
Классификация рнк содержащих вирусов в ветеринарии
Обновлено: 19.04.2024
Васильев Д.А. . Луговцев В. Ю Макаров В.В., Середа А.Д. Классификация и номенклатура вирусов. Ульяновск 1999.
Освещены современные данные в области классификации вирусов с учетом существующих требований и изменений по данным Международного комитета по таксономии вирусов (References: Murphy F.A., Fauquet C.M., Bishop D.H. et al.
Virus Taxonomy. Six Report ICTV. 1995).
Приведен таксономический каталог важнейших зоопатогенных вирусов. Специальное внимание уделено новым таксонам.
Учебное пособие подготовлено сотрудниками УГСХА, РУДН, ВНИИ ветеринарной вирусологии и микробиологии, на основе читаемых для студентов факультета ветеринарной медицины курса вирусологии.
С ростом числа обнаруженных вирусов были предприняты первые попытки классификации внутри этой самостоятельной группы микроорганизмов. В качестве критериев использовали сходство их патогенных свойств, органотропизм, общность эпизоотологического и экологического статуса. При этом происходило объединение весьма различающихся, как было выяснено впоследствие, групп вирусов. Так, вирусы гепатита включили возбудителей, известных теперь как вирусы гепатита А, гепатита В и желтой лихорадки. Пневмотропные (респираторные) вирусы, образующие наряду с нейротропными, пантропными и дермотропными вирусами одну из групп в органотропной классификации, включали вирусы гриппа, риновирусы, аденовирусы. Передающиеся насекомыми арбовирусы теперь известны как флави-, бунья-, реовирусы. Одна из первых классификаций состояла в разделении вирусов по принципу поражения определенных хозяев, т.е. были выделены вирусы растений, животных, насекомых и фаги. Некоторые наиболее удавшиеся групповые названия вирусов из первых классификаций укоренились и до сих пор используются в вирусологии; хотя они не признаны официальной классификацией и номенклатурой, но могут быть полезны в клинических, эпизоотологических и диагностических контекстах.
Одной из основных прагматических целей универсальной таксономической системы является однозначная идентификация вирусов. Поэтому современная классификация вирусов преследует не только академические цели, но имеет и важнейшее практическое преломление (диагностика, антигены, вакцины, иммунология, происхождение эпизоотий и эпидемий, прогностика). В последнее время этот аспект приобретает особое значение из-за возникновения множества эмерджентных инфекций, вызванных новыми вирусами или известными вирусами в необычных экологических условиях. Среди наиболее впечатляющих примеров эмерджентных инфекций - синдром приобретенного иммунодефицита (СПИД) и хантавирусный легочный синдром человека, вирусная геморрагическая болезнь кроликов, респираторно-репродуктивный синдром свиней, морбилливирусная чума тюленей Байкала и Северного моря, зоонотическая морбилливирусная респираторная болезнь лошадей в Австралии, и таксономические проблемы в этом контексте занимают одно из ведущих мест в решении связанных с ними сугубо практических задач. Кроме того, классификационная система весьма полезна как для студентов, так и для преподавателей, и привносит строгость в научную литературу по вирусологии (Ф. Мерфи).
2. ОБЩИЕ СВЕДЕНИЯ.
Современная универсальная система классификации вирусов использует иерархические уровни (таксоны), соответствующие порядку, семейству, подсемейству, роду и виду. Номенклатура по традиции заключается в том, что за словом вирус ставится название болезни основного хозяина, например, вирус чумы КРС. Название порядка оканчивается на -. virales, семейств - на . viridae, подсемейств - на . virinae, рода - на . virus.
В качестве примера приведем полную таксономию вируса вакцины:
Вид вируса - это понятие, представляющее группу штаммов из различных источников, или популяцию штаммов из одного источника, которые в целом обладают набором коррелирующих стабильных свойств, отделяющих данную группу от других групп штаммов. В 1991 году МКТВ принял следующее определение вид - это класс вирусов, характеризующийся большим числом критериев, образующий реплицирующуюся линию и занимающий особую экологическую нишу. Это определение учитывает вариабельность вирусов и не зависит от какого-либо единственного уникального параметра.
Род - это группа видов с определенными общими характеристиками и отличающаяся от вирусов других родов. Критерии для выделения родов различаются у разных семейств и могут включать:
- феномены генетических взаимодействий;
- круг восприимчивых хозяев;
Семейство - это группа родов с общими характеристиками, отличающаяся от в других семейств. Основные критерии, использующиеся для деления на семейства, включают фундаментальные свойства вирионов:
- тип и структуру нуклеиновой кислоты;
- наличие липопротеиновой оболочки;
- стратегию вирусного генома;
- размер и морфологию вирионов.
Учитывая, что семейство является наиболее крупным таксоном вирусов, кратко остановимся на характеристике упомянутых свойств вирионов.
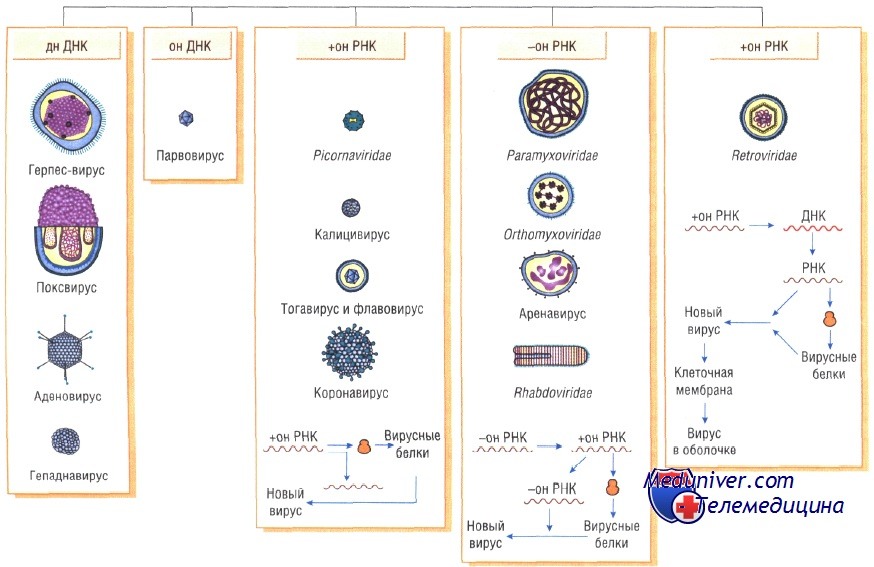
Тип и структура нуклеиновой кислоты. Вирусы содержат либо ДНК, либо РНК. И те, и другие могут быть двух- и односпиральными (дс- и ос-, соответственно). Единственное семейство зоопатогенных вирусов, содержащих ДНК с одной спиралью - Parvoviridae; остальные содержат дс-ДНК. Только представители семейств Reoviridae и Birnaviridae включают вирусы с дс-РНК, остальные зоопатогенные РНК- вирусы содержат односпиральный геном. Дополнительными структурными характеристиками нуклеиновых кислот, имеющими значение для классификации, являются сегментированность генома (Reoviridae -10 сегментов; Orthomyxoviridae -8; Bunyaviridae -3; Arenaviridae -2; Birnaviridae -2), его замкнутость в кольцо (Papovavioridae, Hepadnaviridae) или линейность.
Наличие липопротеиновой оболочки. Большинство вирусов имеют суперкапсидные оболочки, принципиально устроенные по типу мембран иных биологических структур. Не содержат суперкапсидных оболочек все зоопатогенные вирусы с ос-ДНК , дс-РНК и некоторые - с ос-РНК.
На основании двух первых критериев (типа, числа спиралей вирусных нуклеиновых кислот и наличия/отсутствия оболочки) зоопатогенные вирусы делят на 6 кластеров семейств (см. прилагаемую схему).
Стратегия вирусного генома. Под стратегией вирусного генома понимается способ репродукции, используемый вирусом. Реализация генетической информации идет по двум направлениям: во-первых, геном воспроизводит себя, то есть реплицируется; во-вторых, происходит синтез кодируемых геномом белков через процессы транскрипции/трансляции. Механизмы обоих направлений являются значимыми для таксономии. Так, вирусы с дс-ДНК могут реплицироваться по двум разным
классической [()ДНК()ДНК] для всех семейств, кроме Hepadnaviridae,;
по схеме ()ДНК(+)РНК()ДНК()ДНК для Hepadnaviridae.
Синтез вирусспецифических белков у РНК-вирусов так же происходит по-разному. У вирусов с позитивным геномом (Picornaviridae, Flaviviridae) геномная РНК может являться одновременно информационной (мРНК), которая и служит матрицей для синтеза одного белка-предшественника, далее протеолитически расщепляющегося на отдельные вирусные белки. У представителей Coronaviridae геномная РНК служит матрицей для синтеза РНК-полимеразы, которая в свою очередь синтезирует несколько мРНК разного размера, комплементарных геному. У Retroviridae синтез белка осуществляется по схеме РНКДНКмРНКбелок. У РНК-вирусов с негативным геномом (Orthomyxoviridae, Paramyxoviridae, Rhabdoviridae) транскрипция мРНК осуществляется вирионной РНК-полимеразой с использованием в качестве матрицы геномной РНК.
Размер и морфология вирионов. Размеры вирионов колеблятся от 15-20 нм (Circoviridae) до 300240100 нм (Poxviridae). По морфологии вирионы и капсиды делятся на 4 типа: сферические (S); удлиненые без закругленных концов (Е); удлиненные с закругленным одним или двумя концами (U); сложные (Х). Примеры характеристик различной морфологии вирусов (форма вириона/форма капсида):
S/S - Togaviridae, Flaviviridae;
S/E - Coronaviridae, Paramyxoviridae, Orthomyxoviridae, Retroviridae;
Общая характеристика структурной организации зоопатогенных вирусов приведена на прилагаемой схеме.
Подсемейства выделены в некоторых семействах вирусов (Poxviridae, Herpesviridae, Paramyxoviridae), объединяющих несколько родов.
Mononegavirales, включающее семейства Filoviridae, Paramyxoviridae, Rhabdoviridae, Bornaviridae;
Nidovirales, включающее семейства Arteriviridae и Coronaviridae.
структурная организация - морфология, размер, форма вириона и капсида, наличие/отсутствие пепломеров, их природа, наличие/отсутствие суперкапсидной оболочки, симметрия капсида, его структура;
физико-химические и физические свойства - м.м. вириона, плавучая плотность вириона ( в CsCl, сахарозе и т.д.), коэффициент седиментации вириона (S), стабильность при различных значениях рН и температуры, стабильность в присутствии катионов (Mg2+, Mn2+), устойчивость к органическим растворителям, детергентам, облучению;
белки - число, размер и функциональная активность структурных белков, число, размер и функциональная активность неструктурных белков, особенности специальной функциональной активности белков, прежде всего полимераз, обратной транскриптазы, гемагглютинина, нейраминидазы, белков слияния, аминокислотная последовательность, наличие гликозилированных, фосфорилированных, миристированных белков, антигенность;.
липиды - наличие, структурные особенности, локализация в стуктуре вириона;
стратегия генома - организация генома, стратегия репликации, число и положение открытых рамок считывания, характер транскрипции, характер трансляции, место накопления вирусных белков, их модификация, место сборки вирионов, место и природа созревания вируса, характер экзоцитоза;
система бактериальной номенклатуры не применима к вирусам;
номенклатура должна быть международной;
номенклатура должна быть универсально применимой ко всем вирусам;
существующие названия должны быть сохранены всякий раз, где это возможно;
закон приоритета не должен соблюдаться;
сокращения могут быть приняты в качестве групповых названий вирусов, если они имеют смысл для работающих в этой области и рекомендованы международной группой изучения;
персональные имена не используются;
имена должны иметь интернациональное значение;
правила орфографии имен существительных и прилагательных должны соответствовать установленным в специальном протоколе совещания МКТВ;
вид вирусам - это понятие, которое обычно представлено группой штаммов из различных источников, или популяций штаммов из одного источника, которые в целом обладают набором коррелирующих стабильных свойств, отделяющих группу от других групп штаммов;
название вируса вместе с обозначением штамма должно однозначно идентифицировать его без необходимости включать родовое или групповое имя;
название вируса должно нести смысловую нагрузку и состоять из любого числа слов;
можно использовать числа, буквы и их комбинации в качестве официальных обозначений там, где эти буквы и цифры уже широко используются для отдельного вируса;
вновь "сконструированные" серийные номера, буквы, комбинации не принимаются сами по себе для обозначения вида;
искусственно полученные лабораторные гибриды различных вирусов таксономией не рассматриваются;
одобрение МКТВ вновь предложенных названий видов происходит в две стадии. Сначала дается предварительное одобрение. Предварительно одобренные предложения публикуются в отчете МКТВ. Затем, после трехлетнего периода, предложение может быть окончательно одобрено МКТВ;
род - это группа видов с определенными близкими характеристиками;
название рода должно быть единственным смысловым словом, оканчивающимся на ". virus";
семейство - это группа родов с общими характеристиками с окончанием названия на ". viridae";
одобрение нового семейства должно быть связано с одобрением типового рода, одобрение нового рода должно быть связано с одобрением типового вида.
Критерии описания видов могут изменяться в различных семействах вирусов.
Везде, где можно, следует избегать повторения уже одобренных названий.
Если необходимо заменить типовой вид, в МКТВ подается таксономическая заявка.
В будущей номенклатуре следует избегать подстрочных, надстрочных обозначений, дефисов, косых полос, греческих букв.
РНК-вирусы как причина развития рака - вирусный канцерогенез
Открытие вирусов, вызывающих злокачественные опухоли у животных, произошло еще на рубеже XIX и XX веков. В 1910 г. Пейтон Рауш обнаружил, что бесклеточный фильтрат из тканей птичьей саркомы может вызывать развитие аналогичной саркомы у цыплят. Примерно в это же время была доказана вирусная природа птичьего миелобластоза. Позднее было обнаружено, что часто проходит весьма значительный латентный период между инфицированием вирусом и развитием рака.
Тем не менее вплоть до 1960-х годов не существовало четких доказательств того, что инкорпорация вирусной ДНК в геном клетки является необходимым условием развития злокачественной трансформации, как не было и случаев выделения вирусной ДНК из раковых клеток.
РНК-содержащие вирусы вызывают развитие целого ряда различных опухолей у животных, при этом наиболее распространена индукция этими вирусами лимфом, лейкемий и сарком. Типичная структура таких вирусов - это две идентичные цепочки молекул РНК в сочетании с ферментом обратная транскриптаза, одетых в гликопротеиновую оболочку. При инфицировании вирусом его обратная транскриптаза заставляет клетки синтезировать ДНК, комплементарную вирусной РНК.
Эта ДНК затем встраивается в клеточные хромосомы, и на ее основе клетка уже сама начинает синтез новых вирусных белков, вирусных обратных транскриптаз и элементов гликопротеиновой оболочки. Из-за механизма их действия этот тип вирусов получил название ретровирусы. Все они имеют весьма схожий внешний вид на электронных микрофотографиях и являются самыми маленькими из известных вирусов.
Некоторые из ретровирусов (например, вирусы птичьего лейкоза, кошачьей и мышиной лейкемии) содержат в себе только три гена и обладают очень длительным инкубационным периодом с момента заражения до возникновения опухоли. Другие вирусы (например, вирус саркомы Рауса (ВСР)) вызывают очень быструю злокачественную трансформацию и могут быть выделены из культуры опухолевых клеток.
Показано, что вирус ВСР содержит особый ген (v-src), способный вызывать трансформацию фибробластов in vitro. Этот ген кодирует наработку протеинкиназы, которая фосфорилирует тирозин. К сожалению, действие этой протеинкиназы запускает целый каскад различных метаболических процессов, и очень сложно оценить, какой именно из них ведет к злокачественной трансформации.
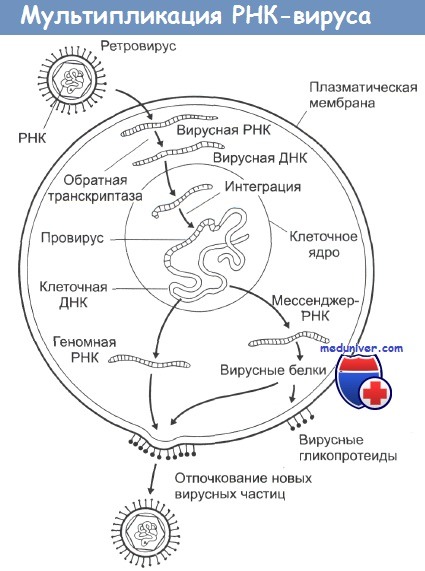
В настоящее время известно, что и нормальные, и злокачественные клетки содержат в своем генотипе участки ДНК, сходные или идентичные ряду последовательностей онкогенных РНК-содержащих вирусов. Такие участки получили название клеточных прото-онкогенов (чтобы отличать их от вирусных онкогенов). Постулируется, что активация этих участков, возникающая в результате канцерогенного воздействия, запускает целую цепочку событий, ведущую в итоге к злокачественной трансформации клетки. Считается также, что ретровирусы инкорпорировали эти клеточные участки в свой геном в процессе эволюции.
В настоящее время мы лучше понимаем механизмы действия продуктов вирусной активации. Примером такого продукта является протеинкиназа, активируемая геном sre, а также целый ряд других вирус-индуцированных канцерогенов. Это и рецепторы к эпидермальному фактору роста, продуцируемые геном v-erb, и тромбоцитарный фактор роста (ТФР), кодируемый фрагментами гена v-sis, и целый ряд связывающихся с ядром клетки белков, продукцию которых вызывает вирус птичьей лейкемии.
Как нормальные, так и раковые клетки содержат в себе участки последовательностей ДНК, гомологичные РНК онкогенных вирусов. Если происходит экспрессия или активация под действием канцерогенов этих клеточных онкогенов, это ведет к злокачественной трансформации клеток.
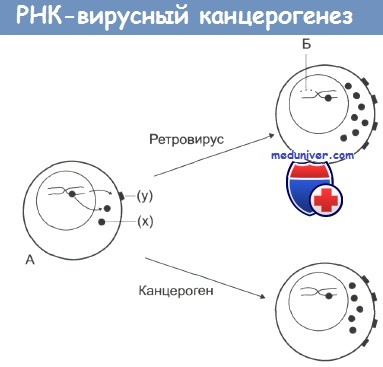
Онкогенная и злокачественная трансформация.
На стадии А нормальная клетка, которая характеризуется низкой протоонкогенной активностью, продуцирует фактор роста (х) или белки дифференцировки или рецепторы (у).
Канцерогены повышают активность протоонкогенов, что дает начало неопластической трансформации.
По другому механизму: при заражении ретровирусом происходит внедрение в ДНК клетки вирусных промоторов или онкогенов (Б), что также ведет к увеличению онкогенной активности и последующей злокачественной трансформации.
Первым ретровирусом, для которого была однозначно доказана связь со злокачественными новообразованиями, был вирус Т-клеточной лейкемии человека (ВТКЛ-1), выделенный из клеток хронической кожной Т-лимфомы. Этот вирус довольно широко распространен, может передаваться половым путем, через кровь, особенно в среде наркоманов, и от беременной женщины к плоду. Первоначально эндемический ареал циркуляции данного вируса был в основном представлен тропическими странами, однако в настоящее время в США серопозитивная реакция на вирус обнаруживается у каждого из 4000 человек населения. Кроме Т-клеточной лейкемии вирус вызывает тропический спастический паралич.
После 20 лет наблюдения за серопозитивными пациентами оценено, что риск развития последнего заболевания составляет около 5%. Один из вирусных генов, а именно tax-ген, вызывает увеличение продукции клеточного интерлейкина-2 (ИЛ-2) и его рецепторов, что является основным фактором, стимулирующим деление Т-клеток.
Ретровирусы могут вызывать опухолевые заболевания не напрямую, а опосредованно, как это было показано для вируса иммунодефицита человека (ВИЧ-1), вызывающего развитие СПИДа. Случаи развития рака у ВИЧ-инфицированных рассмотрены в одном из обзоров. Отмечается, что у ВИЧ-инфицированных наиболее часто развиваются три типа опухолей: мгновенная или высокозлокачественная В-клеточная лимфома; саркома Капоши (СК, которую вызывает другой вирус — герпесвирус ГСК, или герпесвирус 8); карцинома шейки матки.
До разработки эффективных методов терапии ВИЧ-инфицированных более 40% из них заболевали тем или иным видом рака. Тем не менее связь этого вируса с развитием онкологических заболеваний скорее всего косвенная и опосредована развитием общей хронической иммунодепрессии организма, которая и позволяет другим канцерогенным вирусам вызывать рак. В-клеточные лимфомы также характеризуются довольно сложным патогенезом. Хотя В-клетки не поражаются вирусом ВИЧ-1, они могут быть мишенью для других типов вирусов, например для вируса Эпштейна-Барра (ВЭБ). Рак шейки матки у женщин также развивается на основе вторичной вирусной инфекции — вируса папилломы человека (ВПЧ), на фоне общей иммунодепрессии, вызванной ВИЧ-1. По причине общего снижения иммунных реакций организма все эти опухоли развиваются особенно быстро и агрессивно.
Показано, что РНК-содержащий вирус гепатита С значительно увеличивает риск заболевания гепатоклеточной карциномой. У больных, инфицированных этим вирусом, риск развития раковой опухоли возрастает в 100 раз, но механизмы его канцерогенного действия в настоящее время почти не изучены. Если больной заражен еще и вирусом гепатита В, риск развития рака печени возрастает многократно. На земном шаре одним или обоими из этих вирусов заражено около миллиарда человек.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Строение вирусов. Классификация вирусов
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
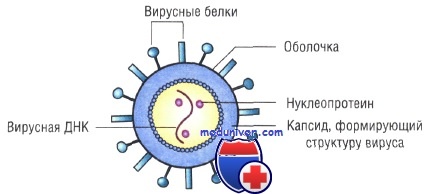
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Репликация вирусов. Размножение вирусов.
Биотехнологии в вирусологии.
Методология выращивания вирусов.
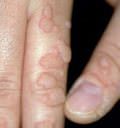
Культивирование вирусов. Антигены вирусов и иммуннитет.
Иммунная система при вирусной инфекции.
Патогенез и механизмы противовирусной защиты организма.
Развитие и образование противовирусного иммунитета.

Вакцинопрофилактика вирусных инфекций.
Физические методы инактивации вирусов для вакцин.
Живые вакцины. Гетерологичные вакцины. Субъединичные вакцины.

Современные субъединированные и рекомбинантные вакцины.

Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: