
Мейоз происходит при размножении вируса гриппа
Обновлено: 24.04.2024
Фазы мейоза половых клеток. Особенности
Мейоз — уникальный для половых клеток процесс, в котором диплоидные клетки порождают гаплоидные гаметы. Мейоз состоит из одного цикла синтеза ДНК и двух циклов расхождения хромосом и деления клетки. Способные к мейозу половые клетки — первичные сперматоциты или первичные овоциты — до наступления мейоза проходят через длинную серию митозов, начиная от зиготы.
Мужские и женские гаметы имеют разные истории; и хотя последовательность событий одинаковая, синхронизация весьма различна. Два последовательных мейотических деления называются мейозом I и мейозом II. Мейоз I так же известен как редукционное деление, поскольку число хромосом уменьшается наполовину вследствие спаривания гомологов в профазе и их расхождения в разные клетки в анафазе. Х- и Y-хромосомы не являются гомологами в строгом смысле, однако имеют гомологичные сегменты на концах коротких и длинных плеч, которыми они конъюгируют в ходе мейоза I.
Мейоз I также примечателен тем, что в нем происходит генетическая рекомбинация, называемая мейотическим кроссинговером.
В ходе этого процесса обмениваются гомологичные сегменты ДНК между разными, несестринскими хроматидами пары гомологичных хромосом. Это приводит к тому, что ни одна из гамет, полученных в результате мейоза, не идентична другой. Рекомбинация — фундаментальное понятие для процесса распределения генов, ответственных за наследственные болезни.
Поскольку рекомбинация предполагает физическое взаимодействие двух гомологичных хромосом в соответствующей точке в течение мейоза I, она также определяет правильность расхождения хромосом в мейозе. Нарушения в процессе рекомбинации могут вызвать нерасхождение хромосом в ходе мейоза I, самую частую причину хромосомных аномалий типа синдрома Дауна.
Мейоз II следует за мейозом I без промежуточного удвоения ДНК. Как и при обычном митозе, хроматиды расходятся и одна хроматида каждой хромосомы переходит в дочернюю клетку.
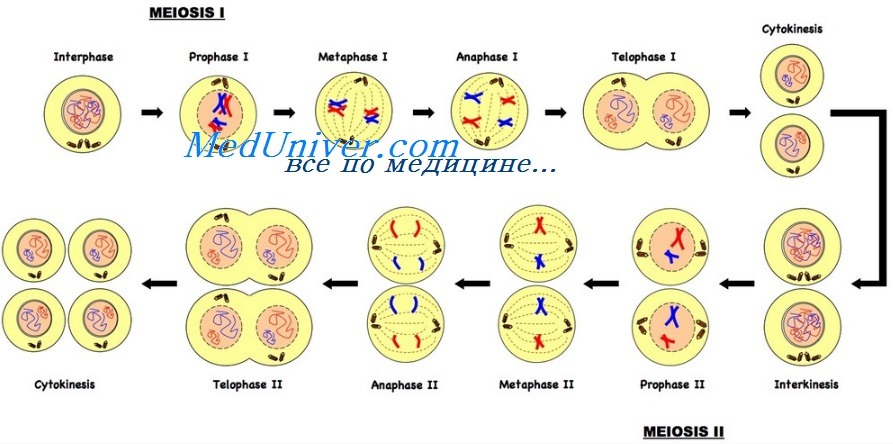
Первое мейотическое деление (мейоз I)
Профаза I мейоза. Профаза мейоза I — сложный процесс, который серьезно отличается от митотической профазы, с важными генетическими последствиями. Выделяют несколько этапов профазы. На всех этапах хромосомы непрерывно конденсируются и становятся короче и толще.
• Лептотена. Хромосомы, уже скопированные в ходе предыдущей S фазы, становятся видимыми как нити, начинается конденсация хроматина. Две однотипных хроматиды каждой хромосомы так тесно сближаются, что их невозможно выделить.
• Зиготена. Гомологичные хромосомы начинают выстраиваться и соединяться вдоль оси. Процесс спаривания, или синапсис, обычно очень точный, так что последовательности ДНК соответствуют друг другу на протяжении всей хромосомы. Хотя молекулярная основа синапсиса не до конца понятна, электронная микроскопия показывает, что хромосомы удерживаются вместе синаптонемальным комплексом — лентообразной белоксодержащей структурой. Синаптонемальный комплекс необходим для процесса рекомбинации.
• Пахитена. Хромосомы становятся более толстыми. Синапсис завершен, и каждая пара гомологов видна как бивалент (иногда называемый тетрадой, поскольку он содержит четыре хроматиды). Пахитена — этап, в котором происходит мейотический кроссинговер.
• Диплотена. После рекомбинации синаптонемальный комплекс начинает разрушаться, и два компонента каждого бивалента начинают отделиться друг от друга. В конце концов два гомолога каждого бивалента касаются друг друга только в точках, называемых хиазмами (пересечениями). Полагают, что они обозначают точки обмена. Среднее число хиазм, наблюдаемых в сперматоцитах, — около 50, т.е. несколько на каждый бивалент.
• Диакинез. На этом этапе хромосомы достигают максимальной конденсации. Метафаза I мейоза. Метафаза I начинается, как и в митозе, когда исчезает ядерная мембрана. Формируется веретено деления и спаренные хромосомы выстраиваются в плоскости экватора клетки, ориентируя центромеры к разным полюсам. Анафаза I мейоза. Компоненты каждого бивалента движутся независимо, а их центромеры с прикрепленными сестринскими хроматидами расходятся к противоположным полюсам клетки.
Этот процесс называют расхождением. Таким образом, число хромосом уменьшается вдвое, и каждая клетка, полученная в результате первого деления мейоза, получает гаплоидное число хромосом. Разные биваленты расходятся независимо друг от друга и в результате исходные отцовский и материнский хромосомные комплекты сортируются в произвольных комбинациях. Возможное количество комбинаций 23 хромосом, которое может образоваться в гаметах, — 223 (более 8 млн). Фактически же вариабельность генетического материала, передающегося от родителей ребенку, значительно больше, что обеспечивается кроссинговером.
В результате этого процесса каждая хроматида обычно содержит сегменты, производные от каждой родительской хромосомной пары; на этом этапе, например, типичная хромосома 1 формируется из трех-пяти сегментов, поочередно отцовского и материнского происхождения. В процессе деления клетки может происходить много ошибок. Некоторые заканчиваются остановкой мейоза и гибелью клетки, другие ведут к неправильному расхождению хромосом в анафазе. Например, оба гомолога хромосомной пары могут переместиться к одному и тому же, а не противоположным полюсам в анафазе мейоза I. Этот патологический процесс называется нерасхождением. Телофаза I мейоза. В телофазе гаплоидные комплекты хромосом группируются в противоположных полюсах клетки.
Цитокинез. После телофазы I клетка делится на две гаплоидные дочерние клетки и входит в мейотическую интерфазу. При сперматогенезе цитоплазма более или менее одинаково делится между двумя дочерними клетками; но при овогенезе одна (вторичный овоцит) получает почти всю цитоплазму, а вторая клетка становится первым полярным тельцем. В отличие от митоза, интерфаза очень короткая, и сразу начинается второе мейотическое деление. Следует обратить внимание на существенное различие между мейотической и митотической интерфазами — отсутствие S-фазы (т.е. синтеза ДНК) между первым и вторым мейотическими делениями.
Второе мейотическое деление (мейоз II)
Второе мейотическое деление подобно обычному митозу, за исключением того, что набор хромосом, получаемый в результате мейоза II, — гаплоидный. Конечный результат мейоза — две дочерних клетки мейоза I — делятся, формируя четыре гаплоидных клетки, каждая из которых содержит 23 хромосомы. Как уже упоминалось, из-за кроссинговера в мейозе I хромосомы результирующих гамет неидентичны.
Подобно тому, как каждая родительская хромосома в паре произвольно и независимо переходит к дочерним клеткам в мейозе I, в ходе мейоза также произвольно распределяются отцовские и материнские аллели каждого гена. Тем не менее от того, в первом или втором делении мейоза произошло разделение аллелей, зависит, участвовали ли они в процессе кроссинговера в ходе первого мейотического деления.
Генетические последствия мейоза:
• Уменьшение числа хромосом от диплоидного до гаплоидного, необходимое для образования гамет.
• Сегрегация аллелей в первом и втором делении мейоза в соответствии с первым законом Менделя.
• Случайное перераспределение генетического материала в гомологичных хромосомах в соответствии со вторым законом Менделя.
• Дополнительное перераспределение генетического материала с помощью кроссинговера, значительно увеличивающее число генетических вариантов, а также играющее важную роль в процессе нормального расхождения хромосом.
Учебное видео: мейоз и его фазы
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Фазы мейоза половых клеток. Особенности
Мейоз — уникальный для половых клеток процесс, в котором диплоидные клетки порождают гаплоидные гаметы. Мейоз состоит из одного цикла синтеза ДНК и двух циклов расхождения хромосом и деления клетки. Способные к мейозу половые клетки — первичные сперматоциты или первичные овоциты — до наступления мейоза проходят через длинную серию митозов, начиная от зиготы.
Мужские и женские гаметы имеют разные истории; и хотя последовательность событий одинаковая, синхронизация весьма различна. Два последовательных мейотических деления называются мейозом I и мейозом II. Мейоз I так же известен как редукционное деление, поскольку число хромосом уменьшается наполовину вследствие спаривания гомологов в профазе и их расхождения в разные клетки в анафазе. Х- и Y-хромосомы не являются гомологами в строгом смысле, однако имеют гомологичные сегменты на концах коротких и длинных плеч, которыми они конъюгируют в ходе мейоза I.
Мейоз I также примечателен тем, что в нем происходит генетическая рекомбинация, называемая мейотическим кроссинговером.
В ходе этого процесса обмениваются гомологичные сегменты ДНК между разными, несестринскими хроматидами пары гомологичных хромосом. Это приводит к тому, что ни одна из гамет, полученных в результате мейоза, не идентична другой. Рекомбинация — фундаментальное понятие для процесса распределения генов, ответственных за наследственные болезни.
Поскольку рекомбинация предполагает физическое взаимодействие двух гомологичных хромосом в соответствующей точке в течение мейоза I, она также определяет правильность расхождения хромосом в мейозе. Нарушения в процессе рекомбинации могут вызвать нерасхождение хромосом в ходе мейоза I, самую частую причину хромосомных аномалий типа синдрома Дауна.
Мейоз II следует за мейозом I без промежуточного удвоения ДНК. Как и при обычном митозе, хроматиды расходятся и одна хроматида каждой хромосомы переходит в дочернюю клетку.
Первое мейотическое деление (мейоз I)
Профаза I мейоза. Профаза мейоза I — сложный процесс, который серьезно отличается от митотической профазы, с важными генетическими последствиями. Выделяют несколько этапов профазы. На всех этапах хромосомы непрерывно конденсируются и становятся короче и толще.
• Лептотена. Хромосомы, уже скопированные в ходе предыдущей S фазы, становятся видимыми как нити, начинается конденсация хроматина. Две однотипных хроматиды каждой хромосомы так тесно сближаются, что их невозможно выделить.
• Зиготена. Гомологичные хромосомы начинают выстраиваться и соединяться вдоль оси. Процесс спаривания, или синапсис, обычно очень точный, так что последовательности ДНК соответствуют друг другу на протяжении всей хромосомы. Хотя молекулярная основа синапсиса не до конца понятна, электронная микроскопия показывает, что хромосомы удерживаются вместе синаптонемальным комплексом — лентообразной белоксодержащей структурой. Синаптонемальный комплекс необходим для процесса рекомбинации.
• Пахитена. Хромосомы становятся более толстыми. Синапсис завершен, и каждая пара гомологов видна как бивалент (иногда называемый тетрадой, поскольку он содержит четыре хроматиды). Пахитена — этап, в котором происходит мейотический кроссинговер.
• Диплотена. После рекомбинации синаптонемальный комплекс начинает разрушаться, и два компонента каждого бивалента начинают отделиться друг от друга. В конце концов два гомолога каждого бивалента касаются друг друга только в точках, называемых хиазмами (пересечениями). Полагают, что они обозначают точки обмена. Среднее число хиазм, наблюдаемых в сперматоцитах, — около 50, т.е. несколько на каждый бивалент.
• Диакинез. На этом этапе хромосомы достигают максимальной конденсации. Метафаза I мейоза. Метафаза I начинается, как и в митозе, когда исчезает ядерная мембрана. Формируется веретено деления и спаренные хромосомы выстраиваются в плоскости экватора клетки, ориентируя центромеры к разным полюсам. Анафаза I мейоза. Компоненты каждого бивалента движутся независимо, а их центромеры с прикрепленными сестринскими хроматидами расходятся к противоположным полюсам клетки.
Этот процесс называют расхождением. Таким образом, число хромосом уменьшается вдвое, и каждая клетка, полученная в результате первого деления мейоза, получает гаплоидное число хромосом. Разные биваленты расходятся независимо друг от друга и в результате исходные отцовский и материнский хромосомные комплекты сортируются в произвольных комбинациях. Возможное количество комбинаций 23 хромосом, которое может образоваться в гаметах, — 223 (более 8 млн). Фактически же вариабельность генетического материала, передающегося от родителей ребенку, значительно больше, что обеспечивается кроссинговером.
В результате этого процесса каждая хроматида обычно содержит сегменты, производные от каждой родительской хромосомной пары; на этом этапе, например, типичная хромосома 1 формируется из трех-пяти сегментов, поочередно отцовского и материнского происхождения. В процессе деления клетки может происходить много ошибок. Некоторые заканчиваются остановкой мейоза и гибелью клетки, другие ведут к неправильному расхождению хромосом в анафазе. Например, оба гомолога хромосомной пары могут переместиться к одному и тому же, а не противоположным полюсам в анафазе мейоза I. Этот патологический процесс называется нерасхождением. Телофаза I мейоза. В телофазе гаплоидные комплекты хромосом группируются в противоположных полюсах клетки.
Цитокинез. После телофазы I клетка делится на две гаплоидные дочерние клетки и входит в мейотическую интерфазу. При сперматогенезе цитоплазма более или менее одинаково делится между двумя дочерними клетками; но при овогенезе одна (вторичный овоцит) получает почти всю цитоплазму, а вторая клетка становится первым полярным тельцем. В отличие от митоза, интерфаза очень короткая, и сразу начинается второе мейотическое деление. Следует обратить внимание на существенное различие между мейотической и митотической интерфазами — отсутствие S-фазы (т.е. синтеза ДНК) между первым и вторым мейотическими делениями.
Второе мейотическое деление (мейоз II)
Второе мейотическое деление подобно обычному митозу, за исключением того, что набор хромосом, получаемый в результате мейоза II, — гаплоидный. Конечный результат мейоза — две дочерних клетки мейоза I — делятся, формируя четыре гаплоидных клетки, каждая из которых содержит 23 хромосомы. Как уже упоминалось, из-за кроссинговера в мейозе I хромосомы результирующих гамет неидентичны.
Подобно тому, как каждая родительская хромосома в паре произвольно и независимо переходит к дочерним клеткам в мейозе I, в ходе мейоза также произвольно распределяются отцовские и материнские аллели каждого гена. Тем не менее от того, в первом или втором делении мейоза произошло разделение аллелей, зависит, участвовали ли они в процессе кроссинговера в ходе первого мейотического деления.
Генетические последствия мейоза:
• Уменьшение числа хромосом от диплоидного до гаплоидного, необходимое для образования гамет.
• Сегрегация аллелей в первом и втором делении мейоза в соответствии с первым законом Менделя.
• Случайное перераспределение генетического материала в гомологичных хромосомах в соответствии со вторым законом Менделя.
• Дополнительное перераспределение генетического материала с помощью кроссинговера, значительно увеличивающее число генетических вариантов, а также играющее важную роль в процессе нормального расхождения хромосом.
Учебное видео: мейоз и его фазы
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Гаметогенез - мейоз: фазы, особенности у мужчин, женщин
Процесс образования гаплоидного набора из диплоидного, происходящий при формировании половых клеток, называют мейозом. В пересчёте на количество центромер происходит сначала редукционное деление (мейоз I), а затем эквационное деление (мейоз II). У мужчин мейоз проходит так же, как и у большинства диплоидных видов, а у женщин данный процесс имеет некоторые отличия.
Кроссинговер между хромосомами отца и матери обеспечивает перегруппировку генетической информации между поколениями. Во время оплодотворения происходит слияние гаплоидных наборов хромосом сперматозоида и яйцеклетки, таким образом в зиготе восстанавливается диплоидный набор.
Мейоз I
Мейоз I имеет много общего с митозом, однако это более сложный и продолжительный процесс.
Мейоз первичных сперматоцитов и овоцитов начинается после фазы G2 митоза, а потому они имеют диплоидный набор хромосом (2N), содержащих реплицированную ДНК в составе сестринских хроматид (4С). Профаза I включает обоюдный обмен между хроматидами матери и отца при помощи крос-синговера.
Профаза I
• Лептотена. Хромосомы представлены в виде длинных нитей, прикреплённых концами к ядерной оболочке.
• Зиготена. Хромосомы сокращаются, образуют пары и гомологи слипаются друг с другом (синапсис). Данный процесс характеризует точное совмещение хромосом (ген к гену на протяжении всего генома). При этом у первичных сперматоцитов хромосомы X и Y образуют синапсис только концами своих коротких плечей.
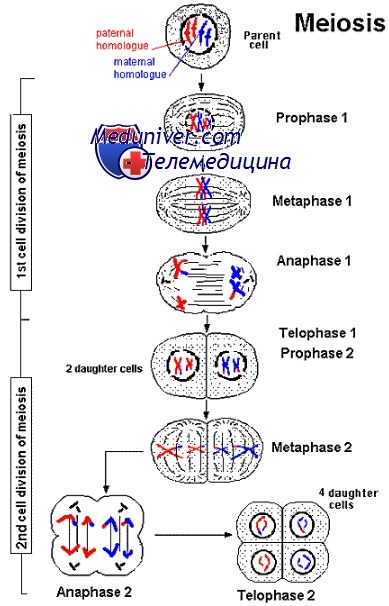
• Пахитена. Сестринские хроматиды начинают разделяться. Пары гомологичных хромосом, называемые бивалентами, имеют по четыре двойных спирали ДНК (тетрада). Одна или обе хроматиды каждой из отцовских хромосом скрещиваются с материнскими и образуют синаптонемальный комплекс. Каждая пара хромосом претерпевает хотя бы один кроссинговер.
• Диплотена. Происходит разделение хроматид, за исключением участков кроссинговера, или хиазм. Хромосомы всех первичных овоцитов находятся в таком состоянии вплоть до овуляции.
• Диакинез. Реорганизованные хромосомы начинают расходиться. В этот момент каждый бивалент содержит четыре хроматиды, соединённые обыкновенными центромерами, и несестринские хроматиды, соединённые хиазмами.
Метафаза I, анафаза 1, тепофаза 1, цитокинез I
Данные стадии мейоза подобны фазам митоза. Основное отличие: вместо разъединения несестринских хроматид происходит распределение по дочерним клеткам парных кроссоверных сестринских хроматид, соединённых центромерами.
В конце мейоза I вторичные сперматоциты и овоциты имеют 23 хромосомы (1N), каждая из которых состоит из двух хроматид (2С).
Мейоз II
При мейозе II возникает кратковременная интерфаза, во время которой не происходит репликации хромосом. Затем следуют профаза, метафаза, анафаза, телофаза и цитокинез. Схожесть каждой фазы мейоза II с подобной ей при митозе заключается в том, что пары хроматид (биваленты), соединённые в области центромер, выстраиваются в линию и образуют метафазную пластинку, а затем расходятся по дочерним клеткам, после чего следует репликация ДНК центромер.
В конце мейоза II в клетках содержится 23 хромосомы (IN), каждая из которых состоит из одной хроматиды (1С).
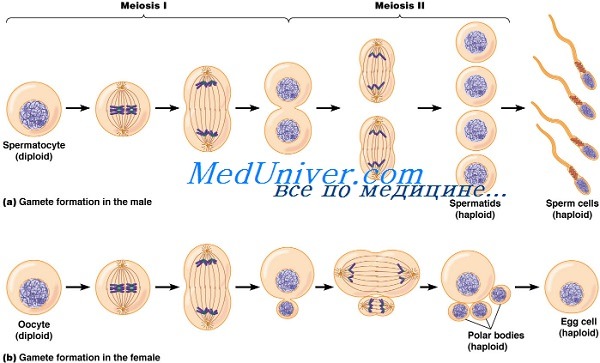
Мейоз у мужчин
Сперматогенезом называют процесс длительностью до 64 дней, включающий все стадии, на протяжении которых сперматогоний превращается в сперматозоид. При этом цитокинез остаётся незавершённым, позволяя каждому поколению клеток быть связанным цитоплазматическими мостиками.
После того как диплоидный первичный сперматоцит проходит стадию мейоза I, возникают два гаплоидных вторичных сперматоцита. Затем следует мейоз II, в результате которого появляются четыре гаплоидные спсрматиды. Во время спермиогенеза сперматиды превращаются в сперматозоиды. Данный процесс включает:
— образование акросомы, содержащей ферменты, которые способствуют проникновению семени;
— конденсацию ядра;
— удаление большей части цитоплазмы;
— формирование шейки, средней части и хвоста.
Мейоз у женщин
Овогенез начинается у плода в возрасте 12 недель и внезапно прекращается к 20-й неделе. Первичные овоциты остаются в форме диплотены профазы I вплоть до овуляции. Данную стадию называют диктиотеной.
Обычно созревает не более одного овоцита в месяц. Под влиянием гормонов первичный овоцит набухает, накапливая цитоплазматический материал. По завершению мейоза I он наследуется одной дочерней клеткой — вторичным овоцитом. Второе ядро переходит в первое направительное тельце, которое обычно не делится и со временем дегенерирует. После окончания мейоза I вторичный овоцит попадает в матку или фаллопиевы трубы.
Мейоз II вторичного овоцита останавливается на стадии метафазы до попадания в него сперматозоида. После этого процесс деления завершается, и образуется большой гаплоидный пронуклеус яйцеклетки, который сливается с пронуклеусом сперматозоида, а также второе направительное тельце, которое дегенерирует.
В зависимости от того, когда произойдёт оплодотворение, продолжительность данного процесса составляет 12—50 лет.
Медицинское значение понимания мейоза
• Диплоидный набор хромосом соматических клеток снижается до гаплоидного в половых клетках.
• Отцовские и материнские хромосомы подвергаются пересортировке, в результате чего количество возможных комбинаций (за исключением рекомбинаций внутри самих хромосом) возрастает до 223 (8 388 608).
• Пересортировка отцовских и материнских аллелей внутри хромосом создаёт между гаметами бесконечное количество генетических вариаций.
• Случайность процесса пересортировки отцовских и материнских аллелей во время мейоза (и оплодотворения) позволяет применять теорию вероятностей к генетическим пропорциям и генетической изменчивости согласно законам Менделя.
• Частота кроссинговера внутри хромосом позволяет предположить относительное расположение того или иного гена.
• Ошибки, возникающие во время конъюгации хромосом и кроссинговера, могут вызвать транслокации, разделение или расхождение хромосом, что часто становится причиной анеуплоидии.
Учебное видео: мейоз и его фазы

Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Вернуться в содержание раздела "генетика" на нашем сайте
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Обзор
Автор
Редакторы
Обратите внимание!
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science.
Эволюция и происхождение вирусов
В 2007 году сотрудники биологического факультета МГУ Л. Нефедова и А. Ким описали, как мог появиться один из видов вирусов — ретровирусы. Они провели сравнительный анализ геномов дрозофилы D. melanogaster и ее эндосимбионта (микроорганизма, живущего внутри дрозофилы) — бактерии Wolbachia pipientis. Полученные данные показали, что эндогенные ретровирусы группы gypsy могли произойти от мобильных элементов генома — ретротранспозонов. Причиной этому стало появление у ретротранспозонов одного нового гена — env, — который и превратил их в вирусы. Этот ген позволяет вирусам передаваться горизонтально, от клетки к клетке и от носителя к носителю, чего ретротранспозоны делать не могли. Именно так, как показал анализ, ретровирус gypsy передался из генома дрозофилы ее симбионту — вольбахии [7]. Это открытие упомянуто здесь не случайно. Оно нам понадобится для того, чтобы понять, чем вызваны трудности борьбы с вирусами.
Из давних письменных источников, оставленных историком Фукидидом и знахарем Галеном, нам известно о первых вирусных эпидемиях, возникших в Древней Греции в 430 году до н.э. и в Риме в 166 году. Часть вирусологов предполагает, что в Риме могла произойти первая зафиксированная в источниках эпидемия оспы. Тогда от неизвестного смертоносного вируса по всей Римской империи погибло несколько миллионов человек [8]. И с того времени европейский континент уже регулярно подвергался опустошающим нашествиям всевозможных эпидемий — в первую очередь, чумы, холеры и натуральной оспы. Эпидемии внезапно приходили одна за другой вместе с перемещавшимися на дальние расстояния людьми и опустошали целые города. И так же внезапно прекращались, ничем не проявляя себя сотни лет.
Вирус натуральной оспы стал первым инфекционным носителем, который представлял действительную угрозу для человечества и от которого погибало большое количество людей. Свирепствовавшая в средние века оспа буквально выкашивала целые города, оставляя после себя огромные кладбища погибших. В 2007 году в журнале Национальной академии наук США (PNAS) вышла работа группы американских ученых — И. Дэймона и его коллег, — которым на основе геномного анализа удалось установить предположительное время возникновения вируса натуральной оспы: более 16 тысяч лет назад. Интересно, что в этой же статье ученые недоумевают по поводу своего открытия: как так случилось, что, несмотря на древний возраст вируса, эпидемии оспы не упоминаются в Библии, а также в книгах древних римлян и греков [9]?
Строение вирусов и иммунный ответ организма

Рисунок 1. Первооткрыватель вирусов Д.И. Ивановский (1864–1920) (слева) и английский врач Эдвард Дженнер (справа).

Почти все известные науке вирусы имеют свою специфическую мишень в живом организме — определенный рецептор на поверхности клетки, к которому и прикрепляется вирус. Этот вирусный механизм и предопределяет, какие именно клетки пострадают от инфекции. К примеру, вирус полиомиелита может прикрепляться лишь к нейронам и потому поражает именно их, в то время как вирусы гепатита поражают только клетки печени. Некоторые вирусы — например, вирус гриппа А-типа и риновирус — прикрепляются к рецепторам гликофорин А и ICAM-1, которые характерны для нескольких видов клеток. Вирус иммунодефицита избирает в качестве мишеней целый ряд клеток: в первую очередь, клетки иммунной системы (Т-хелперы, макрофаги), а также эозинофилы, тимоциты, дендритные клетки, астроциты и другие, несущие на своей мембране специфический рецептор СD-4 и CXCR4-корецептор [13–15].

Одновременно с этим в организме реализуется еще один, молекулярный, защитный механизм: пораженные вирусом клетки начинают производить специальные белки — интерфероны, — о которых многие слышали в связи с гриппозной инфекцией. Существует три основных вида интерферонов. Синтез интерферона-альфа (ИФ-α) стимулируют лейкоциты. Он участвует в борьбе с вирусами и обладает противоопухолевым действием. Интерферон-бета (ИФ-β) производят клетки соединительной ткани, фибробласты. Он обладает таким же действием, как и ИФ-α, только с уклоном в противоопухолевый эффект. Интерферон-гамма (ИФ-γ) синтезируют Т-клетки (Т-хелперы и (СD8+) Т-лимфоциты), что придает ему свойства иммуномодулятора, усиливающего или ослабляющего иммунитет. Как именно интерфероны борются с вирусами? Они могут, в частности, блокировать работу чужеродных нуклеиновых кислот, не давая вирусу возможности реплицироваться (размножаться).

Причины поражений в борьбе с ВИЧ
Тем не менее нельзя сказать, что ничего не делается в борьбе с ВИЧ и нет никаких подвижек в этом вопросе. Сегодня уже определены перспективные направления в исследованиях, главные из которых: использование антисмысловых молекул (антисмысловых РНК), РНК-интерференция, аптамерная и химерная технологии [12]. Но пока эти антивирусные методы — дело научных институтов, а не широкой клинической практики*. И потому более миллиона человек, по официальным данным ВОЗ, погибают ежегодно от причин, связанных с ВИЧ и СПИДом.

Подобный вирусный механизм характерен не только для ВИЧ. Он описан и при инфицировании некоторыми другими опасными вирусами: такими, как вирусы Денге и Эбола. Но при ВИЧ антителозависимое усиление инфекции сопровождается еще несколькими факторами, делая его опасным и почти неуязвимым. Так, в 1991 году американские клеточные биологи из Мэриленда (Дж. Гудсмит с коллегами), изучая иммунный ответ на ВИЧ-вакцину, обнаружили так называемый феномен антигенного импринтинга [23]. Он был описан еще в далеком 1953 году при изучении вируса гриппа. Оказалось, что иммунная система запоминает самый первый вариант вируса ВИЧ и вырабатывает к нему специфические антитела. Когда вирус видоизменяется в результате точечных мутаций, а это происходит часто и быстро, иммунная система почему-то не реагирует на эти изменения, продолжая производить антитела к самому первому варианту вируса. Именно этот феномен, как считает ряд ученых, стоит препятствием перед созданием эффективной вакцины против ВИЧ.

Открытие биологов из МГУ — Нефёдовой и Кима, — о котором упоминалось в самом начале, также говорит в пользу этой, эволюционной, версии.

Сегодня не только ВИЧ представляет опасность для человечества, хотя он, конечно, самый главный наш вирусный враг. Так сложилось, что СМИ уделяют внимание, в основном, молниеносным инфекциям, вроде атипичной пневмонии или МЕRS, которыми быстро заражается сравнительно большое количество людей (и немало гибнет). Из-за этого в тени остаются медленно текущие инфекции, которые сегодня гораздо опаснее и коварнее коронавирусов* и даже вируса Эбола. К примеру, мало кто знает о мировой эпидемии гепатита С, вирус которого был открыт в 1989 году**. А ведь по всему миру сейчас насчитывается 150 млн человек — носителей вируса гепатита С! И, по данным ВОЗ, каждый год от этой инфекции умирает 350-500 тысяч человек [33]. Для сравнения — от лихорадки Эбола в 2014-2015 гг. (на состояние по июнь 2015 г.) погибли 11 184 человека [34].
* — Коронавирусы — РНК-содержащие вирусы, поверхность которых покрыта булавовидными отростками, придающими им форму короны. Коронавирусы поражают альвеолярный эпителий (выстилку легочных альвеол), повышая проницаемость клеток, что приводит к нарушению водно-электролитного баланса и развитию пневмонии.

Рисунок 8. Электронная микрофотография воссозданного вируса H1N1, вызвавшего эпидемию в 1918 г. Рисунок с сайта phil.cdc.gov.
Почему же вдруг сложилась такая ситуация, что буквально каждый год появляются новые, всё более опасные формы вирусов? По мнению ученых, главные причины — это сомкнутость популяции, когда происходит тесный контакт людей при их большом количестве, и снижение иммунитета вследствие загрязнения среды обитания и стрессов. Научный и технический прогресс создал такие возможности и средства передвижения, что носитель опасной инфекции уже через несколько суток может добраться с одного континента на другой, преодолев тысячи километров.

Чтобы жизненный цикл эукариот, обладающих половым процессом, мог продолжаться, должен был развиться механизм, посредством которого из диплоидных клеток снова могли образовываться гаплоидные. Таким механизмом стал мейоз – особый вид клеточного деления, при котором число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой.
В статье Ю.Ф.Богданова, крупнейшего специалиста по эволюции мейоза, рассматриваются современные представления о происхождении механизмов мейотического деления у эукариот.

В ходе митоза (а также в ходе второго деления мейоза) хромосомы вступают в метафазу поодиночке, и нити веретена деления присоединяются к каждой хромосоме с двух сторон.
Именно благодаря указанным особенностям первого деления мейоза и обеспечивается уменьшение числа хромосом: к полюсам клетки расходятся не сестринские хроматиды, а гомологичные хромосомы, по одной из каждой пары (см. схему).
Эти ключевые особенности первого деления мейоза развились на основе одного и того же ароморфоза – возникновения так называемого синаптонемного комплекса (СК).
Одним из этих исходных блоков стал уже имевшийся к тому времени у эукариот механизм митоза, другим – механизм репарации (починки повреждений) ДНК, основанный на гомологичной рекомбинации (см.: Глазер В.М. Гомологичная генетическая рекомбинация ). Суть процесса в том, что поврежденный участок одной молекулы ДНК заменяется его неповрежденной копией, взятой из другой (гомологичной) молекулы ДНК.

Упрощенная схема одного из вариантов гомологичной рекомбинации (обмена участками между гомологичными молекулами ДНК) с образованием соединения Холлидея. Показаны две двухцепочечные молекулы ДНК. Тут важно не перепутать: каждая молекула ДНК состоит из двух параллельных цепей нуклеотидов (двойная спираль); в состав одной хроматиды входит одна молекула ДНК; каждая хромосома (вплоть до анафазы II) состоит из двух хроматид. Соответственно, каждая пара гомологичных хромосом (например, в метафазе I) состоит из ДВУХ хромосом, ЧЕТЫРЕХ хроматид, ЧЕТЫРЕХ молекул ДНК, ВОСЬМИ нуклеотидных цепочек. Рекомбинация происходит между хроматидами отцовской и материнской хромосом. На этом рисунке показана рекомбинация у одного из вирусов, у эукариот во время мейоза все несколько иначе и сложнее
Починка разрывов ДНК – одна из основ мейоза
Механизмы репарации и рекомбинации ДНК – очень древние, они должны были возникнуть еще на заре жизни.
Жизненный цикл древних эукариот (как и многих прокариот), вероятно, состоял из двух фаз и двух соответствующих типов метаболизма: 1) вегетативная фаза (условия благоприятны, клетки размножаются митозом); 2) фаза споруляции (условия стали неблагоприятными, клетки превращаются в споры). У низших эукариот – например, у дрожжей, - перед споруляцией происходит мейоз, и из получившихся гаплоидных клеток образуются споры.
Становление мейоза в ходе эволюции, так же как и сам мейоз в жизненном цикле современных организмов, начинается с репарации повреждений (разрывов) ДНК. Разрывы эти в начале мейоза возникают не сами собой – их создает специальный фермент, эндонуклеаза SPO11. Этот фермент является модификацией другого белка – ДНК-топоизомеразы VI, унаследованной эукариотами от своих прокариотических предков – архей.
Белковые оси хромосом
В клетках эукариот, делящихся путем митоза, сестринские хроматиды (образовавшиеся в результате репликации исходной молекулы ДНК) соединены посредством белков-когезинов. Чтобы хроматиды могли разойтись к полюсам клетки (в анафазе митоза), когезины должны гидролизоваться и исчезнуть из межхроматидного пространства.
Синаптонемные комплексы. Ароморфоз.
Происхождение белков СК


Сравнительный анализ структуры белков СК показал, что они возникли независимо у предков современных эукариот, давших начало разным крупным таксонам (растений, грибов, животных).
Почему в первом делении мейоза сестринские хроматиды не расходятся?
С фактами и теориями, касающимися происхождения мейоза, можно ознакомиться в статьях Ю.Ф.Богданова и его коллег:
Читайте также: