
Генетика человека и гепатит в
Обновлено: 25.04.2024
На основании филогенетического анализа нуклеотидных последовательностей полного генома ВГВ подразделяют на 10 генотипов, обозначающихся латинскими буквами от A до J. Для каждого генотипа характерна определенная географическая и этническая зона распространенности. Генотип A превалирует в Северной Америке, Западной Европе и Центральной Африке. Генотипы B и C характерны для Китая и стран Юго-Восточной Азии. Генотип D доминирует в странах Восточной Европы, Средиземноморья и Индии, генотип E – в Западной Африке, генотип F – в Южной Америке и на Аляске, а генотип H – среди жителей Центральной Америки. Генотипы I и J выявлены в Юго-Восточной Азии. Распространенность генотипа G изучена недостаточно.
В ходе исследований, проведенных в Центральном НИИ эпидемиологии Роспотребнадзора, на территории Российской Федерации была выявлена циркуляция трех генотипов ВГВ – D, A и С. Генотип D доминирует на территории РФ. Вторым по встречаемости является генотип А, причем в Европейской части России его доля составляет 5–10%, в Сибирском регионе – в среднем 10–15%, достигая максимума (почти 50%) в Республике Саха (Якутия). Генотип С является эндемичным для коренного населения Чукотского АО, где его доля достигает 25%. В остальных регионах РФ крайне редко регистрируются единичные и, как правило, завозные случаи инфекции, вызванные генотипом С ВГВ.
Заболевания, вызванные различными генотипами ВГВ, могут отличаться по клиническому течению и исходам. ГВ, вызванный вирусом генотипа С, чаще принимает хроническое течение и имеет больший риск трансформации в цирроз печени или гепатоцеллюлярную карциному по сравнению с гепатитами, вызванными другими генотипами ВГВ.
Генотип ВГВ может влиять на эффективность лечения ХГВ препаратами интерферона. Пациенты, инфицированные вирусом генотипа A, значительно лучше отвечают на лечение препаратами интерферона по сравнению с пациентами, инфицированными другими генотипами ВГВ. Связь между эффективностью лечения ХГВ аналогами нуклеозидов/нуклеотидов и генотипом ВГВ не установлена.
Определение генотипа ВГВ способствует выявлению завозных случаев инфекции, определению прогноза течения и исхода заболевания ГВ и выбору оптимальной тактики лечения ХГВ.
Показания к обследованию. Больные ХГВ
Методы лабораторных исследований
- ПЦР;
- обратная гибридизация с зондами на мембране (LiPA);
- прямое секвенирование.
Материал для исследования. Плазма или сыворотка крови.
Особенности интерпретации результатов лабораторных исследований. Генотип ВГВ, в совокупности с другими клинико-лабораторными данными, учитывается при определении тактики лечения.
Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.
Вирус гепатита В не способен размножаться in vitro. Некоторые этапы его репродукции остаются плохо изученными. Геном представлен двухцепочечной молекулой ДНК, но плюс-нить несколько короче, каждая нить имеет на концах одинаковые двойные сегменты, обеспечивающие интеграцию в геном клетки. Вирионы содержат полимеразу, способную транскрибировать ДНК в РНК.
После раздевания вирусные ДНК и полимераза мигрируют в ядро, где под действием последней осуществляется дополнение плюс-цепи ДНК (в качестве матрицы используется минус-цепь). Полная двухцепочечная ДНК транскрибируется с образованием небольших молекул мРНК и полной молекулы +РНК (эта прегеномная РНК служит матрицей для репликации генома). Синтез различных вирусных ферментов и белков реализуется на небольших молекулах мРНК. Репликация генома осуществляется через ряд, последовательных процессов.
• Прегеномная +РНК взаимодействует с вирусной полимеразой. Фермент действует как обратная транскриптаза и синтезирует -ДНК, используя +РНК как матрицу (позднее она разрушается).
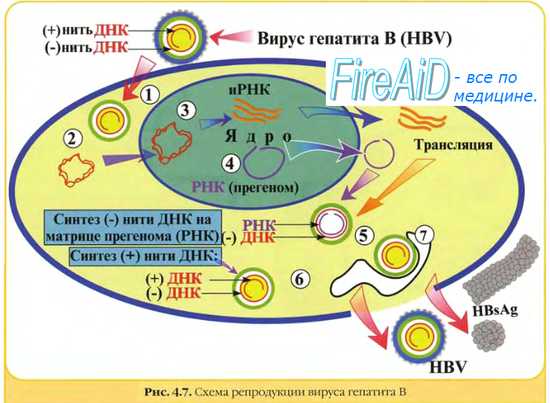
• Затем -ДНК используется как матрица для транскрипции +ДНК, но этот процесс носит незавершённый характер и заканчивается образованием двойной молекулы с неполной плюс-цепью.
Сборка вирионов вируса гепатита В заканчивается формированием оболочки, включающей специфический поверхностный Аг (HBsAg).
Интеграция вируса гепатита В. Одновременно с реализацией сложного репродуктивного цикла вирусная ДНК может интегрироваться в геном клетки путём расщепления на множество фрагментов, встраивающихся в различные участки клеточной ДНК. После встраивания доступным для считывания остаётся лишь фрагмент, кодирующий HBsAg, что даёт ему возможность реплицироваться даже при отсутствии возможности для полной репродукции вируса.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Мутации вируса гепатита В. Генетическая изменчивость ВГВ
Мутантом (от лат. mutare — меняться) называют ген, хромосому, организм или популяцию, отличающуюся от соответствующего дикого типа одним или более генетическим изменением — мутацией. Мутация — это относительно устойчивое и передающееся по наследству изменение генетического материала, представляющее собой биохимическое изменение в кодоне или физическое изменение в межхромосомных взаимоотношениях. Мутации HBV в современной научной литературе описывают довольно часто.
Установлено, что HBV подвержен более частым мутациям, чем предполагалось ранее, поскольку для его репликации необходима обратная транскрипция РНК-прегенома — процесс, при котором вероятность ошибок считывания весьма высока. Эта вероятность, как установлено, составляет 2x10-4 замен оснований на сайт в год, то есть на 4 порядка превышает частоту мутаций других ДНК-вирусов, хотя и не достигает частоты мутаций РНК вирусов.
Наличие длинных участков делеций свидетельствует о присутствии и других механизмов мутагенеза. Это могут быть ошибки считывания на других этапах репликации, действие клеточной топоизомеразы и сплайсинг генов с альтернативными донорными и акцепторными сайтами. Мутации могут возникнуть также в результате рекомбинации между коинфицирующими штаммами. Мутации HBV не ограничиваются отдельными ОРС и происходят во всех генах вируса и регуляторных элементах.
Среду мутантов HBV, выделяемых от пациентов как с острой (фульминантной), так и хронической инфекцией, различают 4 группы: мутанты генов pre-сorе и core, мутанты генов preS и S (поверхностных) белков, мутанты генов полимеразы/обратной транскриптазы и мутанты гена X. Нередко у одного пациента мутанты сосуществуют с вирусами дикого типа. Фактически слугаи инфекции с угастием мутантных штаммов вируса следует рассматривать как микст-инфекцию с разными исходными ДНК, взаимодействующими друг с другом путем trans-активации и рекомбинации.
Впервые мутанты HBV обнаружены у пациентов с хроническим гепатитом В, в крови которых выявляли вирусную ДНК на фоне отсутствия серологических признаков текущей или перенесенной инфекции (т. е. HBsAg и анти-HBs). Поскольку HBV может персистировать в организме в течение нескольких лет и даже десятилетий, мутации способны накапливаться и становиться клинически значимыми.
Точечные мутации в S-гене представляют наибольший интерес для практического здравоохранения, поскольку они способны влиять на иммуногенность HBsAg и, особенно, его детерминанты а, к которой вырабатываются протективные антитела. Для того чтобы объяснить, как мутация в одном участке гена может влиять на структуру двух или более удаленных друг от друга эпитопов, была предложена новая модель большого гидрофильного региона (БГР) S-белка.
Исходная модель детерминанты а белка S предполагала наличие двухпетлевой структуры в районе аминокислотных остатков 124-147 с дисульфидными мостиками между аминокислотными остатками 124 и 137. В соответствии с вновь предложенной моделью (36), которая не отрицает возможного наличия дисульфидных мостиков, большой гидрофильный регион белка S (аминокислотные остатки 100-160 или 169) рассматривают как паутиноподобную структуру, образованную цистеиновыми остатками в позициях 107, 137, 138, 139 и 149 и локализованную на наружной оболочке вириона. Пептидные петли, образованные аминокислотными остатками 107-137 и 139-147, формируют выступы на оболочке вириона, а еще одна плотная петля образована аминокислотными остатками 121-124.
Геном вирусного гепатита В. Профилактика вирусного гепатита В.
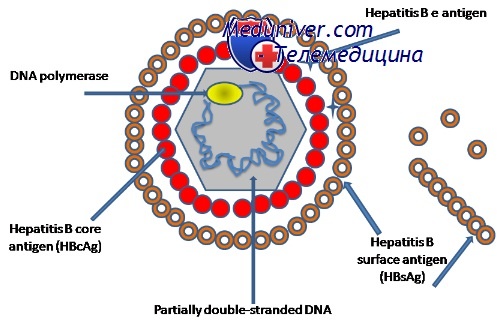
Геном вирусного гепатита В представлен циркулярной, частично двуцепочечной разновидностью ДНК (3,2 тн), циркулярность которой поддерживается связью 5'-концов. Структура генома отличается тем, что две цепи ДНК не вполне симметричны. Негативная цепь имеет полноразмерную длину, и ее 5'-конец ковалентно связан с белком. Позитивная цепь имеет меньшую длину и олигонуклеотидную последовательность (кэп-структуру на 5'-конце). Таким образом, вирионная ДНК имеет односпиральный участок (пробел) фиксированной полярности, но различной длины. Кодирующая организация вирионной ДНК характеризуется высокой компактностью: каждый нуклеотид генома находится внутри кодирующей области и более половины последовательностей транслируется более чем с одной из четырех рамок считывания. Вирусная полимераза, играющая центральную роль в репликации генома, а также концевой белок негативной цепи ДНК кодируются Р геном. С-область генома кодирует структурный белок нуклеокапсида, а S-область — поверхностные гликопротеины. Одной из особенностей структуры ДНК ВГБ является наличие открытой рамки считывания X, продуктом считывания которой является комплекс регуляторных белков, необходимых для проявления инфекционности in vivo и модификации экспрессии гетерологичных и гомологичных генов, по крайней мере в культуре клеток.
Субгеномные транскрипты функционируют исключительно как мРНК для трансляции белков оболочки пре-Sl, npe-S2 и S (соответственно L, М и S белки) и X белка. Субвирусные частицы содержат преимущественно S белок и различное количество М белка, и следовые количества L белка. Частицы Дейна содержат относительно большое количество L белка. Соотношение субъединиц L к М к S выражалось примерно как 1:1:4. Вирусы гепатита млекопитающих почти идентичны по структуре вирионов и генома со езначительной ДНК-гомологией (около 60%). Вирусы гепатита В пекинских уток и лесных североамериканских сурков отличаются от других гепаднавирусов отсутствием гена X, и они кодируют только 2 белка оболочки L и S.

Парадокс состоит в том, что естественная гепаднавирусная инфекция, сопровождавшаяся массовой продукцией высокоиммуногенных частиц, способных вызывать образование ВН-антител в инфицированном организме, не завершается быстрым выздоровлением. Данное явление, вероятно, обусловлено тем, что избыточное количество частиц, содержащих HBsAg, адсорбирует ВН-антитела и тем самым помогает вирусу ускользнуть от хозяинной зависимости. Однако образование антител ограничивает распространение вируса и в дальнейшем ведет к удалению циркулирующего вируса из организма.
Образование ВН-антител к белкам оболочки зависит от Т-лимфоцитов. У некоторых людей ВН-антитела к белкам оболочки вируса не образуются, и развивается персистентная инфекция. Антительный ответ на белки сердцевины ви-риона зависит и не зависит от Т-клеточного ответа. Ответ на HBcAg является первым серологическим доказательством иммунного ответа, который сохраняется годами или даже всю жизнь. Антитела к HBeAg часто сопровождает выздоровление от острого течения заболевания.
Для специфической профилактики гепатита В первоначально использовали плазму крови инфицированных пациентов. Сыворотку крови, содержащую ВГБ (107,5 ИД для шимпанзе), разводили 1:10 и нагревали при 98°С в течение 1 минуты. Вирус инактивировали формолгликолем или формалином и нагреванием. В 1981 г. в США лицензирована вакцина из высокоочищенных инактивированных формалином субвирусных частиц (HBsAg) с использованием в качестве адьюванта гидрата окиси алюминия (ГОА). HBs-антиген не содержал обнаруживаемых количеств нуклеиновой кислоты. С конца 80-х годов прошлого века Hepatovax-B в США заменена рекомбинантной дрожжевой вакциной — Recombivax-HB. HBs антиген не содержал ДНК дрожжей и содержал менее 2% белка дрожжей. Перед адсорбцией на ГОА антиген инактивировали формальдегидом. В отличие от вакцины из плазмы крови, вирусный антиген не был гликозилированным, но вызывал защитный эффект у вакцинированных людей. Трехкратное введение (0, 7 и 21 дни) вакцины Engerix с бустеризацией через 12 месяцев обеспечивает наступление иммунитета в течение трех недель продолжительностью 5 лет. Аналогичная вакцина лицензирована и используется во многих странах. Кроме дрожжей, для изготовления рекомбинантной вакцины используют линию клеток яичника китайского хомяка (СНО).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Вирус гепатита В сопровождал человечество с древности и, вероятно, распространялся вместе со степняками. Об этом сообщили две группы ученых, опубликовавшие результаты генетических исследований древних евразийцев в Nature и на BioRxiv. Одна группа ученых предположила, вирус циркулировал среди европейцев как минимум семь тысяч лет назад, другая группа посчитала, что современные генотипы вируса появились в период 20,4–8,6 тысяч лет назад.
По оценкам ВОЗ, в мире гепатитом В заражены более 250 миллионов человек. В 2015 году от болезни или ее осложнений умерли почти 900 тысяч человек. Передается заболевание через кровь или выделения зараженного человека. Несмотря на то, что вирус гепатита В хорошо изучен, его происхождение и эволюция до сих пор остаются предметом дискуссий. Вирус гепатита В человекообразных обезьян, шимпанзе и горилл, филогенетически очень близок человеческому, поэтому по одной из гипотез древние люди впервые заразились гепатитом В в Африке после того, как съели мясо зараженной обезьяны или контактировали с раненым животным. После исхода с континента 60–70 тысяч лет назад, вирус вместе с сапиенсами распространился по планете. По другой гипотезе человеческий и обезьяний вирусы гепатита В эволюционировали параллельно, так как у приматов Старого и Нового света встречаются очень похожие генотипы вируса. Но молекулярный анализ дает противоречивые данные о времени происхождении человеческого вируса гепатита В, по некоторым оценкам, он появился несколько столетий назад. Стоит отметить, что древнейшая на сегодняшний день вирусная ДНК, выделенная из останков мумии XVI века, оказалась очень похожа на геном современного вируса гепатита В. Почти полное отсутствие генетических изменений за 500 лет кажется очень странным.
Чтобы выяснить происхождение вируса гепатита В и изучить его эволюционную историю, ДНК древнего вируса анализировали сразу две группы ученых. Германские и швейцарские генетики под руководством Йоханнеса Краузе (Johannes Krause) из Института изучения истории человечества Макса Планка в Йене проанализировали геномы трех человек, найденных в Германии. Два из них жили в эпоху неолита (их останки датировались периодом 7056–6959 и 5335–5107 тысяч лет назад, соответственно), третий — в средние века (и умер в 1020–1116 годах). Все трое болели гепатитом В и ученым удалось реконструировать вирусную ДНК. Для сравнения они использовали восемь геномов современного человеческого вируса гепатита В и восемь геномов вирусов приматов.
Результаты показали, что вирус гепатита В циркулировал среди европейцев уже семь тысяч лет назад. Два неолитических генома демонстрировали сходство друг с другом. Но при этом на филогенетическом дереве они занимали промежуточное положение между геномами вируса человека и африканских приматов. Геном средневекового вируса был схож с одним из генотипов современного человеческого вируса гепатита В.
Во второй статье исследователи из 12 стран под руководством Эске Виллерслева (Eske Willerslev) из Копенгагенского университета изучили распространение вируса гепатита В в Евразии за период примерно от 4500 до 800 лет назад. Ученые проанализировали геномы 304 кочевников из Центральной и Западной Евразии, у части из которых обнаружили вирусную ДНК, и отобрали 12 образцов для филогенетического анализа. Для сравнения авторы работы использовали 112 геномов современного вируса гепатита В человека, приматов, грызунов и летучих мышей.
Исследователи построили филогенетическое дерево и рассчитали время возникновения современных генотипов вируса — период с 8,6 до 20,9 тысяч лет назад. Также они выяснили, что генотипы, типичные сейчас для Африки, Центральной Азии и Индии, встречались в Евразии еще в древности и перемещались вместе со степняками во время миграций железного и бронзового веков. Авторы работы предположили, что разнообразие генотипов современного вируса гепатита (сейчас известно девять) возникли приблизительно 25–13,4 тысяч лет назад, после разделения генотипов, встречающихся в Старом и Новом свете.
В последние годы разновидности гепатитов стали одной из ведущих причин смерти на планете и заняли седьмое место среди причин смерти мирового населения. С 1990 по 2013 годы количество смертей от вирусных гепатитов выросло с 900 тысяч до 1,45 миллиона в год.
Читайте также: