и Poxviridae. Все РНК-геномные вирусы собираются в цитоплазме.
Исходы вирусной инфекции для конкретной клетки зависят от того, в каком состоянии находится в этой клетки вирусная нуклеиновая кислота, т.е., собственно, сам вирус.
Если нуклеиновая кислота клетки интегрирована в ее геном, то вирусная инфекция летки может привести к опухолевому перерождению клетки или развитию латентной инфекции. Такие же последствия для клетки несет нахождение в ней вирусной нуклеиновой кислоты в виде плазмиды.
В случае продуктивной инфекции клетка из нее выходят сформировавшиеся вирионы или путем ее лизиса, или без такового.
1. Адсорбция – пусковой механизм, связанный с прикреплением вириона к поверхности клетки. Выделяют две основных фазы адсорбции:
а) неспецифическая обусловлена ионным притяжением между вирусом и клеткой.
б) высокоспецифическая обусловлена гомологией, комплементарностью специфических рецепторов вируса и хозяина (у вируса гриппа – гемагглютинин к сиаловой кислоте гликопротеинов клеток дыхательных путей, у вируса иммунодефицита человека - гликопротеин gp 120 - к СД4 рецепторам Т-хелперов, моноцитов, макрофагов).
Проникновение
а) путем слияния оболочки вириона с мембраной клетки, характерно для некоторых оболочечных вирусов (парамиксовирусов, ретровирусов, герпесвирусов)
б) путем эндоцитоза (пиноцитоза) в результате захватывания и поглощения вириона клеткой: клеточная мембрана с прикрепленным вирионом впячивается с образованием внутриклеточной вакуоли (эндосомы), содержащей вирус.
3. Освобождение нуклеиновых кислот (депротеинизация) – “раздевание” нуклеокапсида и активация нуклеиновой кислоты. В результате депротеинизации удаляются поверхностные структуры вируса и высвобождается его внутренний компонент, способный вызвать инфекционный процесс.
4. Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса. Синтез нуклеиновых кислот и белков вируса разобщен во времени и пространстве, т.к. осуществляется в разных частях клетки. Такой способ размножения вирусов называется дизъюнктивным.
Синтез белков в клетке осуществляется благодаря процессам транскрипции – переписывания генетической информации с нуклеиновой кислоты в нуклеотидную последовательность информационной РНК (иРНК или мРНК) и трансляции – механизмов, при помощи которых последовательность нуклеотидных оснований мРНК переводится в специфическую последовательность аминокислот в синтезируемом полипептиде на рибосомах клетки хозяина.
5. Сборка вирионов – многоступенчатый процесс, включающий в себя соединение всех компонентов вириона.
Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов. У сложно устроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса). У минус-нитевых РНК вирусов в сборку вовлекается матриксный белок (М-белок), который расположен под модифицированной клеточной мембраной.
6. Выход вирусных частиц из клетки взрывным путем или почкованием, экзоцитозом. Полный цикл репродукции вирусов завершается через 5-6 ч (вирус гриппа) или через несколько суток (вирус кори). По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки. Из погибающей клетки одновременно выходит большое количество вирионов. Почкование, экзоцитоз характерен вирусам, имеющим липопротеиновую оболочку, которая является производной клеточных мембран. Сначала образовавшийся нуклеокапсид транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. Клетка способна длительное время сохранять жизнеспособность и продуцировать вирусное потомство.
Исходы взаимодействия вирусов с клеткой хозяина
1. Продуктивный процесс - репликация (продукция) вирусов. Выделяют:
· продуктивный цитолитический процесс с образованием инфекционного потомства и выходом вирионов во внеклеточную среду.
· продуктивной нецитолитический процесс с образованием инфекционных вирусных частиц без лизиса клетки, которая продолжает функционировать.
Цитопатический эффект является результатом интенсивного размножения и формирования большого количества вирусных частиц (характерный результат продуктивного процесса), вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит узнаваемый специфический характер;
2. Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина (вирогения). Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК - геном хозяина могут только ДНК- вирусы (принцип “ДНК - в ДНК”). Единственные РНК - вирусы, способные интегрироваться в геном клетки хозяина - ретровирусы, имеют для этого специальный механизм. Синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы (ревертазы) с последующим встраиванием ДНК в геном хозяина.
3. Абортивный процесс - клетки освобождаются от вируса, не вызывая появление дочерней популяции или появление в меньшем количестве. При инфицировании дефектным вирусом, для репликации которого нужен вирус- помощник, самостоятельная репликация этих вирусов невозможна. Например, дельта вирус или вирус гепатита D может реплицироваться в клетке только при наличии вируса гепатита B, его HВs – антигена. При инфицировании вирусом генетически нечувствительных к нему клеток или при заражении чувствительных клеток вирусом в неблагоприятных условиях.
Возможные последствия инфекционного процесса, вызванного вирусами для клетки:
- сохранение жизнеспособности клетки
- деструкция клетки, возникающая при цитолитической инфекции (цитопатогенное действие вируса - ЦПД)
- образование многоядерных клеток в результате их слияния (симпластообразование)
- образование вирусных внутриклеточных включений
· онкогенная трансформация клетки при интеграции вирусного генома с геномом клетки (вирогении, интегративной инфекции)
Возможность взаимодействия вируса и клетки обусловлены генетическими особенностями вируса и клетки. Условия среды определяют исход этого взаимодействия.
1) Продуктивная вирусная инфекция
Клетка погибает, происходит репродукция вирусов;
2) Абортивная вирусная инфекция
Клетка не погибает, репродукции вирусов нет;
3) Латентная вирусная инфекция
репродукция вирусов и сохранение жизнеспособности клетки; органы и ткани, построенные из таких клеток, сохраняют свою функциональную активность;
4) Вирус-индуцированные трансформации
Клетки, инфицированные вирусом, приобретают новые, ранее не присущие им свойства; происходит репродукция вирусов.
Продуктивная вирусная инфекция
Лежит в основе развития острых вирусных инфекций. Сопровождается гибелью клетки и репродукцией вирусов. Условно делится на периоды→фазы→этапы.
I. Начальный период
1. Фаза адсорбции
а) неспецифическая адсорбция
действуют силы межмолекулярного взаимодействия (электростатические силы, ван-дер-ваальсовы силы); осуществляется за счет сродства химических группировок; она непрочная, непродолжительная; если смесь вируса и клетки встряхнуть, то эта связь может нарушиться;
б) специфическая адсорбция
в основе – химическое сродство рецепторных белков вириона и рецепторов соответствующей клетки;
за счет этих рецепторов вирусы поражают строго определенные клетки (например, вирус гриппа поражает эпителий верхних дыхательных путей).
Таким образом специфическая адсорбция лежит в основе тропизма вирусов.
Рецепторы вирусов настолько специфичны, что они различны даже для близкородственных вирусов.
Факторы, влияющие на адсорбцию:
(количество вирионов на одну чувствительную клетку);
(в среде, богатой электролитами, адсорбция идет эффективнее);
– суспензия или пласт клеток и т.п.
(в суспензии адсорбция идет лучше, так как больше площадь взаимодействия);
(при снижении температуры адсорбция снижается, при повышении температуры адсорбция повышается);
(антитела, лекарственные вещества и т.п. угнетают развитие адсорбции);
(в условиях макроорганизма;
например: гормон паращитовидной железы снижает эффективность адсорбции, гормон щитовидной железы – повышает).
Процесс адсорбции протекает за короткий промежуток времени. Через 15-20 минут она становится специфичной.
2. Фаза проникновения вирионов в клетку
Первый способ проникновения самый простой и наиболее частый – виропексис (фагоцитоз) – в клетках, обладающих фагоцитарной активностью: макрофаги, нейтрофильные лейкоциты.
На месте адсорбции вириона образуется инвагинация→края мембраны слипаются → вирион оказывается в клетке, окруженный частью клеточной мембраны. Формируется фагосома, которая сливается с клеточными лизосомами и формируется фаголизосома, внутри которой находится вирион. Лизосомальные ферменты начинают разрушать белковую оболочку вириона (депротеинизация), что приводит к освобождению нуклеиновой кислоты.
Второй способ проникновения в клетку:
Фаги впрыскивают свою нуклеиновую кислоту в клетку;
Третий способ проникновения:
Некоторые вирусы на своей поверхности имеют ферменты, расщепляющие компоненты клеточной стенки (нейраминидаза вируса) → образуется отверстие, через которое нуклеиновая кислота попадает в клетку.
3. Фаза депротеинизации
Не всегда эти процессы идут в такой последовательности. Например, у сложных вирусов депротеинизация начинается с момента адсорбции.
Если клетка разрушает вирусную нуклеиновую кислоту → сохранение клетки – абортивная вирусная инфекция.
II. Средний период
РВБ – белки, которые обеспечивают подавление клеточного генома.
2. Фаза репликации вирусной нуклеиновой кислоты
нити расплетаются → на матрице каждой нити формируются дочерние молекулы двуцепочечной вирусной ДНК с участием клеточной ДНК-зависимой ДНК-полимеразы.
синтез дочерних молекул на материнской ДНК происходить не может (т.к. способна синтезироваться только комплементарная ДНК). Поэтому на матрице материнской ДНК сначала синтезируется комплементарная нить (репликативная форма), которая служит основой для синтеза дочерних ДНК.
Вирусная РНК может быть в двух формах:
Ретровирусы – РНК-вирусы. У них реализация генетической информации идет по схеме РНК → ДНК → РНК → белок и осуществляется при участии уникального фермента обратной транскриптазы (ревертаза, РНК-зависимая ДНК-полимераза).
3. Фаза синтеза вирусных белков
т.к. клеточный геном подавлен, клетка начинает воспроизводить вирусные белки. Репликация нуклеиновой кислоты происходит в ядре клетки, а синтез белка на рибосомах. Первый процесс предшествует второму, т.е. они разобщены во времени и пространстве – дисъюнктивный способ репродукции вирусов.
III. Заключительный период
Фаза 1. Сборка новых вирионов
Фаза 2. Выход из клетки
Вновь собранные вирионы покидают инфицированную клетку.
1. Клеточный геном погиб → гибель и разрушение клетки → вирионы выходят в межклеточное пространство;
2. Механизм обратного виропексиса (≈экзоцитоз)
Новые вирионы подходят к клеточной мембране → вытягивание → разрыв мебраны. В ходе этого процесса новые вирионы могут включить в состав своих суперкапсидных структур элементы клетки-хозяина.
Латентные вирусные инфекции - лежат в основе медленных вирусных инфекций. Это большая группа заболеваний человека и животных. Они протекают длительно, десятилетиями, без клинических проявлений, но неизбежно прогрессируют и заканчиваются летально. К ним относятся рассеянный склероз, амиотрофический склероз, прогрессирующий склерозирующий панэнцефалит. И другие.
Два типа латентных вирусных инфекций:
- безусловная (абсолютная) – все клетки органа или ткани поражены вирусом. Выделить клон свободных от вирусов клеток не удается. Но в пораженных клетках клеточный и вирусный геномы сосуществуют: 95-98% синтеза в такой клетке детерминированы клеточным геномом, а 2-5% - вирусным. Однако это соотношение непостоянно и может сдвигаться в пользу вирусных и тогда – клиническое проявление. А т.к. поражены все клетки органа и ткани, то они все перестают функционировать. Как они сосуществуют? Если не вся, то часть ДНК вируса встраивается в геном клетки. Если РНК, то она существует наряду с другими РНК этой клетки.
- условная или относительная – вирусом поражены единичные клетки ткани или органа, в них инфекция протекает по продуктиному типу, но т.к. поражены единичные клетки, то ткань или орган в целом сохраняют свою функцию. Почему поражаются отдельные клетки? Различная клональная структурная организация. Невысокая урожайность. Существует целый ряд ингибиторов, которые препятствуют распространению вирусной инфекции на все клетки.
Такой тип взаимодействия может быть нарушен на любом этапе до фазы смены информации → формируется абортивная вирусная инфекция.
Читайте также: