
Интегративная вирусная инфекция это
Обновлено: 25.04.2024
Первый вирус – вирус табачной мозаики был открыт русским ученым Ивановским в 1892 г.
Основные отличия вирусов от других форм жизни.
Вирусы представляют собой особую форму жизни, отличную от других следующими основными признаками: вирусная частица содержит только один тип нуклеиновой кислоты, вирусы не имеют клеточного строения, у них отсутствуют белоксинтезирующие и энергозапасающие системы, некоторые вирусы способны интегрироваться в клеточный геном и реплицироваться в его составе синхронно с ним.
Основные признаки, используемые для классификации вирусов.
Вирусы классифицируются по типу нуклеиновой кислоты, структуре генома (количеству нитей нуклеиновой кислоты, его целостности или фрагментированности), наличию суперкапсидной оболочки.
Царство Vira подразделяется на два подцарства: ДНК- и РНК-геномных вирусов. Большинство семейств ДНК-геномных вирусов содержат двухцепочечную ДНК и могут быть как простыми (Adenoviridae, Papillomaviridae, Polyomaviridae), так и сложными (Poxviridae, Herpesviridae, Hepadnaviridae). Два семейства, содержащие одноцепочечную ДНК, относятся к простым вирусам (Parvoviridae и Circinoviridae). Большинство семейств РНК-геномных вирусов содержат одноцепочечную РНК. Подавляющее большинство из них – сложные вирусы, содержащие или позитивный геном (Retroviridae, Togaviridae, Flaviviridae, Coronaviridae) или негативный геном, который, в свою очередь может быть как цельным (Paramyxoviridae, Rhabdoviridae, Filoviridae), так и фрагментированным (Orthomyxoviridae, Bunyaviridae, Arenaviridae). К простым одноцепочечным РНК-геномным вирусам относятся семейства Picornaviridae и Caliciviridae, содержащие позитивный геном. И только вирусы семейства Reoviridae содержат двухцепочечную РНК (фрагментированную), одна цепочка которой представляет собой плюс-цепь, а вторая – минус-цепь.
Близкие к вирусам инфекционные агенты.
В природе существуют инфекционные агенты, к вирусам не относящиеся, но традиционно рассматриваемые в курсе вирусологии. Это вироиды (инфекционные нуклеиновые кислоты, не содержащие белковой оболочки) и прионы (инфекционные белки, не содержащие нуклеиновых кислот).
Принцип строения вириона.
Вирусная частица (вирион) представляет собой нуклеиновую кислоту, окруженную белковой оболочкой (капсидом). У сложных вирусов, в отличие от простых, есть еще дополнительная оболочка, называемая суперкапсидной. Капсид состоит не непосредственно из белковых молекул, а из капсомеров (субъединиц, сформированных несколькими полипептидами). Капсид с заключенной в него нуклеиновой кислотой всегда симметричен. В зависимости от того, как именно уложены в нем капсомеры, различают два основных типа симметрии вирусов: спиральный (капсомеры уложены вдоль спирально закрученной нуклеиновой кислоты) и кубический (капсомеры формируют вокруг нуклеиновой кислоты двадцатигранник – икосаэдр).
Принцип строения суперкапсида.
Суперкапсидная оболочка у всех сложных вирусов построена по одному и тому же принципу: матричный белок примыкает к капсиду, снаружи от него располагается билипидный слой, в который погружены гликозилированные белки, формирующие шипы, отличающиеся у разных вирусов своим внешним видом.
Формы существования вирусов.
Вирион (вирусная частица) – внеклеточная форма существования вирусов. Внутрь клетки проникает только нуклеиновая кислота вирусов, которая – без капсидной и суперкапсидной оболочек – и представляет собой внутриклеточную форму их существования.
Размеры вирусов (точнее, вирионов) – от 15 нм до 400 нм, в зависимости от семейства.
Все простые вирусы имеют форму икосаэдра (двадцатигранника). Большинство сложных вирусов – округлой формы. Лишь три семейства составляют исключение из этого правила: Rhabdoviridae – пулевидной формы, Filoviridae – нитевидной, Poxviridae имеют форму, которую можно охарактеризовать как кирпичеобразную.
Общая характеристика ДНК вирусов.
ДНК в составе вириона может быть или линейной или кольцевой.
Общая характеристика РНК вирусов.
РНК в составе вириона также может быть или линейной или кольцевой. Кроме того, РНК вирусов может быть как цельной, так и фрагментированной. Если вирионная (т.е. содержащаяся в вирионе) РНК может выполнять информационную функцию (т.е. выступать матрицой для синтеза на рибосомах клетки белка), то такая РНК называется плюс-нитью, а вирусы, содержащие ее – вирусами с позитивным геномом. Если вирионная РНК не может выполнять информационную функцию, а выступает лишь как матрица для синтеза информационной РНК, то она называется минус-нитью, а вирусы, ее содержащие – вирусами с негативным геномом.
Общая характеристика белков вирусов.
Белки, входящие в состав вириона, могут быть или структурными (например, белки капсидной оболочки) или функциональными (т.е. вирионными ферментами).
Строгий цитотропизм вирусов.
Каждый конкретный вирус может поражать только конкретный тип (один или несколько) клеток. Это свойство вирусов лежит в основе их строго цитотропизма – способности к репликации в строго определенных клетках и органах макроорганизма.
Типы взаимодействия вирусов с чувствительной клеткой.
Существуют три основных типа взаимодействия вирусов с клеткой. Продуктивный тип (продуктивная инфекция) заканчивается образованием новых вирионов. При абортивном (абортивная инфекция) происходит прерывание инфекционного процесса в клетке и новые вирионы не образуются. Интегративный тип (интегративная инфекция) характеризуется встраиванием вирусной ДНК в геном клетки и совместной репликацией генома вируса (в виде провируса) с геномом клетки. В последнем случае происходит как бы совместное сосуществование вируса и клетки, такой процесс называется еще вирогенией.
Этапы репликации вирусов в чувствительной клетке.
При продуктивной инфекции размножение вирусов в чувствительной клетке осуществляется путем последовательной смены пяти основных этапов: прикрепления вириона к поверхности клетки, проникновения в нее нуклеиновой кислоты вируса (процесс ее освобождения от белоксодержащих оболочек называется депротеинизацией), синтеза компонентов вируса, сборки вирионов (при этом места сборки вирионов могут проявляться в виде внутриклеточных включений), выхода вирионов из клетки.
Место сборки вирионов.
ДНК-геномные вирусы собираются, как правило, в ядре. В цитоплазме собираются лишь Hepadna- и Poxviridae. Все РНК-геномные вирусы собираются в цитоплазме.
Транскрипция вирусного генома.
Генетическая рекомбинация вирусов.
Генетическая реактивация вирусов.
При совместной репликации в одной клетке двух одинаковых вирусов, у которых инактивированы разные части генома, вирусное потомство содержит полностью активированный геном, поскольку одна часть генома (а именно – активированная) берется от одного вируса, а другая (тоже активированная – от другого).
При проникновении в клетку двух вирусов один из них может детерминировать синтез в клетке белка, ответственного за репродукцию другого вируса.
Фенотипическое смешивание вирусов.
Фенотипическим смешиванием называется такое явление, когда – опять же при совместной репликации в одной клетке двух вирусов – вирионы вновь образованных вирусов будут содержать нуклеиновую кислоту одного вируса, а капсидную оболочку – другого.
Патологические процессы, вызываемые вирусами.
Вирусы могут вызывать два основных патологических процесса: инфекционные болезни (вирусные инфекции) и опухоли.
Особенности вирусных инфекций.
В отличие от бактериальных, при вирусных инфекциях возможна интегративная инфекция (вирогения), присутствует стадия вирусемии (кроме тех инфекций, при которых вирус-возбудитель распространяется по организму нейрогенным путем), очень часто вследствие поражения вирусом иммунокомпетентных клеток, развиваются различные иммунопатологические состояния (чаще – иммунодефицит), а некоторые вирусы могут пожизненно персистировать в организме человека.
Исходы интегративной вирусной инфекции.
При интегративной вирусной инфекции нуклеотидные последовательности вируса интегрированы в геном клетки и называются провирусом. Провирус может не транскрибироваться, в этом случае вирусная инфекция внешне никак себя не проявляет – просто все потомство инфицированной клетки содержит в своем геноме провирус. В ряде случаев провирус может транскрибироваться. В этом случае возможны два варианта развития событий. Если провирус транскрибируется частично, то в клетке синтезируется лишь часть вириона. Так, например, клетка, инфицированная вирусом гепатита В, в ряде случае продуцирует лишь суперкапсидную оболочку вириона вируса гепатита В (так называемый HBs-антиген). И, наконец, если провирус транскрибируется полностью, то развивается продуктивная инфекция с синтезом полноценных вирионов.
Исходы активации персистирующего вируса.
В случае активации персистирующего вируса у человека может развиться рецидив заболевания, вызываемого персистрирующим вирусом, или разовьется другое заболевание, вызываемое тем же вирусом, или персистирующий вирус может активировать иной вирус, который вызовет соответствующее (то есть то, этиологическим факторов которого он является) заболевание.
Механизмы опосредования инфекционности вирусов.
Обобщённая схема взаимодействия вирусов с человеческим организмом.
После проникновения вируса в организм человека происходит его репликация в первичном органе-мишени, после чего вирионы попадают в кровь (как правило). Нахождение в крови вирусов называется вирусемией. С кровью вирусы разносятся по организму человека, проникая и реплицируясь во вторичных органах-мишенях. При этом первичные и вторичные органы-мишени чаще всего бывают разными, а клиническая картина вирусной инфекции обуславливается, как правило, поражением именно вторичных органов-мишеней.
Действие факторов противовирусного иммунитета.
Антивирусные антитела действуют только на внеклеточную форму существования вируса – вирион – или до его проникновения в клетку или после его выхода из инфицированной клетки, перед его проникновением в соседние. Антитела действуют и на инфицированную вирусом клетку, активируя на ее поверхности комплемент и обуславливая тем самым ее лизис. Уничтожают инфицированную вирусом клетку так же NK-клетки и цитотоксические лимфоциты (Т-киллеры). Интерферон делает клетки, окружающие поражённую вирусом клетку, нечувствительными к этому вирусу, предотвращая тем самым дальнейшее распространение данного вируса.
Взаимодействие вирусов с фагоцитами.
Роль фагоцитов при вирусных инфекциях неоднозначна. С одной стороны, макрофаги, поглощая вирусы, индуцируют развитие противовирусного иммунного ответа. С другой стороны, макрофаги и другие фагоцитирующие клетки не могут инактивировать поглощенные вирионы и способствуют их распространению по макроорганизму.
Общая классификация препаратов, применяемых для химиотерапии вирусных инфекций.
Все химиотерапевтические средства, которые применяют при вирусных инфекциях, можно классифицировать на четыре основных группы: этиотропные препараты (действующие на сам вирус), иммуномодулирующие препараты (неспецифически стимулирующие противовирусный иммунитет), патогенетические препараты (действующие на то или иное патогенетическое звено конкретной вирусной инфекции), симптоматические препараты (подавляющие проявления того или иного симптома вирусной инфекции).
Препараты для иммунопрофилактики вирусных инфекций.
В настоящее время с помощью вакцинации можно предотвратить развитие желтой лихорадки, натуральной оспы, бешенства, кори, краснухи, эпидемического паротита (для иммунопрофилактики этих инфекций используются живые вакцины), полиомиелита (используется как живая, так и убитая – инактивированная – вакцина), клещевого энцефалита, гепатита А (инактивированные вакцины), гепатита В (генно-инженерная вакцина).
Препараты для иммунотерапии вирусных инфекций.
Для иммунотерапии вирусных инфекций применяют иммуноглобулины, интерферон и интерфероногены (препараты, индуцирующие синтез эндогенного интерферона).
Методы лабораторной диагностики вирусных инфекций.
Для лабораторной диагностики вирусных инфекций используются цитологический (точнее – цитоскопический), вирусологический, серологический (как правило, метод парных сывороток для выявления нарастания титра специфических антител) и молекулярно-генетический (ПЦР) методы.
Общая схема вирусологического метода.
Вирусологический метод заключается в выделении вируса из патологического материала и его идентификации. Культивировать вирусы можно в курином эмбрионе, культуре клеток, в организме лабораторного животного. Выявление наличие вируса в системе культивирования называется индикацией, определение типа выделенного вируса – идентификацией, которая проводится с помощью серологических реакций.
Использование для вирусологического метода диагностики куриного эмбриона.
Наиболее часто куриный эмбрион заражают патологическим материалом одним из пяти основных способов: на хорион-аллантоисную оболочку, в хорион-аллантоисную полость, в полость желточного мешка, в полость амниона, в тело эмбриона. О репликации вируса в курином эмбрионе свидетельствуют гибель эмбриона, морфологические изменения эмбриона или его оболочек, положительная реакция агглютинации с жидкостью из той полости, в которую производилось заражение. Для идентификации вируса, выделенного путем культивирования в курином эмбрионе, используют реакцию нейтрализации (в том числе ее разновидность – реакцию торможения гемагглютинации), реакцию связывания комплемента.
Использование для вирусологического метода диагностики культур клеток.
Чаще всего для культивирования вирусов используют однослойные перевиваемые культуры клеток. Вирусы, реплицируясь в них, могут оказывать цитопатическое действие – любое изменение клеток монослоя, включая бляшкообразование и цветную пробу. Кроме этого для индикации можно ставить реакцию гемадсорбции, а также РИФ. В последнем случае одновременно с индикацией проводится и идентификация вируса. Кроме РИФ для идентификация вируса, культивируемого в культуре клеток, можно использовать реакцию нейтрализации (в том числе ее разновидность – реакцию торможения гемадсорбции), а также реакцию связывания комплемента.
Использование для вирусологического метода диагностики лабораторного животного.
Вид животного, использующегося для вирусологического метода исследования (диагностики), а также способ его заражения, зависит от вида выделяемого вируса. О репликации вируса в организме животного свидетельствует заболевание или гибель последнего. Для идентификации выделенного вируса используется реакция нейтрализации.
Экспресс-методы диагностики вирусных инфекций.
Экспресс-методы диагностики вирусных инфекций можно условно классифицировать на две группы: малоспецифические и высокоспецифические. К малоспецифическому экспресс-методу диагностики относится цитоскопический, при котором в пораженных клетках выявляют внутриклеточные включения, образующиеся в местах сборки вирионов. Такие включения могут быть как внутриядерные (например, при аденовирусной инфекции), так и цитоплазматические (например, включения Бабеша-Негри при бешенстве). К высокоспецифическому экспресс-методу диагностики вирусных инфекций относится обнаружение пораженных вирусом клеток с помощью серологических реакций с мечеными антителами, которые реагируют со специфическими вирусными антигенами в составе инфицированных клеток.
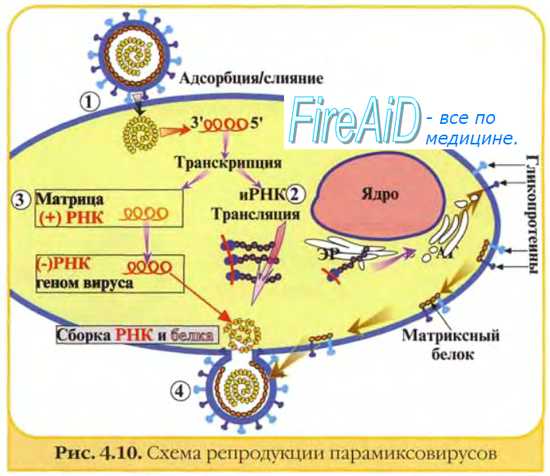
Типы инфицирования клеток вирусами. Репродуктивный цикл вирусов. Основные этапы репродукции вирусов. Адсорбция вириона к клетке.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Репродуктивный цикл вирусов
Изображённые на рис. 2-3 этапы репродукции (от адсорбции вирионов до высвобождения дочерней популяции) происходят при продуктивном взаимодействии вируса с клеткой.
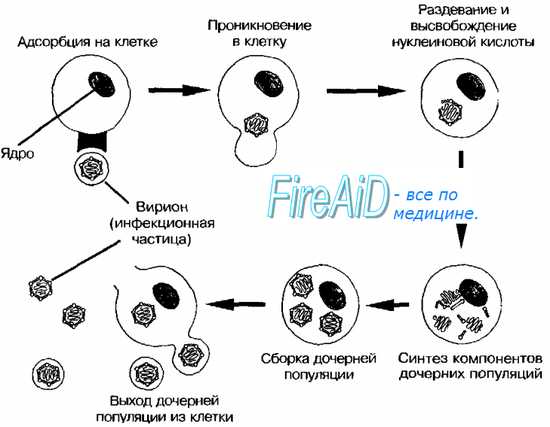
Рис. 2-3. Основные этапы репродукции вирусов.
Адсорбция вириона к клетке
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вирусные инфекции. Особенности патогенеза вирусных инфекций. Основные этапы патогенеза вирусных инфекций.
В основе патогенеза вирусных инфекций лежит взаимодействие генома вируса с генетическим аппаратом чувствительной клетки. Исключая поражения, вызываемые вирусами, распространяющимися по нервной ткани, патогенез вирусных инфекций сопровождает вирусемия (виремия)— циркуляция возбудителя в крови. В кровоток возбудитель проникает прямым путем или из лимфатической системы. Многие вирусы (например, ВИЧ, вирусы гриппа, кори, герпеса) поражают иммунокомпетентные клетки, что чаще проявляется в нарушении функций и уменьшении числа Т-хелперов, увеличении содержания и активации Т-супрессоров или В-клеток. Некоторые возбудители образуют внутриядерные или цитоплазматические тельца включений (например, тельца Бабеша~Нёгри). имеющие диагностическое значение.
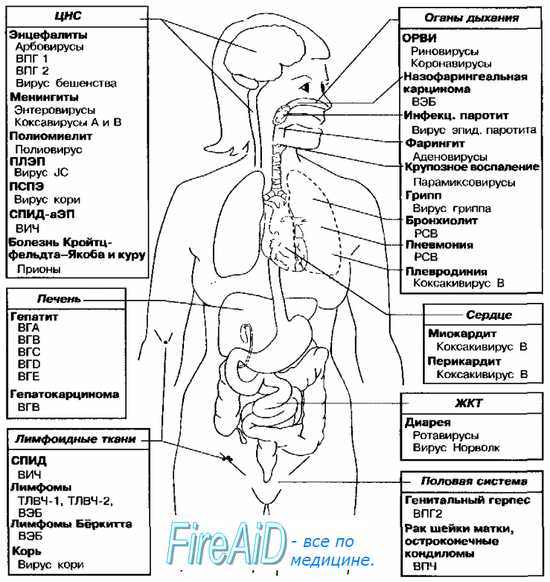
Рис. 5-9. Органы-мишени наиболее распространённых вирусных инфекций человека. ВЭБ — вирус Эпстайна-Барр, ВГА — вирус гепатита А, ВГВ — вирус гепатита В, ВГС — вирус гепатита С, BID — вирус гепатита D, ВГЕ — вирус гепатита Е, ВПЧ — вирус папилломы человека, ВПГ — вирус простого герпеса, ТЛВЧ — Т-пимфотропный вирус человека, ВИЧ — вирус иммунодефицита человека, ПЛЭП — прогрессирующая лейкоэнцефалопатия, РСВ — респираторно-синцитиальный вирус, ПСПЭ — подострый склерозирующий панэнцефалит, ОРВИ — острая респираторная вирусная инфекция, СПИД-аЭП — СПИД-ассоциированная энцефалопатия.
Основные этапы патогенеза вирусных инфекций
Проникновение вируса в организм. Основные входные ворота для возбудителей вирусных инфекции человека — дыхательные пути и ЖКТ, реже — кожные покровы. В некоторых случаях развиваются локальные поражения, но чаще в месте проникновения не возникает каких-либо проявлений или они носят стёртый характер, а возбудитель мигрирует в чувствительные ткани. Распространение возбудителя в организме может носить локальный или системный характер.
Локальные поражения вирусами типичны для возбудителей респираторных и кишечных инфекций, а также для некоторых кожных заболеваний. Продолжительность инкубационного периода большинства подобных инфекций составляет 2-3 сут. Первичную репликацию часто сопровождает вирусемия. Она обычно протекает бессимптомно или по типу продромальных явлений, но может возникать и на фоне выраженной клинической картины, не вызывая развития дополнительной симптоматики. Для подобных заболеваний характерно повторное заражение, так как циркулирующие AT не проявляют протективный эффект, а секреторный иммуноглобулин А (IgA) оказывает лишь кратковременное нейтрализующее действие на слизистой оболочке. Системные поражения. Из места проникновения возбудители попадают в кровоток, вызывая вирусемию, и постепенно фиксируются в чувствительных тканях. Первичное распространение обычно вызывает продромальные явления. Поскольку вирусемия предшествует поражению чувствительных тканей, то продолжительность инкубационного и продромального периодов подобных инфекций могут увеличиваться до 2-3 нед. Вирусемия при системных инфекциях обычно носит двухэтапный характер. Первый этап заканчивается поглощением циркулирующих вирусов клетками ретикулоэндотелиальной системы.
В дальнейшем возможно несколько вариантов:
• полная элиминация возбудителя (абортивная инфекция);
• размножение вирусов в фагоцитах с последующим выходом и развитием выраженной вторичной вирусемии, сопровождающейся появлением характерных клинических признаков заболевания (например, энцефалитов);
• некоторые вирусы (например, вирус гепатита В, пикорна- и тогавирусы) слабо поглощаются фагоцитами и могут циркулировать в крови в свободном состоянии, а возбудители колорадской клещевой лихорадки и лихорадки долины Рифт внедряются в эритроциты.
Основные органы-мишени наиболее распространённых вирусных инфекций представлены на рис. 5-9. Многие из указанных на рисунке возбудителей могут поражать, кроме названных, и другие ткани (так, полиовирусы способны вызывать поражения ЖКТ, а вирус эпидемического паротита обладает тропностью к эпителию извитых канальцев яичек).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Взаимодействие вируса с клеткой хозяина - это сложный «ногоступенчатый процесс, который начинается с адсорбции вирусных частиц на рецепторах клетки хозяина и продолжается после их проникновения внутрь клетки. В результате такого взаимодействия мовивается либо продуктивная, либо абортивная, либо интегративная форма клеточной инфекции. При продуктивной форме происходит размножение, точнее, репродукция (лат. reproduce - воспроизводить) вируса, при абортивной - ее нарушение на одном из этапов, при интегративной - интеграция вирусной нуклеиновой кислоты в клеточный геном.
Продуктивная инфекция. Репродукция вирусов
Как отмечалось выше, вирусы являются самореплицирующейся формой, неспособной к бинарному делению, в отличие от микроорганизмов с клеточной организацией. В 50-х годах было установлено, что размножение, или репродукция, вирусов происходит путем репликации их нуклеиновой кислоты и биосинтеза белков с последующей самосборкой вириона. Этот процесс происходит в разных частях клетки - ядре или цитоплазме, вследствие чего получил название дизъюнктивного, т.е. разобщенного размножения.
Вирусная репродукция представляет собой уникальную форму выражения чужеродной (вирусной) информации в клетках человека и животных, насекомых, растений и бактерий, которая состоит в подчинении клеточных матрично-генетических механизмов вирусной информации.
1-я стадия - адсорбция - характеризуется прикреплением вириона к клеточным рецепторам, представляющим собой гликопротеины клеточной мембраны, содержащей нейраминовую кислоту. Такие рецепторы имеются у ряда клеток, в частности эритроцитов, на которых адсорбируются многие вирусы. Для орто- и парамиксовирусов специфическими рецепторами являются гликолипиды, содержащие сиаловую кислоту (ганглиозиды), для других - белки или липиды клеточной мембраны. Рецепторами вирусов являются так называемые прикрепительные белки, располагающиеся в составе капсидов простых вирионов и суперкапсидов сложных вирионов. Они могут иметь форму нитей (фибры у аденовирусов) или шипов (гликопротеиновые образования на внешней оболочке орто- и парамиксо-, рабдо-, арено- и буньявирусов). Первый этап адсорбции определяется неспецифическими силами межмолекулярного притяжения, второй - специфической структурной гомологией или комплементарностью рецепторов чувствительных клеток и вирусов.
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
2-я стадия - проникновение вируса в клетку хозяина - происходит несколькими путями.
Рецепторно-опосредованный эндоцитоз характеризуется образованием в месте взаимодействия вириона с клеточным рецептором окаймленных пузырьков, в формировании которых принимают участие белки-клатрины.
Виропексис. Этим путем в клетку проникают сложноустроен-ные вирусы. Он заключается в слиянии мембран - вирусного суперкапсида с клеточной или ядерной мембраной. Данный процесс происходит при помощи специального бека слияния - F-белка, который находится в суперкапсиде. В результате виропексиса капсид оказывается в клетке хозяина, а суперкапсид вместе с белком встраивается в ее плазматическую мембрану (вследствие чего клетка приобретает способность сливаться с другими клетками, что приводит к передаче вируса этим клеткам).
Фагоцитоз. Данным путем вирус проникает в фагоцитирующие клетки, что приводит к незавершенному фагоцитозу.
3-я стадия - транспорт вируса внутри клетки. Он происходит с помощью внутриклеточных мембранных пузырьков, в которых вирус переносится на рибосомы, эндоплазматическую сеть или в ядро.
5-я стадия называется эклипс-фазой, которая характеризуется исчезновением вириона, поскольку он перестает обнаруживаться при элданой микроскопии. В эту стадию начинается синтез компонентов вириона, т.е. его репродукция. Она носит дизъюнктивный (раздельный) характер, поскольку компоненты вириона синтезируются в разных частях клетки: белки на рибосомах, нуклеиновые кислоты в ядре или цитоплазме. Вирус использует для этого генетический аппарат клетки, подавляя необходимые ей самой синтетические реакции.
Эта стадия начинается с транскрипции и репликации вирусного генома. Транскрипция вирусного генома двунитевых ДНК-содержащих вирусов происходит, так же как и клеточного генома, по триаде ДНК- иРНК- белок. Различия касаются только происхождения фермента ДНК-зависимой РНК-полимеразы, необходимой для данного процесса. У вирусов, геном которых транскрибируется в цитоплазме клетки хозяина (например, вирус оспы), имеется собственная вирусспецифическая РНК-полимераза. Вирусы, геномы которых транскрибируются в ядре (папова- и аденовирусы, вирусы герпеса), используют содержащуюся там клеточную РНК-полимеразу II или III.
Теневая фаза ( фаза эклипса )
После депротеинизации вирусы невозможно выделить из культуры клеток. Этот этап репродукции известен как теневая фаза, или фаза эклипса [от англ. eclipse, затмение].
Она включает репликацию нуклеиновых кислот вируса и синтез вирусных белков. Теневая фаза не происходит при температуре 0-4 °С (исключая вирус гриппа). Различия в энергетических потребностях для теневой фазы разных групп вирусов указывают на возможное участие в этом процессе различных клеточных реакций. Теневая фаза заканчивается после образования составных компонентов вируса, необходимых для сборки дочерних популяций.
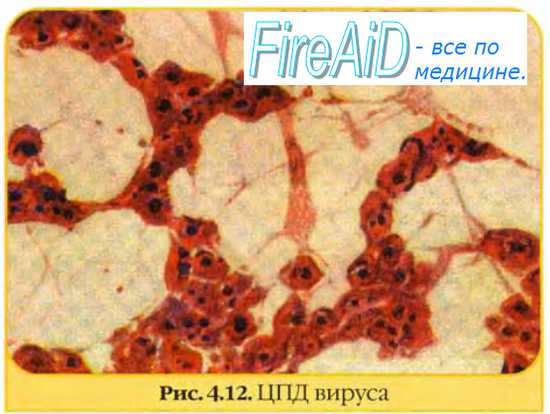
ЦПД — видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения от стекла), возникающие в результате внутриклеточной репродукции вирусов.
Образование вирусных частиц
Образование дочерних вирусных частиц в заражённой клетке подразумевает необходимость трёх процессов:
экспрессия генетического материала в виде его транскрипции и последующей трансляции, что приводит к появлению вирусных белков;
синтез генетического материала вируса (репликация);
сборка из генетического материала и вирусных белков дочерних популяций.
Следует помнить, что генетическим материалом вирионов может быть либо ДНК, либо РНК
У РНК-содержащих вирусов транскрипция их генома осуществляется несколькими путями:
1. Вирусы с негативным геномом (минус-нитевые), к которым относятся орто-, парамиксо- и рабдовирусы, имеют в своем составе вирусспецифическую РНК-полимеразу или транскриптазу. Они синтезируют иРНК на матрице геномной РНК. Подобный фермент отсутствует в нормальных клетках, но синтезируется клетками, зараженными вирусами. Он находится в составе как однонитевых, так и двунитевых РНК-содержащих вирусов.
2. У вирусов с положительным геномом (плюс-нитевые), к которым относятся пикорна-, тогавирусы и др., функцию иРНК выполняет сам геном, который транслирует содержащуюся в нем информацию на рибосомы клетки хозяина.
Транскрипция -РНК-содержащих вирусов и вирусов, имеющие две нити РНК. Функции мРНК выполняют транскрипты, комплементарные -РНК вириона. Поэтому у таких вирусов транскрипция существует как самостоятельный этап репродуктивного цикла. Для образования транскриптов в составе вирионов имеется собственная РНК-полимераза (транскриптаза).
Транскрипция +РНК-содержащих вирусов. Функции мРНК выполняет геном (+РНК), поэтому у таких вирусов для синтеза вирусных белков (трансляция) нет необходимости в процессе транскрипции. Другими словами, у +РНК-содержащих вирусов транскрипция отсутствует.
Транскрипция ДНК-вирусов. Транскрипция — самостоятельный этап репродуктивного цикла, так как геном ДНК-вирусов должен транскрибироваться для образования мРНК. Вирусы, репродуцирующиеся в ядре (например, герпес- и аденовирусы) для этой цели используют клеточную ДНК-зависимую РНК-полимеразу (транскриптазу). Вирусы, репродуцирующиеся в цитоплазме (например, поксвирусы) лишены такой способности и содержат (как и вирусы с -РНК) собственную транскриптазу.
Читайте также: