
Как заставить вирус размножатся
Обновлено: 24.04.2024
Перед развитием в культуре клеток многие вирусы культивируются в куриных яйцах с развивающимся эмбрионом. На сегодняшний день этот метод в большинстве случаев используется для выращивания вируса гриппа. Превосходный урожай вируса из куриных яиц привел к их широко распространенному использованию в исследовательских лабораториях и при производстве вакцин. В действительности подавляющее большинство противогриппозных вакцин, как инактивированных, так и инфекционных, производится в куриных яйцах. Как вирус гриппа размножается в яйцах?
Рис. 41 демонстрирует срез куриного яйца с развивающимся эмбрионом. Показаны различные пути инокуляции в яйцо, так же как и различные отделения, где размножаются вирусы.
Рисунок 41.
Для размножения вируса гриппа используются свободные от патогена яйца 11 – 12-го дня после оплодотворения. Яйцо располагают перед источником света для определения области аллантоисной полости без прожилок, расположенной под воздушным мешком. Она помечается карандашом. После того, как все яйца просвечены таким образом, делается небольшая зарубка в скорлупе в этой позиции с использованием ювелирного резца. Затем просверливается отверстие сверху яйца при помощи моторизированного инструмента Дремеля (Dremel). Если этого не сделать, то после введения вируса давление в воздушном мешке просто вытеснит посевной материал.
После того как на всех яйцах поставлена зарубка и они просверлены, в них вводится вирус при помощи шприца для инъекций туберкулина – шприц на 1 мл, заполненный на ½ дюйма, стрелка указателя в позиции 27. Игла проходит через отверстие в скорлупе, сквозь хориоаллантоисную мембрану, и вирус помещается в аллантоисной полости, заполненной аллантоисной жидкостью. Два отверстия в скорлупе запечатываются расплавленным парафином, и яйца помещают на 48 часов в термостат, установленный на 37°С.
В течение инкубационного периода вирус размножается в клетках, составляющих хориоаллантоисную мембрану. Так как новые вирусные частицы образуются почкованием (см. 4.1.), они выходят в аллантоисную жидкость. Чтобы собрать вирус, удаляют верхушку яичной скорлупы – часть, покрывающую воздушный мешок. Для этого когда-то мы пользовались специальным инструментом, который располагался над яйцом. Было легко удалить эту часть скорлупы пинцетом. Скорлуповая оболочка и хориоаллантоисная мембрана протыкались пипеткой, которая использовалась для удаления аллантоисной жидкости – примерно 10 мл в каждой яйце. Достаточное количество вируса для производства 15 мкг дозы вакцины могло быть собрано из одного или двух яиц (в зависимости от штамма вируса).
Когда-то мы выращивали так много вируса гриппа, что большая термальная комната с отдельным входом использовалась как инкубатор для яиц. Когда вы открывали дверь инкубатора и слышали писк, это значило, что кто-то оставил неиспользованные яйца на слишком долгое время и они вывелись. В этом случае вам предстояло поймать неуловимых цыплят.
Взрослые обладают перекрестно-реагирующими антителами к A/California/04/2009 (H1N1)
[Главы нет в содержании, но есть ссылка на нее из глав 7.2. и 7.3.]
Дает ли предыдущее знакомство с вирусами гриппа H1N1, обусловленное заражением или вакцинацией, какую-либо защиту против заражения новыми штаммами H1N1? Ответ на этот вопрос может прояснить, почему свыше 60% подтвержденных случаев гриппа были вызваны вирусами, подобными вирусам свиного гриппа H1N1, в США у людей от 5 до 24 лет, как сообщалось на пресс-конференции CDC.
Для ответа на этот вопрос CDC проанализировал образцы сыворотки, собранные в течение предыдущих исследований вакцины. Эти сыворотки собрали от детей и взрослых до и поле того, как их привили вакциной против гриппа, в сезоны гриппа 2005 – 2006, 2006 – 2007, 2007 – 2008 и 2008 – 2009 гг. Применялись методы нейтрализации вируса и торможения гемагглютинации для определения того, содержат ли эти сыворотки антитела, перекрестно-реагирующие с новым штаммом H1N1. В качестве образца новых изолятов вируса H1N1 использовался A/California/04/2009.
Результаты показали, что предварительная вакцинация детей (в возрасте от 6 месяцев до 9 лет, всего 79 образцов) или сезонной тривалентной инактивированной вакциной, или инфекционной аттенуированной противогриппозной вакциной в предыдущие 4 года не вызвала перекрестно-реагирующих антител для ответа на A/California/04/2009 у взрослых. Среди людей возраста от 18 до 64 лет был двукратный рост перекрестной реактивности антител к вирусу в сравнении с 12 – 19-кратным ростом в титрах антител против сезонных штаммов. У людей старше 60 лет не было роста кросс-реактивности антител. Эти данные показывают, что иммунизация сезонной противогриппозной вакциной, содержащей прежние штаммы H1N1 (2005 – 2009 гг.), похоже, не дает защиты против инфицирования новыми штаммами H1N1.
Важным является вопрос, содержат ли сыворотки, полученные до введения вакцины, перекрестно-реагирующие титры антител против A/California/04/2009. Такой анализ показал бы, дает ли естественное заражение штаммами H1N1 некоторую защиту против новых изолятов. У 79 детей в возрасте от 6 месяцев до 9 лет до вакцинирования не было обнаружено перекрестно-реагирующих антител в сыворотке к A/California/04/2009. Однако 6% взрослых в возрасте от 18 до 40 лет, 9% – от 18 до 64 лет и 33% старше 60 лет имели перед вакцинацией нейтрализующие титры антител к A/California/04/2009 выше 160 или равные этому значению. Эти антитела, скорее всего, были приобретены с инфицированием вирусом H1N1, по антигенным свойствам более близким к A/California/04/2009, чем к другим штаммам сезонного H1N1. Неизвестно, могут ли эти антитела дать защиту против заражения, но они могут уменьшить тяжесть симптомов болезни.
Я подозреваю, что не все знакомы с методами микронейтрализации и торможения гемагглютинации, используемыми в этом исследовании. Впоследствии я объясню механизм действия этих методов и важность их результатов.
Рекомендуемая литература.
- J Katz, PhD, K Hancock, PhD, V Veguilla, MPH, W Zhong, PhD, XH Lu, MD, H Sun, MD, E Butler, MPH, L Dong, MD, PhD, F Liu, MD, PhD, ZN Li, MD, PhD, J DeVos, MPH, P Gargiullo, PhD, N Cox, PhD (2009). Serum Cross-Reactive Antibody Response to a Novel Influenza A (H1N1) Virus After Vaccination with Seasonal Influenza Vaccine Morbid.Mortal. Weekly Rep., 58 (19), 521-524.
7.2. Метод торможения гемагглютинации гриппа
Центры по контролю и профилактике заболеваний определили, что некоторые взрослые имеют сывороточные перекрестно-реагирующие антитела к новому вирусу гриппа H1N1. Чтобы сделать такой вывод, использовали одну из методик, называемую методом торможения гемагглютинации. Как он действует?
7.3. Анализ микронейтрализации гриппа
Анализ микронейтрализации – другой метод, с помощью которого Центры по контролю и профилактике заболеваний определили, что некоторые взрослые имеют перекрестно-реагирующие антитела к новому вирусу гриппа H1N1. Рассмотрим, как действует этот метод.
Репликация вируса часто исследуется в лаборатории путем инфицирования клеток, выращиваемых в пластмассовых планшетах или колбах, которые обычно называются клеточными культурами. На рис. 43 – пример клеток HeLa, убиваемых полиовирусом.
Рисунок 43.
Верхний левый квадрат показывает неинфицированные клетки, а другие части показывают клетки в течение определенного времени после заражения. По мере размножения вируса зараженные клетки округляются и отделяются от планшета с культурой клеток. Эти видимые изменения называются цитопатическим эффектом.
Есть и другой путь сделать видимым уничтожение клеток вирусом без использования микроскопа – окрашивание клеток красителем. На рис. 44 клетки были расположены в маленьких лунках 96-луночного планшета. В одну лунку добавили вирус, а в другие – нет. После периода инкубации клетки окрасились в кристаллический фиолетовый, который окрашивает только живые клетки. Видно, какие клетки были поражены вирусом, а какие нет.
7.4. Определение вирусов: анализ бляшкообразования
Одним из наиболее важных процессов в вирусологии при измерении титра вируса является концентрация вирусов в образце. Широко используемый подход для определения количества инфекционного вируса – это анализ бляшкообразования. Данная методика впервые была разработана для подсчета титров линии бактериофагов. Р. Дульбекко (Renato Dulbecco) модифицировал этот процесс в 1952 г. для использования в вирусологии животных, и с тех пор он используется для надежного определения титров многих различных вирусов.
Рисунок 46.
Для проведения анализа бляшкообразования приготавливают 10-кратные разведения линии вирусов и вводят 0,1 мл аликвоты в чувствительные клеточные монослои. После инкубационного периода, во время которого вирус прикрепляется к клеткам, монослои покрываются питательной средой, содержащей вещество (обычно агар), способствующее образованию геля. Когда планшеты инкубируют, в изначально зараженных клетках выводится вирусное потомство. Распространение новых вирусов ограничено гелем до соседних клеток. Следовательно, каждая инфекционная частица производит круговую зону зараженных клеток, называемую бляшкой. В итоге бляшка становится достаточно большой, чтобы стать видимой невооруженным глазом. Краситель, помечающий живые клетки, часто используется для усиления контраста между живыми клетками и бляшками. Этим способом пригоден для анализа только тех вирусов, которые причиняют видимый ущерб клеткам. Пример бляшек, образованных полиовирусом на монослое клеток HeLa, показан на рис. 46. Клетки окрашены кристаллическим фиолетовым, а бляшки без труда видны там, где клетки разрушены вирусной инфекцией.
Титр линии вируса может быть подсчитан в бляшкообразующих единицах (PFU) на миллиметр. Для определения титра вируса подсчитываются бляшки. В целях снижения погрешности подсчитываются только планшеты, содержащие от 10 до 100 бляшек, в зависимости от размера используемого планшета с культурой клеток. Согласно правилам статистики, при подсчете 100 бляшек титр образца будет варьироваться в пределах ± 10%. Каждое разведение проводится дважды для повышения точности. На рис. 47 показано 17 бляшек на планшете, сделанном из разведения 10 -6 . Поэтому титр вируса составит 1,7 × 10 8 PFU/мл.
Рисунок 47.
Впоследствии мы обсудим, как анализ бляшкообразования может использоваться для приготовления клональных линий вирусов – очень важной стадии в исследовании вирусной генетики.
Рекомендуемая литература.
- Dulbecco, R., & Vogt, M. (1953). Some problems of animal virology as studied by the plaque technique. Cold Spring Harbor Symp. Quant. Biol., 18, 273-279.
7.5. Сколько вирусов требуется для образования бляшки?
Анализ бляшкообразования – главный инструмент для определения титров вируса. Идея проста: вирусная инфекция ограничивается до соседних клеток полутвердым верхним слоем. Подсчитывая количество бляшек, можно вычислить титр вируса в PFU на мл. Основной вопрос: сколько нужно вирусов для формирования одной бляшки?
Для большинства вирусов животных одной инфекционной частицы бывает достаточно для начала инфекции. К этому выводу можно прийти, изучая отношения между количеством вирусных частиц и подсчетом бляшек. Линейная зависимость означает, что бляшку может образовать одна инфекционная частица. Считается, что в этом случае вирус заразил клетку с динамикой в один удар. Эта концепция продемонстрирована на рис. 48. Количество бляшек, образованных с динамикой одного или двух ударов показано на графике в сравнении с относительной концентрацией вируса.
Рисунок 48.
Есть некоторые примеры вирусов с динамикой в два удара, другими словами, два различные типа вирусных частиц должны заразить клетку для запуска инфекционного цикла. Примеры включают геномы некоторых РНК вирусов растений с (+) нитью, состоящие из двух молекул РНК, упакованных в различные частицы. Кривая зависимости от дозы подобных вирусов, скорее, параболическая, чем линейная.
Когда единичная вирусная частица способна образовать бляшку, вирусное потомство внутри бляшки представляет собой клоны. Вирусная линия, приготовленная из единичной бляшки, называется очищенной линией вирусов из бляшки. Чтобы приготовить такую линию вирусов, наконечник маленькой пипетки вставляют в верхний слой агара над бляшкой. Пробка агара удаляется и помещается в буферный раствор. Вирусы в пробке агара перемещаются в буферный раствор, который затем можно использовать для заражения клеточных культур. Для обеспечения чистоты этот процесс обычно повторяют как минимум еще раз. В значительной степени очищение бляшки используется в вирусологии для основания клональных линий вирусов. Способность подготовки клональных линий вирусов было важным шагом, позволившим проводить генетический анализ вирусов.
7.6. Измерение вирусов методом конечного разведения
Анализ бляшкообразования – замечательный метод определения титров вируса, но он не применим для всех вирусов. К счастью, доступно несколько альтернативных методов, включая метод конечного разведения.
Метод конечного разведения использовался для измерения титра вируса до развития метода бляшкообразования и все еще применяется для вирусов, не образующих бляшек. Подготавливаются последовательные разведения линии вируса, которые вносятся в копируемые культуры клеток, часто в многолуночных форматах (например, в 96-луночный пластмассовый планшет). Количество заражаемых культур клеток определяется впоследствии для каждого разведения вируса, обычно путем поиска цитопатического эффекта.
В данном примере метода конечного разведения каждый из 10 монослоев клеточных культур был инфицирован разведением вируса. После инкубационного периода планшеты, показавшие цитопатические эффекты, были помечены +. При высоких разведениях ни одна из клеточных культур не была заражена, так как не было частиц. При низких разведениях каждая культура клеток была инфицирована. Половина клеточных культур показала цитопатические эффекты при разведении 10 -5 . Это и есть конечная точка: разведение вируса, при котором 50% клеточных культур оказалось заражено. Это количество может быть высчитано на основании данных и выражено как 50% инфекционной дозы (ИД50) на миллилитр. Линия вирусов в приведенном примере содержит 10 5 ИД50 на мл.
Рисунок 49.
В реальной жизни конечная точка со значением 50% обычно не встречается, как мы показали на примере разведения. Поэтому привлекаются статистические методы подсчета конечной точки титрования.
Методы конечного разведения также могут использоваться при определении вирулентности вируса у животных. Применяется тот же подход: производятся последовательные разведения вирусов и вводятся в ряд подопытных животных. Заражение животного можно определить по его смерти или клиническим симптомам, таким как повышенная температура, потеря веса или паралич. Результаты выражаются в виде 50% летальной дозы (LD50) на мл или 50% паралитической дозы (PD50) на мл, когда смертность или паралич используются как конечные точки.
Следующий пример поясняет использование конечного разведения при подсчете смертности от полиовируса у мышей. Восьми мышам ввели разведение вируса, а конечной точкой была смерть. Статистический метод Рида – Мюнха использовался для определения конечной точки со значением 50%. В этом методе объединены результаты, и смертность подсчитана для каждого разведения. Конечная точка в 50%, которая находится между пятым и шестым разведениями, подсчитана как 10 -6,5 . Поэтому образец вируса содержит 10 6,5 LD50 единиц.

Обзор
Автор
Редакторы
Обратите внимание!
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science.
Эволюция и происхождение вирусов
В 2007 году сотрудники биологического факультета МГУ Л. Нефедова и А. Ким описали, как мог появиться один из видов вирусов — ретровирусы. Они провели сравнительный анализ геномов дрозофилы D. melanogaster и ее эндосимбионта (микроорганизма, живущего внутри дрозофилы) — бактерии Wolbachia pipientis. Полученные данные показали, что эндогенные ретровирусы группы gypsy могли произойти от мобильных элементов генома — ретротранспозонов. Причиной этому стало появление у ретротранспозонов одного нового гена — env, — который и превратил их в вирусы. Этот ген позволяет вирусам передаваться горизонтально, от клетки к клетке и от носителя к носителю, чего ретротранспозоны делать не могли. Именно так, как показал анализ, ретровирус gypsy передался из генома дрозофилы ее симбионту — вольбахии [7]. Это открытие упомянуто здесь не случайно. Оно нам понадобится для того, чтобы понять, чем вызваны трудности борьбы с вирусами.
Из давних письменных источников, оставленных историком Фукидидом и знахарем Галеном, нам известно о первых вирусных эпидемиях, возникших в Древней Греции в 430 году до н.э. и в Риме в 166 году. Часть вирусологов предполагает, что в Риме могла произойти первая зафиксированная в источниках эпидемия оспы. Тогда от неизвестного смертоносного вируса по всей Римской империи погибло несколько миллионов человек [8]. И с того времени европейский континент уже регулярно подвергался опустошающим нашествиям всевозможных эпидемий — в первую очередь, чумы, холеры и натуральной оспы. Эпидемии внезапно приходили одна за другой вместе с перемещавшимися на дальние расстояния людьми и опустошали целые города. И так же внезапно прекращались, ничем не проявляя себя сотни лет.
Вирус натуральной оспы стал первым инфекционным носителем, который представлял действительную угрозу для человечества и от которого погибало большое количество людей. Свирепствовавшая в средние века оспа буквально выкашивала целые города, оставляя после себя огромные кладбища погибших. В 2007 году в журнале Национальной академии наук США (PNAS) вышла работа группы американских ученых — И. Дэймона и его коллег, — которым на основе геномного анализа удалось установить предположительное время возникновения вируса натуральной оспы: более 16 тысяч лет назад. Интересно, что в этой же статье ученые недоумевают по поводу своего открытия: как так случилось, что, несмотря на древний возраст вируса, эпидемии оспы не упоминаются в Библии, а также в книгах древних римлян и греков [9]?
Строение вирусов и иммунный ответ организма

Рисунок 1. Первооткрыватель вирусов Д.И. Ивановский (1864–1920) (слева) и английский врач Эдвард Дженнер (справа).

Почти все известные науке вирусы имеют свою специфическую мишень в живом организме — определенный рецептор на поверхности клетки, к которому и прикрепляется вирус. Этот вирусный механизм и предопределяет, какие именно клетки пострадают от инфекции. К примеру, вирус полиомиелита может прикрепляться лишь к нейронам и потому поражает именно их, в то время как вирусы гепатита поражают только клетки печени. Некоторые вирусы — например, вирус гриппа А-типа и риновирус — прикрепляются к рецепторам гликофорин А и ICAM-1, которые характерны для нескольких видов клеток. Вирус иммунодефицита избирает в качестве мишеней целый ряд клеток: в первую очередь, клетки иммунной системы (Т-хелперы, макрофаги), а также эозинофилы, тимоциты, дендритные клетки, астроциты и другие, несущие на своей мембране специфический рецептор СD-4 и CXCR4-корецептор [13–15].

Одновременно с этим в организме реализуется еще один, молекулярный, защитный механизм: пораженные вирусом клетки начинают производить специальные белки — интерфероны, — о которых многие слышали в связи с гриппозной инфекцией. Существует три основных вида интерферонов. Синтез интерферона-альфа (ИФ-α) стимулируют лейкоциты. Он участвует в борьбе с вирусами и обладает противоопухолевым действием. Интерферон-бета (ИФ-β) производят клетки соединительной ткани, фибробласты. Он обладает таким же действием, как и ИФ-α, только с уклоном в противоопухолевый эффект. Интерферон-гамма (ИФ-γ) синтезируют Т-клетки (Т-хелперы и (СD8+) Т-лимфоциты), что придает ему свойства иммуномодулятора, усиливающего или ослабляющего иммунитет. Как именно интерфероны борются с вирусами? Они могут, в частности, блокировать работу чужеродных нуклеиновых кислот, не давая вирусу возможности реплицироваться (размножаться).

Причины поражений в борьбе с ВИЧ
Тем не менее нельзя сказать, что ничего не делается в борьбе с ВИЧ и нет никаких подвижек в этом вопросе. Сегодня уже определены перспективные направления в исследованиях, главные из которых: использование антисмысловых молекул (антисмысловых РНК), РНК-интерференция, аптамерная и химерная технологии [12]. Но пока эти антивирусные методы — дело научных институтов, а не широкой клинической практики*. И потому более миллиона человек, по официальным данным ВОЗ, погибают ежегодно от причин, связанных с ВИЧ и СПИДом.

Подобный вирусный механизм характерен не только для ВИЧ. Он описан и при инфицировании некоторыми другими опасными вирусами: такими, как вирусы Денге и Эбола. Но при ВИЧ антителозависимое усиление инфекции сопровождается еще несколькими факторами, делая его опасным и почти неуязвимым. Так, в 1991 году американские клеточные биологи из Мэриленда (Дж. Гудсмит с коллегами), изучая иммунный ответ на ВИЧ-вакцину, обнаружили так называемый феномен антигенного импринтинга [23]. Он был описан еще в далеком 1953 году при изучении вируса гриппа. Оказалось, что иммунная система запоминает самый первый вариант вируса ВИЧ и вырабатывает к нему специфические антитела. Когда вирус видоизменяется в результате точечных мутаций, а это происходит часто и быстро, иммунная система почему-то не реагирует на эти изменения, продолжая производить антитела к самому первому варианту вируса. Именно этот феномен, как считает ряд ученых, стоит препятствием перед созданием эффективной вакцины против ВИЧ.

Открытие биологов из МГУ — Нефёдовой и Кима, — о котором упоминалось в самом начале, также говорит в пользу этой, эволюционной, версии.

Сегодня не только ВИЧ представляет опасность для человечества, хотя он, конечно, самый главный наш вирусный враг. Так сложилось, что СМИ уделяют внимание, в основном, молниеносным инфекциям, вроде атипичной пневмонии или МЕRS, которыми быстро заражается сравнительно большое количество людей (и немало гибнет). Из-за этого в тени остаются медленно текущие инфекции, которые сегодня гораздо опаснее и коварнее коронавирусов* и даже вируса Эбола. К примеру, мало кто знает о мировой эпидемии гепатита С, вирус которого был открыт в 1989 году**. А ведь по всему миру сейчас насчитывается 150 млн человек — носителей вируса гепатита С! И, по данным ВОЗ, каждый год от этой инфекции умирает 350-500 тысяч человек [33]. Для сравнения — от лихорадки Эбола в 2014-2015 гг. (на состояние по июнь 2015 г.) погибли 11 184 человека [34].
* — Коронавирусы — РНК-содержащие вирусы, поверхность которых покрыта булавовидными отростками, придающими им форму короны. Коронавирусы поражают альвеолярный эпителий (выстилку легочных альвеол), повышая проницаемость клеток, что приводит к нарушению водно-электролитного баланса и развитию пневмонии.

Рисунок 8. Электронная микрофотография воссозданного вируса H1N1, вызвавшего эпидемию в 1918 г. Рисунок с сайта phil.cdc.gov.
Почему же вдруг сложилась такая ситуация, что буквально каждый год появляются новые, всё более опасные формы вирусов? По мнению ученых, главные причины — это сомкнутость популяции, когда происходит тесный контакт людей при их большом количестве, и снижение иммунитета вследствие загрязнения среды обитания и стрессов. Научный и технический прогресс создал такие возможности и средства передвижения, что носитель опасной инфекции уже через несколько суток может добраться с одного континента на другой, преодолев тысячи километров.
Мы кратко рассмотрели структуру вирионов гриппа и процесс кодирования вирусными РНК одного и более белков. Теперь обратимся к размножению вирусов гриппа.
Вирусы – это исключительно внутриклеточные паразиты, они не могут воспроизводиться вне клетки. Появление новых инфекционных частиц должно протекать внутри клетки. Войдя в клетки, вирусы начинают паразитировать в структуре хозяина для воспроизводства вирусного потомства. Все события, происходящие в инфицированной вирусом клетке, называются инфекционным циклом, или вирусной репликацией. Вирусологи искусственно разделяют инфекционный цикл на несколько стадий, чтобы проще было его изучить. Эти стадии включают прикрепление и вхождение вириона, трансляцию мРНК в белок, репликацию генома (создание большего количества РНК или ДНК), скопление новых частиц и выход частиц из клетки. Мы рассмотрим каждую из названных стадий, а затем обсудим, как вирус гриппа инфицирует нас и вызывает болезнь.
Остановимся на первой стадии – прикреплении вириона к клетке. Возьмем обычную клетку, с которой, я уверен, знаком каждый. Но не будет лишним повториться.
Рисунок 10.
Слева показан клеточный белок, прикрепленный к цитоплазматической мембране. Снизу изображена внутренняя часть клетки – цитоплазма. Часть белка пересекает мембрану, и также видны части цитоплазматической и внеклеточной сторон. Сферы – это сахара, прикрепленные ко множеству белков (белок + сахар = гликопротеин). Сиаловая кислота всегда является последним сахаром в цепи, которая прикрепляется к белку. Справа показана химическая структура сиаловой кислоты; следующий сахар (справа) – галактоза. Вирионы гриппа прикрепляются к клеткам, когда HA захватывает очень маленькую сиаловую кислоту.
Сахар в сравнении с HA в действительности очень мал – он подходит к небольшому кармашку на острие шипа. На рис. 11 изображена молекулярная модель, демонстрирующая, как HA прикрепляется к аналогу сиаловой кислоты. Шаровидный конец HA показан в верхней части рис. 11. Маленькие красные и белые сферы показывают места будущего прикрепления сиаловой кислоты – кармашки на верхнем конце HA.
Рисунок 13.
Процесс вхождения вируса гриппа в клетки – наиболее понятный из всех известных механизмов проникновения вирусов. После прикрепления вириона к сиаловой кислоте, имеющей рецепторы на поверхности клетки, вирус-рецепторный комплекс входит в клетки посредством эндоцитоза – процесса, в ходе которого клетки обычно берут молекулы из внеклеточной жидкости. Пока эндосомальные везикулы, содержащие частицы вируса, движутся к ядру клетки, их pH понижается. Это изменение в pH завершает клеточный канал, закачивающий протоны (H+) в везикулу. Когда эндосомальная pH достигает 5,0, вирусный белок HA подвергается конформационной перестройке. Это изменение подвергает пептид слияния на HA короткой гидрофобной последовательности, вставляющей его в эндосомальную мембрану, которая от этого сливается с вирусной оболочкой. Когда это происходит, вирусные РНК проникают в цитоплазму. Затем они транспортируются в ядро клетки, где начинается размножение.
В вирионе гриппа вирусные РНК не оголены, поскольку к ним прикреплены вирусные белки, включая белок M1. Этот белок образует оболочку, лежащую в основе липидной мембраны вириона. К сожалению, если вирусные РНК прикрепляются к белку M1, когда выходят из вириона, они не могут достичь ядра. Для решения этой проблемы вирион гриппа имеет в своей мембране несколько копий белка, называемого M2. Этот вирусный белок образует канал в мембране, который активно закачивает протоны из эндосомы во внутреннюю часть вириона. Данные протоны понижают pH внутри вириона, освобождая вирусные РНК от M1. В этом случае РНК могут войти в ядро.
Ионный канал M2, являющийся мишенью для противовирусных адамантанов, показан на рис. 14. Эти образования забивают канал и не позволяют протонам закачиваться в вирион. В присутствии адамантанов вирусным РНК нужно прикрепляться к M1, отчего они не могут достичь ядра. Поэтому вирусная репликация подавляется. Устойчивость к адамантанам возможна при изменении в аминокислотах, прокладывающих канал M2. Такие изменения не дают лекарству забить канал.
Рисунок 15.
На рис. 15 сферический конец белка HA, который прикрепляется к рецепторам клетки, изображен сверху, а вирусная мембрана – снизу. Для наглядности помечен только сайт расщепления HA. Нерасщепленная форма белка называется HA0; после расщепления клеточным ферментом образуются два белка, называемые HA1 (синий) и HA2 (красный). Две субъединицы остаются вместе на поверхности вирусной частицы. Новая амино(N)-концевая область HA2, образовавшегося при расщеплении, содержит последовательность гидрофобных аминокислот, называемых пептидом слияния. Во время вхождения вируса гриппа в клетки пептид слияния вставляется в эндосомальную мембрану и вызывает слияние вирусной и клеточной оболочек. Следовательно, вирусные РНК гриппа могут войти в цитоплазму. Процесс слияния описан в предыдущем параграфе.
Если белок HA не расщеплен, чтобы образовать HA1 и HA2, слияния не происходит. Поэтому вирусы гриппа с нерасщепленным HA не заразны. Расщепление вирусного HA происходит после того, как вновь синтезированные вирионы выходят из клетки. Вирусы гриппа эффективно размножаются в яйцах из-за присутствия протеазы в аллантоисной жидкости, способной расщеплять HA. Однако репликация многих штаммов вируса гриппа в культурах клеток требует добавления в среду соответствующей протеазы (как привило, трипсина).
У людей репликация вируса гриппа ограничена дыхательными путями, потому что это единственный участок, где производится протеаза, расщепляющая HA. Однако белок HA высокопатогенных штаммов H5 и H7 вируса птичьего гриппа может расщепляться протеазами, производимыми во многих различных тканях. В результате эти вирусы способны размножаться во многих органах птиц, включая селезенку, печень, легкие, почки и мозг. Это свойство объясняет способность штаммов H5N1 вируса птичьего гриппа размножаться вне дыхательных путей человека.
Подобно белкам HA высокопатогенных вирусов H5 и H7, HA штамма вируса гриппа 1918 г. может также расщепляться повсеместно присутствующими клеточными протеазами. Следовательно, вирус может размножаться в культурах клеток без добавления трипсина.
Белки HA H5 и H7 имеют множество основных аминокислотных остатков на сайте расщепления HA1-HA2, которые делают возможным расщепление посредством широко представленных протеаз. Но HA вируса гриппа H1 1918 г. не обладал этим свойством. Также N1 1918 г. не был в состоянии привлечь протеазы, которые расщепляли бы HA, т. е. не было механизма, позволяющего штамму A/WSN/33 вируса гриппа размножаться в клетках без трипсина. Понимание того, как белок HA вируса H1 1918 г. мог быть расщеплен протеазами – основной в понимании высокой патогенности данного штамма.
Рекомендуемая литература.
Chaipan, C., Kobasa, D., Bertram, S., Glowacka, I., Steffen, I., Solomon Tsegaye, T., Takeda, M., Bugge, T., Kim, S., Park, Y., Marzi, A., &Pohlmann, S. (2009). Proteolytic Activation of the 1918 Influenza Virus Hemagglutinin Journal of Virology, 83 (7), 3200-3211 DOI:10.1128/JVI.02205-08.
Одни вирусы способны интегрироваться в геном клетки-мишени и таким образом оставаться во всех дочерних клетках, которые будут в будущем получены после ее деления. К таким вирусам относятся гаммаретровирусы и лентивирусы. Другие делать этого не умеют (например, адено- и аденоассоциированные вирусы). Но для производства белков и репликации (размножения) все они используют клетку и ее синтетический аппарат.
Как это работает?
Для того чтобы вирус мог проникнуть в клетку, белки его оболочки должны связаться с мембранными белками клетки-мишени. Важно отметить, что проникает вирус только в те клетки, которые могут в дальнейшем помочь его репликации. Вирус ВИЧ живет в клетках иммунной системы, вирус гепатита С — в клетках печени. Есть особые вирусы, которые поражают только растения или даже только бактерии.
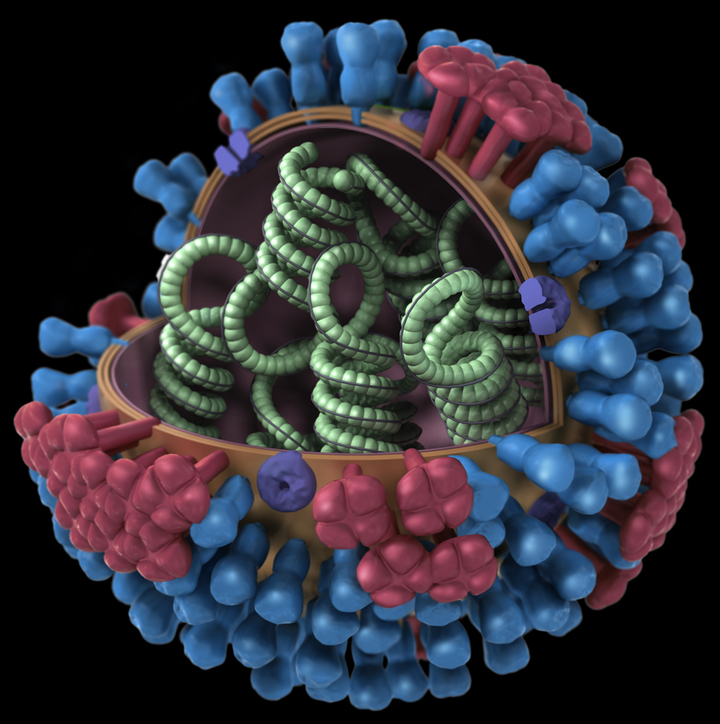
Строение вируса гриппа. Под оболочкой вириома - генетический материал вируса, необходимый для его воспроизводства в клетке.
Содержащие неактивный вирус и оставшиеся в живых клетки иногда сохраняют возможность нормального функционирования. В этом случае клетки могут быть заражены, но вирус проявит себя спустя длительный период времени. Так устроен герпес.
В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы. И тут стоит остановиться на классификации.
Типы вирусов. Коротко о главном
Современная типология вирусов содержит 7 классов и была предложена Дэвидом Балтимором еще в 1971 году. С тех пор, впрочем, она была уточнена и расширена, в том числе советскими учеными. И выглядит в настоящее время таким образом:
Вирусы, содержащие двухцепочечную ДНК
Описание
Для репликации вирусу необходимо попасть в ядро клетки-мишени и воспользоваться ее ДНК-полимеразой. Иногда вирус вызывает незапланированное деление самое клетки, то есть становится онкогенным. Эти вирусы хорошо изучены.
Пример: Вирус герпеса, адено- и папилломавирусы
Вирусы, содержащие одноцепочечную ДНК
Описание
Попадая в ядро клетки, вирусы образуют двухцепочечную ДНК, после чего реплицируются так же, как вирусы класса I.
Пример: Парво- и цирковирусы
Вирусы, в которых РНК способна к репликации (редупликации)
Описание
Вирусы этого класса могут размножаться в цитоплазме клетки, им не нужна молекула ДНК. Каждый ген, находящийся в РНК вируса, кодирует только один вирусный белок.
Пример: Бирна- и реовирусы
Вирусы, содержащие одноцепочечную (+) РНК
Описание
Из геномной (+) РНК на рибосомах хозяина создаются вирусные белки. В одном фрагменте РНК могут быть закодированы разные белки, что увеличивает сложность вируса без удлинения генов.
Пример: Пикорнавирусы (полиомиелит, гепатит А) и коронавирусы
Вирусы, содержащие одноцепочечную (–) РНК
Описание
(–) РНК этих вирусов предварительно должна быть транскрибирована в (+) РНК вирусными РНК-полимеразами, после чего может начаться синтез вирусных белков. Вирусы этого класса делятся еще на две группы, в зависимости от их генома и места его репликации (цитоплазма или ядро).
Пример: Филовирусы, аренавирусы (геморрагическая лихорадка Ласса), ортомиксовирусы (вирусы гриппа) и так далее.
Вирусы, содержащие одноцепочечную (+) РНК, реплицирующиеся через стадию ДНК
Описание
Такие вирусы используют фермент обратную транскриптазу для превращения (+) РНК в ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина.
Пример: Ретровирусы (в том числе ВИЧ)
Вирусы, содержащие двухцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК
Описание
Молекула ДНК замкнута в кольцо и является матрицей для синтеза мРНК и дополнительных молекул РНК, которые используются при репликации вирусного генома обратными транскриптазами.
Пример: Колимовирусы (вызывают инфекции растений) и гепаднавирусы (например, гепатит В)
Вакцинация и лечение
Как правило, организмы умеют бороться с паразитирующими на них вирусами. На примере млекопитающих и человека мы обычно говорим о главном инструменте — врожденном иммунитете.
Впрочем, наиболее эффективен этот вид защиты в отношении бактериальных инфекций и не может обеспечить продолжительную и надежную защиту, особенно от инфекций вирусных.
Именно поэтому огромное значение имеет приобретенный иммунитет, в результате которого клетки иммунной системы обучаются вырабатывать специфические к вирусу антитела, способные уничтожать как саму вирусную частицу, так и зараженные ею клетки.
Еще одна врожденная система борьбы с вирусными инфекциями — внутриклеточная. Как правило, клетка способна распознать чужеродную РНК в своей цитоплазме, куда ее сперва и доставляют многие вирусы, и имеет специальные комплексы для ее деградации. Но часть вирусов научились обходить и эту ловушку. К примеру, ротавирусы, которые даже внутри клетки сохраняют капсид с геномной РНК.
Миссия: уничтожить
Основная сложность в лечении вирусных заболеваний заключается в том, что они используют естественные функции клеток-мишеней для своего размножения, поэтому ученым зачастую оказывается не так-то просто придумать препарат, который будет токсичен для вируса и безопасен для самой клетки. Если такой безопасности достичь не удастся, лекарство будет иметь слишком много побочных эффектов, повреждающих сам организм, что окажется нецелесообразно для использования.

Сравнение жизненных циклов ВИЧ и вируса гриппа. Если первый использует обратную транскрипцию и живет в клетках иммунной стистемы, вирион второго, проникая в эпительные клетки дыхательных путей целиком - а именно там он и обитает - распадается уже внутри клетки, а репликация вирусной РНК происходит в ядре с помощью вирусных полимераз PA, PB1 и PB2 путем комплементарного копирования.
По принципу действия противовирусные препараты подразделяются на две группы: стимулирующие иммунную систему атаковать вирусы (например, за счет индукции синтеза белков-интерферонов) и атакующие вирусы напрямую. Препараты второй группы различаются по этапу жизненного цикла вируса, на котором они активны: это препараты, препятствующие проникновению вируса в клетку, препятствующие размножению вируса внутри клетки и препятствующие выходу копий вируса из клетки.
Еще один класс противовирусных препаратов блокирует ферменты, необходимые для создания и модификаций белков вируса. Такие лекарства называют протеазными ингибиторами.
Вместо заключения: а могут ли вирусы приносить пользу?
Безусловно, да. Несмотря на то, что вирусы ассоциируются у большинства людей с однозначным вредом, они могут приносить и пользу — если речь идет о так называемых вирусных векторах и терапевтических подходах на их основе.
Исследователи давно научились помещать в белковую оболочку вируса интересующие их нуклеиновые кислоты, чтобы доставлять нужный ген в клетки, а также убирать те гены, которые делают вирус опасным для организма.
Это позволило сделать возможной генную терапию, помогающую бороться с заболеваниями, вызванными известными генетическими мутациями. Создание вирусных векторов — достаточно непростая задача, к тому же ограниченная свойствами самих вирусных частиц: количеством помещающейся генетической информации, местом ее вставки, стабильностью. Кроме того, вирусный вектор, используемый в медицине, не должен вызывать иммунного ответа или критично влиять на жизнедеятельность клетки. Тем не менее эти сложности решаются, поэтому уже одобрен ряд вполне успешных и безопасных генных терапий. А в качестве основы для вирусных векторов чаще всего используются ретро-, ленти-, адено- и аденоассоциированные вирусы.
Читайте также: