
Методы определения вирусов в объектах окружающей среды
Обновлено: 15.04.2024
1. Дайте понятие вирусам. Охарактеризуйте особенности строения и жизни вирусной частицы.
2. Какими факторами осуществляется защита организма человека от вируса.
3. Назовите группу и механизм действия препаратов на вирусы. Приведите примеры препаратов.
4. Назовите типы инфекции, вызываемые вирусами.
5. Назовите представителей кишечных, кровяных, респираторных вирусных инфекций, инфекций кожных покровов и слизистых.
7. Назовите, как называются мероприятия, ликвидирующие эпидемический процесс.
Самостоятельная работа студентов:
Запишите определения методов исследования вирусных инфекций.
Зарисйте в атлас внутриклеточные включения при натуральной оспе (тельца Гварниери), при бешенстве (тельца Бабеша-Негри).
3. Составьте план противоэпидемических мероприятий на вирусную инфекцию (инфекцию определяет преподаватель).
Краткие теоретические положения
Введение
Расширение возможностей в лечении и профилактике вирусных болезней с использованием противовирусных препаратов, иммуномодуляторов и вакцин с различным механизмом действия нуждается в быстрой и точной лабораторной диагностике. Узкая специфичность некоторых противовирусных препаратов также требует быстрой и высокоспецифичной диагностики инфицирующего агента. Появилась необходимость в количественных методах определения вирусов для мониторинга противовирусной терапии. Помимо установления этиологии заболевания лабораторная диагностика имеет важное значение в организации противоэпидемических мероприятий.
Ранняя диагностика первых случаев эпидемических инфекций позволяет своевременно провести противоэпидемические мероприятия – карантин, госпитализацию, вакцинацию и пр. Реализация программ по ликвидации инфекционных заболеваний, например натуральной оспы, показала, что по мере их выполнения возрастает роль лабораторной диагностики. Существенную роль играет лабораторная диагностика в службе крови и акушерской практике, например, выявление доноров, инфицированных вирусом иммунодефицита человека (ВИЧ), вирусом гепатита В (HBV), диагностика краснухи и цитомегаловирусной инфекции у беременных.
Методы диагностики вирусных инфекций
Для успешного выделения вирусов клинический материал должен быть взят в соответствии с патогенезом предполагаемого заболевания и в наиболее ранние сроки.
Как правило, берутся:
– при респираторных инфекциях – носоглоточный смыв;
– при энтеровирусных инфекциях – смыв и фекалии (рео-, энтеровирусы);
– при поражениях кожи и слизистых оболочек – соскобы, содержимое пузырьков (герпес, ветряная оспа);
– при экзантемных инфекциях – смывы (корь, краснуха);
– при арбовирусных инфекциях – кровь, спинномозговая жидкость.
1. Быстрые (экспресс-методы) — прямое обнаружение вируса или его компонентов (антигенов, НК), включений непосредственно в клиническом материале.
А. Вирусоскопический метод заключается в обнаружении вируса в исследуемом материале под микроскопом. Чаще всего используют электронный микроскоп. Световая микроскопия из-за ничтожно малых размеров вирусов практически не применяется. При данном методе можно определить тип НК, размеры вириона, форму вириона, а также выявить внутриклеточные включения, которые образуются в пораженных клетках при некоторых инфекциях.


II. Вирусологический метод основан на:
Культивировании вирусов в чувствительных биологических системах (клеточных культурах, курином эмбрионе, организмах лабораторных животных),их индикации по цитопатогенному действию на биологическую систему (рис.1), идентификации по ингибиции действия вирусов соответствующими противовирусными антителами (рис.2).


Рис. 1. Цитопатическое действие вирусов на клетку: А - нормальный рост, Б - ЦПД вирусов на клетку

Рис.2 Ингибиция вируса антителами
Вирусологическое исследование - это "золотой стандарт" вирусологии и должно проводится в специализированной вирусологической лаборатории. В настоящее время оно используется практически только в условиях возникновения эпидемической вспышки того или иного вирусного инфекционного заболевания.
III. Серологический метод — определение противовирусных антител (оптимально — IgM) и/или определение динамики нарастания их титров за определенный период заболевания в парных сыворотках. Диагностически значимым считают нарастание титра антител в 4 и более раз.
Метод парных сывороток: осуществляем сбор венозной крови в количестве 10 мл в начале болезни и в конце, приготавливаем сыворотку, определяем количество антител в первой и второй сыворотке.
При этом четырехкратное нарастание титра антител во второй сыворотке в большинстве случаев служит показателем протекающей или свежеперенесенной инфекции. При исследовании одной сыворотки, взятой в острой стадии болезни, диагностическое значение имеет обнаружение антител класса Ig М, свидетельствующее об острой инфекции.
Современные методы диагностики:
1. ПЦР - выявляют персистирующие вирусы по НК, находящиеся в клиническом материале, с трудом обнаруживаемые или не обнаруживаемые другими методами.
2. Радиоизотопный иммунный анализ (РИА) - метод основан на метке антител радиоизотопами, что обеспечивало высокую чувствительность в определении вирусного антигена. Широкое распространение метод получил в 80-е годы, особенно для определения маркеров HBV и других некультивируемых вирусов. К недостаткам метода относится необходимость работать с радиоактивными веществами и использования дорогостоящего оборудования (гамма-счетчиков).
3. Иммуноферментный анализ (ИФА) – Иммуноферментные методы определения вирусных антигенов в принципе сходны с РИФ, но основываются на мечении антител ферментами, а не красителями. Наиболее широко используется пероксидаза хрена и щелочная фосфатаза, применяют также b-галактозидазу и b-лактамазы. Меченые антитела связываются с антигеном, и такой комплекс обнаруживается при добавлении субстрата для фермента, с которым конъюгированы антитела. Конечный продукт реакции может быть в виде нерастворимого осадка, и тогда учет проводится с помощью обычного светового микроскопа, или в виде растворимого продукта, который обычно окрашен (или может флюоресцировать или люминесцировать) и регистрируется инструментально.
Поскольку с помощью ИФА можно измерять растворимые антигены, то не требуется наличия интактных клеток в образце и таким образом могут использоваться различные виды клинического материала.
Другое важное преимущество метода ИФА – возможность количественного определения антигенов, что позволяет применять его для оценки клинического течения болезни и эффективности химиотерапии. ИФА, как и РИФ, может применяться как в прямом, так и в непрямом варианте.
Твердофазный ИФА, дающий растворимый окрашеный продукт реакции, нашел наибольшее распространение. ИФА может быть использован как для определения антигена (тогда на твердую фазу – дно лунки полистиролового планшета – наносятся антитела), так и для определения антител (тогда на твердую фазу наносятся антигены).
4. Реакция иммунофлюоресценции (РИФ) – Метод основан на использовании антител, связанных с красителем, например флюоресцеинизотиоцианатом. РИФ широко применяется для выявления вирусных антигенов в материале больных и для быстрой диагностики.
В практике применяются два варианта РИФ: прямой и непрямой. В первом случае применяются меченные красителем антитела к вирусам, которые наносятся на инфицированные клетки (мазок, культура клеток). Таким образом, реакция протекает одноэтапно. Неудобством метода является необходимость иметь большой набор конъюгированных специфических сывороток ко многим вирусам.
При непрямом варианте РИФ на исследуемый материал наносится специфическая сыворотка, антитела которой связываются с вирусным антигеном, находящимся в материале, а затем наслаивается антивидовая сыворотка к гамма-глобулинам животного, в котором готовилась специфическая иммунная сыворотка, например антикроличья, антилошадиная и т. п. Преимущество непрямого варианта РИФ состоит в потребности лишь одного вида меченых антител.
Метод РИФ широко применяется для быстрой расшифровки этиологии острых респираторных вирусных инфекций при анализе мазков-отпечатков со слизистой оболочки верхних дыхательных путей. Успешное применение РИФ для прямой детекции вируса в клиническом материале возможно лишь в случае содержания в нем достаточно большого числа инфицированных клеток и незначительной контаминации микроорганизмами, которые могут давать неспецифическое свечение.
5. Другие методы диагностики –
РТГА используется для диагностики заболеваний, вызванных гемагглютинирующими вирусами. Она основана на связывании антителами сыворотки больного добавленного стандартного вируса. Индикатором реакции являются эритроциты, агглютинирующиеся вирусом (формирование характерного "зонтика") при отсутствии специфических антител и оседающие на дно неагглютинированными при их наличии.
РСК является одной из традиционных серологических реакций и используется для диагностики многих вирусных инфекций. В реакции принимают участие две системы: антитела сыворотки больного + стандартный вирус и эритроциты барана + антитела к ним, а также оттитрованный комплемент. При соответствии антител и вируса этот комплекс связывает комплемент и лизиса бараньих эритроцитов не происходит (положительная реакция). При отрицательной РСК комплемент способствует лизису эритроцитов. Недостатком метода является его недостаточно высокая чувствительность и трудность стандартизации реагентов.
Для учета значимости РСК также, как и РТГА, необходимо титрование парных сывороток, то есть взятых в начале заболевания и в период реконвалесценции.
РПГА – агглютинация сенсибилизированных вирусными антигенами эритроцитов (или полистироловых шариков) в присутствии антител. На эритроцитах могут быть сорбированы любые вирусы, независимо от наличия или отсутствия у них гемагглютинирующей активности. В связи с наличием неспецифических реакций сыворотки исследуются в разведении 1:10 и более.
РНГА – агглютинация эритроцитов, сенсибилизированных специфическими антителами в присутствии вирусных антигенов. Наибольшее распространение РОПГА получила при выявлении HBs-антигена как у больных, так и у доноров крови.
Вирус – неклеточная форма жизни, обладающая геномом (РНК или ДНК), но лишенная собственного синтезирующего аппарата и, поэтому, способная к воспроизведению лишь в клетках более высокоорганизованных существ.
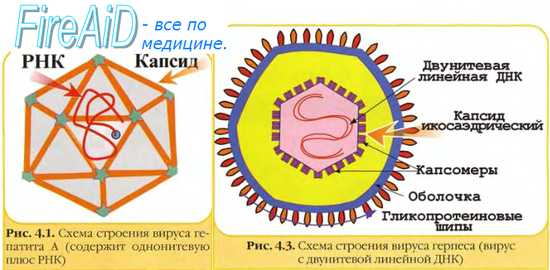
По химическому составу и потенциальной патогенности вирусы называют инфекционными нуклеопротеидами. Для вирусов характерны две формы существования: внеклеточная (покоящаяся) и внутриклеточная (репродуцирующаяся, вегетативная). Внеклеточная форма называется вирусной частицей или вирионом. Вирионы состоят из нуклеиновой кислоты, окруженной снаружи белковой оболочкой – капсидом (от лат. сapsa – футляр). Капсид вместе с заключенной в нем нуклеиновой кислотой называют нуклеокапсидом. Морфологическими субъединицами капсида, видимыми в электронный микроскоп, являются капсомеры – белковые субъединицы, состоящие из одной или нескольких молекул белка. Существуют три типа строения капсидов, основанных на расположении морфологических субъединиц (рис. 53):
1. вирионы со спиральной симметрией;
2. вирионы с кубической (икосаэдрической) симметрией;
3. вирионы, имеющие смешанный тип симметрии.
У первого типа капсомеры расположены в виде спирали, нуклеиновая кислота (преимущественно РНК) также скручена в виде пружины, располагаясь между витками белковых молекул. У вирусов с кубической симметрией капсомеры расположены в виде правильного икосаэдра со скрученной в клубок нитью ДНК или РНК. Икосаэдр имеет 20 граней (каждая представляет равносторонний треугольник), 12 вершин. Общее количество капсомеров (N) можно определить по формуле:
n – число капсомеров на одной стороне каждого равностороннего треугольника, которое у различных вирусов варьирует от 2 до 6. Так, аденовирус содержит 252, вирус герпеса – 162 капсомера.
К третьему типу относится вирус осповакцины. Вирус имеет внешнюю оболочку, состоящую из трех слоев, под оболочкой расположены два белковых тела, в центре вириона находится нуклеоид, в состав которого входит ДНК и внутренний белок.
Просто устроенные вирусы, такие как пикорна-, парвовирусы состоят из нуклеокапсида, сложноустроенные вирусы имеют еще дополнительную внешнюю оболочку – суперкапсид или пеплос (производное мембранных структур клетки-хозяина). Форма таких вирионов приближается к сферической. Суперкапсидные белки формируют морфологические субъединицы (пепломеры), которые в электронном микроскопе выглядят в виде шипов (тогавирус, коронавирус, ортомиксовирус и др.). Капсид и суперкапсид защищают вирионы от воздействий окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с определенными клетками, а также антигенные и иммуногенные свойства вирионов (рис. 53). Размеры вириона колеблются от 20–30 нм (пикорна-, парвовирусы) до 150–250 нм (герпес-, рабдовирусы) и даже 350–400 нм (поксвирусы).
Кроме обычных вирусов, известны и, так называемые, неканонические вирусы: прионы и вироиды. Прионы – это белковые инфекционные частицы, имеющие вид фибрилл размером 10-20х200 нм, они вызывают у животных и человека энцефалопатии в условиях медленной вирусной инфекции (болезнь Крейтцфельда –Якобы, куру и др.). Вироиды – это небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка и вызывающие заболевание растений.
Методы исследования вирусов. Для характеристики вирусных частиц широко применяют физические и физико-химические методы. Пользуясь ими, можно определить размер, форму, коэффициент седиментации, коэффициент диффузии, плотность и молекулярный вес как самой вирусной частицы, так и ее компонентов.
Рис. 53. Строение и основные типы симметрии вирусов. А – безоболочечный вирус с икасаэдрическим типом симметрии; Б – оболочечный вирус с икасаэдрическим типом симметрии; В – безоболочечный вирус со спиральным типом симметрии; Г – оболочечный вирус со спиральным типом симметрии. (Медицинская микробиология Под редакцией Покровского В.И. и Поздеева О.К., М., 1998.)
Для определения размеров вирусных частиц используют: фильтрование вируссодержащего материала через мембраны, ультрацентрифугирование, электрофорез, электронную микроскопию.
Фильтрование через коллодиевые мембраны. Метод основан на пропускании вируссодержащего материала через мембраны с известным размером пор. Размер вирусной частицы в данном случае определяется весьма приблизительно.
Осаждение при ультрацентрифугировании. Многие способы определения размеров вирионов основаны на анализе скорости их движения в суспендирующей жидкости. Частицы, взвешенные в жидкости, оседают с разной скоростью, благодаря чему компоненты взвеси можно быстро разделить центрифугированием. Скорость осаждения частицы прямо пропорциональна разности плотности частиц и жидкости, квадрату угловой скорости и квадрату радиуса окружности и обратно пропорциональна вязкости жидкости. Скорость осаждения зависит от формы оседающих частиц. Сферическая частица радиуса r, находящаяся в жидкости с плотностью d0, будет седиментировать в гравитационном поле или поле центробежных сил, если плотность частицы d больше, чем d0.
r – радиус частицы, v – скорость осаждения частицы, ŋ – вязкость среды, с – центробежное ускорение, d – плотность частицы, d0 – плотность жидкости. Это равенство вытекает из формулы Стокса и описывает движение сферических частиц в жидкости при идеальных условиях. Величина S=v/s называется константой седиментации и характеризует поведение данной частицы в данной среде при данной температуре. Константа седиментации выражается в единицах Сведберга, одна единица Сведберга соответствует скорости седиментации в воде при 20 0 С под действием единицы центробежной силы. Так как центробежная сила, плотность среды и ее вязкость могут быть измерены, то можно определить радиус и массу сферической частицы при условии, если мы сможем измерить ее плотность и скорость осаждения в центрифуге. Вирусы хорошо седиментируют в скоростных ультрацентрифугах (60000 об/мин и выше). Используя аналитические роторы, в которых луч видимого или ультрафиолетового света проходит через центрифужные ячейки с прозрачными стенками, можно проводить измерения при движении центрифуги. При седиментации однородной популяции светопоглощающих частиц (вирионов) образуется резкая подвижная граница, положение которой определяют либо непосредственно путем измерения поглощения света, либо по положению области, где показатель преломления жидкости резко изменяется.
Прямое исследование в электронном микроскопе. Электронная микроскопия – наиболее широко применяемый метод определения размеров вирусных частиц. Для этого увеличение на электронных микрофотографиях должно быть откалибровано с использованием внутреннего маркера. С этой целью можно использовать частицы вируса табачной мозаики, имеющие среднюю длину 300 нм и шаг спирали 2,3 нм; сывороточный альбумин – 5 нм; глобулин – 7 нм; гемоцианин – 23 нм. Метод исключительно быстр, прост и позволяет судить не только о размере вирионов, но отчасти об их форме и характере симметрии.
Методы изучения морфологии вирусных частиц. Детали структуры вируса можно различить только в электронном микроскопе (рис. 54). Широко используется метод негативного контрастирования. Он сводится к смешиванию суспензий вирусных частиц с раствором соли тяжелого металла, нанесению тонкого слоя полученной суспензии на сетку из вольфрамовой пленки и высушиванию полученного препарата. Соль образует плотный слой, на фоне которого материал выглядит сравнительно прозрачным. Обычно соль проникает в различные компоненты вирусной частицы неодинаково, благодаря чему возникает достаточный контраст, способствующий выявлению тончайших деталей структуры вирусной частицы.К числу соединений, наиболее широко применяемых для негативного контрастирования, относятся уксуснокислый и муравьинокислый уранил, кремневольфрамовокислый натрий и молибдат аммония, натриевая или калиевая соль фосфорновольфрамовой кислоты (ФВК).
Рис. 54. Вид аденовируса (А) при электронной микроскопии (ув. 600000 раз) и его модель (Б). (Авакян А.А., Быковский А.Ф., М .,Атлас анатомии и онтогенеза вирусов человека и животных, 1970).
Важные сведения о структуре и морфогенезе вирусов дает также метод тонких срезов, используемый для изучения препаратов вирусов, находящихся в осадке. Для того чтобы установить локализацию специфических белков в вирусной частице используют вспомогательные методы, например радиоавтографию, обработку тонких срезов мечеными антителами.
В ряде случаев используется действие на вирионы поверхностно-активных веществ. При этом оценивают формы, оставшиеся после обработки и делаются выводы о том, какие компоненты из структуры были удалены.
Для изучения вирусных нуклеиновых кислот используют методы гибридизации нуклеиновых кислот.
Изучение физико-химических свойств вирусов
Электрофорез. Электрофоретические методы позволяют определить специфическую физическую характеристику вирусной частицы – относительную электрофоретическую подвижность под влиянием электрического поля. В противоположность коэффициентам седиментации и диффузии эта величина практически не зависит от массы частицы и основывается главным образом на суммарном заряде поверхности частицы. Расчет константы электрофоретической подвижности производится по формуле:
где χ – удельная проводимость, в Ω -1 .см -1 ; q – поперечное сечение электрофоретической колонки, в см; s – путь, пройденный частицей; i – сила тока в А; t – время пробега в сек. Подвижность выражается в см 2 /вольт/сек.
Классический метод электрофореза, или метод движущейся границы Тизелиуса, широко используемый для анализа белков применяется для физико-химической характеристики вирусных частиц – определения изоэлектрической точки и электрофоретической подвижности.
Фронтальный электрофорез – единственный метод, позволяющий определить электрофоретическую подвижность и изоэлектрическую точку с очень высокой точностью. Кроме того, этот метод широко используется при исследовании гомогенности и степени чистоты вирусных препаратов.
Еще более разрешающий метод – метод зонального электрофореза в градиенте плотности и в гелях. Его используют не только для изучения специфической электрофоретической подвижности вирусов, их дифференцировки и идентификации, но и для выделения генетически однородных штаммов и изучении изменчивости микроорганизмов.
Методы фракционирования вирусов. Эти методы позволяют проводить дезинтегрирование вирусных частиц на отдельные компоненты и их фракционирование. Выделенные компоненты в дальнейшем можно подвергнуть биохимическому анализу с целью изучения их тонкой структуры и свойств. Фракционирование компонентов вирусной частицы осуществляется при помощи центрифугирования в зональном и равновесном градиентах плотности, зонального электрофореза и хроматографии.
Хроматография. Для очистки и фракционирования вирусов применяют три типа хроматографии: адсорбционную, ионообменную и молекулярно-ситовую.
Метод адсорбционной хроматографии основан на различной степени адсорбции компонентов смеси при фильтровании через неподвижный твердый адсорбент. Решающее значение имеют поверхностные свойства вирусной частицы и адсорбента, а также состав буфера, в котором суспендирован вирус. При элюировании соответствующим буфером вирусные частицы можно отделить от примесей. В качестве адсорбентов для наполнения хроматографических колонок в вирусологической практике чаще всего используют фосфат калия и гидроксилапатит.
При ионообменной хроматографии вирусы пропускают через ионообменник. Ионообменниками называют такие соединения, которые содержат фиксированные функциональные группы и подвижные противоионы. Последние могут обратимо обмениваться с другими ионами того же заряда, не изменяя физические свойства нерастворимой матрицы. Ионообменниками могут быть органические и неорганические соединения. В качестве матрицы могут быть использованы алюмосиликаты, синтетические смолы, полисахариды, белки и целлюлоза.
Лабораторные исследованияпри проведении идентификации вирусов и диагностике вирусных инфекций включают следующие этапы: выделение, культивирование, индикация (выявление) и идентификация вирусов.
2.3.1 Культивирование вирусов
Вирусы не растут на искусственных питательных средах, а размножаются только внутриклеточно. Крупным достижением было предложение Р. Гудпасчура в 1932 г. использовать для культивирования вирусов куриные эмбрионы. Окончательное решение проблемы культивирования вирусов оказалось возможным лишь после того, как были разработаны основные способы культивирования клеток вне организма.
Использование куриных эмбрионов. Куриные эмбрионы – практически идеальные модели для культивирования некоторых вирусов (например, гриппа и кори). Замкнутая полость эмбриона препятствует проникновению микроорганизмов извне, а также развитию спонтанных вирусных инфекций. Эмбрионы применяют для первичного выделения вирусов из патологического материала; для пассирования и сохранения их, а также для получения необходимых количеств вируса. Некоторые возбудители (например, герпесвирусы) вызывают характерные изменения (по ним можно распознавать заболевание).
Для заражения обычно используют куриные эмбрионы 7–12-дневного возраста. Перед заражением определяют жизнеспособность эмбриона путем овоскопирования (просматривают в проходящем свете). Живые эмбрионы при овоскопировании проявляют двигательную активность, хорошо виден сосудистый рисунок. Простым карандашом очерчивают границы воздушной камеры.
Куриные эмбрионы заражают вируссодержащим материалом в асептических условиях стерильными инструментами, предварительно обработав скорлупу над воздушным пространством йодом и спиртом. Заражение проводят на хорион-аллантоисную оболочку, в амниотическую или аллантоисную полость, либо в желточный мешок (рисунок 29). Выбор метода заражения зависит от биологических свойств вируса.
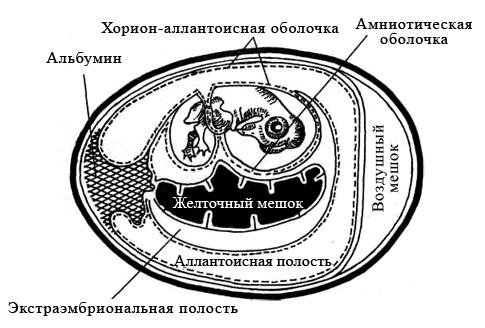
Рисунок 29 – Схематическое изображение развивающегося куриного эмбриона
Культура клеток. Вначале был использован метод переживающих тканей. Он заключался в том, что в колбу, содержащую питательную среду, вносили кусочек ткани. Клетки некоторых тканей в таких условиях могут переживать (но не размножаться) до 30 дней, а в них могут размножаться вирусы. Однако этот способ давал очень небольшой выход вирусов. Необходимо было разработать условия, при которых клетки ткани могли бы свободно размножаться.
Для получения культур клеток необходимо было решить четыре главных задачи:
– получить в необходимом количестве свободные (т. е. изолированные друг от друга) клетки;
– создать такие питательные среды и условия, в которых клетки могли бы активно размножаться;
– обеспечить условия, при которых в культурах клеток не могли бы размножаться бактерии;
– определить методы, с помощью которых можно было бы распознавать рост вируса в культуре клеток и идентифицировать его.
Для выделения изолированных (разобщенных), но жизнеспособных клеток из разрушенных тканей, стали использовать обработку их слабым раствором трипсина, разрушающего межклеточные мостики. Для культивирования клеток были предложены различные среды, содержащие все необходимые для размножения клеток питательные вещества (аминокислоты, основания, витамины и другие), минеральные соли, имеющие оптимальную рН и т. д. К питательным средам добавляли индикатор, по изменению цвета которого можно было судить о метаболизме клеток и их размножении. Было установлено, что в качестве основы, на которой клетки размножаются и образуют монослой, может быть использовано хорошо обработанное стекло пробирок и колб. Для подавления возможного роста бактерий вируссодержащий материал перед посевом его в культуры клеток стали обрабатывать антибиотиками.
В 1949 г. Дж. Эндерс, Т. Веллер и Ф. Роббинс показали, что вирус полиомиелита хорошо размножается в первично-трипсинизированных культурах клеток, полученных из почек обезьян. Основной недостаток первично-трипсинизированных клеток заключается в том, что после нескольких пересевов они перестают размножаться. Поэтому предпочтением стали пользоваться культуры таких клеток, которые способны размножаться in vitro бесконечно долго. Такие перевиваемые культуры клеток (клеточные линии характеризуются бессмертием и гетероплоидным кариотипом) получают из опухолевых тканей (HeLa получена из карциномы шейки матки, НЕр-2 – из карциномы гортани; Детройт-6 – из метастаза рака легкого в костный мозг; RН – из опухоли почки человека) или из мутантных клеток с полиплоидным набором хромосом. Однако опухолевые клетки нельзя применять для получения вакцин. Для этих целей используют только культуры таких клеток, которые не содержат никаких контаминантных вирусов и не обладают злокачественностью. Лучше всего этим требованиям отвечают культуры диплоидных клеток.
Полуперевиваемые (диплоидные) культуры клеток – клетки одного генотипа, способные in vitro выдерживать 50–100 пассажей, сохраняя при этом свой исходный диплоидный набор хромосом. Диплоидные линии фибробластов эмбриона человека используются как для диагностики вирусных инфекций, так и при производстве вирусных вакцин. Как оказалось, вирусы могут размножаться не только в культурах клеток, образующих монослой на стекле пробирок, но и в суспензиях живых клеток.
Для обеспечения жизнедеятельности культивируемых клеток необходимы питательные среды. По назначению они делятся на ростовые и поддерживающие. В ростовых питательных средах должно содержаться больше питательных веществ, обеспечивающих активное размножение клеток и формирование монослоя. Поддерживающие среды обеспечивают переживание клеток в уже сформированном монослое в период размножения в них вирусов.
2.3.2 Выделение вирусов
Выделение вирусов в культурах клеток. При выделении вирусов из различных инфекционных материалов (кровь, моча, слизистые отделяемые, смывы из органов) применяют культуры клеток, обладающих наибольшей чувствительностью к предполагаемому вирусу. Для заражения используют культуры в пробирках с хорошо развитым монослоем клеток. Перед заражением клеток питательную среду удаляют и в каждую пробирку вносят по 0,1–0,2 мл взвеси исследуемого материала, предварительно обработанного антибиотиками для уничтожения бактерий и грибов. После 30-60 мин контакта вируса с монослоем клеток удаляют избыток материала, в культуру вносят поддерживающую среду и пробы оставляют в термостате до выявления признаков размножения вируса.
Выделение вирусов на лабораторных животных. При невозможности выделить и идентифицировать вирус стандартными методами in vitro инфекционный материал вводят чувствительным к возбудителю животным, и после развития типичного инфекционного процесса проводят повторное заражение чувствительных клеточных культур. Наиболее часто используют мышей, кроликов и обезьян; для выделения некоторых вирусов (например, вирусов Коксаки) заражают мышат-сосунков. Вследствие дороговизны и сложности содержания лабораторных животных, практически повсеместно их вытеснили клеточные культуры. Тем не менее животные модели активно используют для изучения особенностей патогенеза и формирования иммунных реакций при вирусных инфекциях.
Таким образом, для выделения чистых культур вирусов в лабораторных условиях в настоящее время используются следующие живые объекты (биологические модели): 1) культура клеток (тканей, органов); 2) куриные эмбрионы; 3) лабораторные животные.
2.3.3 Индикация вирусов
Индикация вирусов в культурах клеток. Индикатором наличия вируса в зараженных культурах клеток может служить:
1) развитие специфической дегенерации клеток – цитопатическое действие вируса (ЦПД), имеющее три основных типа: крупно- или мелкоклеточная дегенерация; образование многоядерных гигантских клеток (симпластов); развитие очагов клеточной пролиферации, состоящих из нескольких слоев клеток (гроздевидная дегенерация клеток).
Различают два механизма гибели клеток, вызываемой вирусами, – некроз и апоптоз. Некроз происходит из-за необратимых нарушений целостности клеточных мембран, апоптоз – вследствие фрагментации ядерной ДНК под действием клеточной эндонуклеазы.
Цитопатические эффектыоценивают при микроскопии клеточных культур. По степени поражения клеток выделяют вирусы с высокой или умеренной цитопатогенностью:
2) обнаружение внутриклеточных включений, располагающихся в цитоплазме и/или в ядрах пораженных клеток;
3) положительная реакция гемагглютинации (РГА) или гемадсорбции (РГАдс). Некоторые вирусы, в частности, вирус гриппа, обладают особыми рецепторами (гемагглютининами), с помощью которых они адсорбируются на эритроцитах и вызывают их склеивание (гемагглютинацию). Такие вирусы легко обнаруживаются с помощью реакции гемагглютинации или гемадсорбции (эритроциты адсорбируются на инфицированных вирусами клетках культуры тканей);
4) феномен бляшкообразования. Широкое распространение получил предложенный в 1952 г. Р. Дюльбекко метод бляшек (негативных колоний), позволяющий производить количественное определение вирусов. Для выделения вирусов монослой клеток после удаления питательной среды заражают вируссодержащим материалом и покрывают слоем агара, содержащего индикатор нейтральный красный. Чашки (флаконы) инкубируют при 37 °С. Через 48–96 ч выявляются пятна – бляшки. Они имеют диаметр 1–3 мм и выглядят неокрашенными на розовом фоне. Пятна возникают за счет цитопатического действия вируса;
5) цветная реакция Солка. О росте вирусов в клетках можно судить с помощью индикатора, добавляемого к питательной среде. Если клетки активно осуществляют метаболизм, рН среды сдвигается в кислую сторону, и среда окрашивается в желтый цвет. В случае размножения вируса клетки погибают, рН среды мало меняется, и она сохраняет первоначальный (малиновый) цвет или (при нейтральной рН) приобретает оранжевый;
6) реакция интерференции (используется при отсутствии ЦПД, гемагглютинации и гемадсорбции): исследуемая культура повторно заражается вирусом, вызывающим ЦПД. В положительном случае ЦПД будет отсутствовать (реакция интерференции положительна). Если в исследуемом материале вируса не было, наблюдается ЦПД.
Кроме того, для обнаружения вируса в культурах клеток могут быть использованы различные серологические реакции.
Индикация вирусов на лабораторных животных. Индикация вируса основана на обнаружении у животных признаков инфекционного заболевания, регистрации их гибели, изучении характера патоморфологических и патогистологических изменений в тканях и органах, выявлении положительной реакции гемагглютинации.
2.3.4 Методы идентификации вирусов
Определение типа вируса (его идентификация) основано на нейтрализации биологической активности вируса с помощью типоспецифических сывороток. Конечный результат ее может быть установлен на основании следующих признаков:
1) нейтрализация цитопатического действия: в культуральную среду, содержащую изучаемый вирус, вносят коммерческую сыворотку (например, к вирусу краснухи при подозрении на неё), инкубируют и заражают вторую культуру; через 1–2 дня в неё вносят известный цитопатогенный вирус. При наличии цитопатогенного эффекта делают вывод о том, что первая культура была заражена вирусом, соответствовавшим антителам примененной сыворотки;
2) нейтрализация реакции гемадсорбции;
3) изменение проявления цветной пробы;
4) задержка (торможение) реакции гемагглютинации: смешивают культуральную среду, содержащую возбудитель, с известной коммерческой антисывороткой и вносят в культуру клеток. После инкубации определяют способность культуры к гемагглютинации и при её отсутствии делают заключение о несоответствии вируса антисыворотке.
5) нейтрализация в опытах на животных.
Таким образом РН (реакция нейтрализации) основана на подавлении соответствующей реакции, феномена, развития инфекционного процесса после внесения в культуру или введения в организм животного смеси вируса со специфичными AT, содержащимися в диагностической сыворотке.
Вопросы для самоконтроля
1 Назовите основные принципы классификации вирусов.
2 Приведите русские и латинские названия основных семейств вирусов человека и животных.
3 Назовите типовых представителей основных семейств вирусов и заболевания, вызываемые ими.
4 Каковы особенности морфологии и ультраструктуры вирусов человека и животных (основных семейств)?
5 Назовите РНК-геномные и ДНК-геномные фитовирусы.
6 Какие этапы включают в себя лабораторные исследования при идентификации вирусов и диагностике вирусных инфекций?
7 Какие биологические модели используются для выделения и культивирования вирусов человека и животных?
8 Как происходит заражение куриных эмбрионов в лабораторных условиях?
9 Какие методы получения культуры клеток вы знаете?
10 Как проводят идентификацию вирусов в курином эмбрионе и на лабораторных животных?
11 Какие существуют методы индикации вирусов на культуре клеток?
12 В чем заключается назначение и сущность реакций нейтрализации вирусов?
Лабораторные методы при диагностике вирусных инфекций включают:
• выделение и идентификацию возбудителя;
• обнаружение и определение титров противовирусных AT;
• обнаружение Аг вирусов в образцах исследуемого материала;
• микроскопическое исследование препаратов исследуемого материала.
Забор материала для выявления вирусов
При заборе материала для исследований необходимо выполнять следующие условия:
• образцы следует отбирать как можно раньше либо с учётом ритма циркуляции возбудителя;
• материал следует отбирать в объёме, достаточном для всего комплекса исследований;
• образцы следует доставлять в лабораторию незамедлительно (!), при относительно кратковременной транспортировке (не более 5 сут) образцы сохраняют на льду, при более длительной — при температуре -50 С.
Выделение и культивирование вирусов
Выделение и идентификация возбудителя — золотой стандарт в диагностике вирусных инфекций.
Культуры клеток для выявления вирусов
Вирусы размножаются только в живых клетках, и выделение возбудителя в заражённой культуре клеток — один из основных методов диагностики вирусных инфекций. Поскольку большинство патогенных вирусов отличает тканевая и типовая специфичность, то почти к каждому вирусу можно подобрать соответствующие клеточные или тканевые чультуры, а также создать стандартные условия культивирования (наличие клеток одного типа). Размножение вируса обеспечивают чувствительные (пермиссивные) клетки. Поэтому при выделении неизвестного возбудителя проводят одномоментное заражение 3~4 культур клеток, предполагая, что одна из них может оказаться пермиссивной. Культуры клеток получают диспергированием соответствующих органов и тканей, но чаще используют эмбриональные ткани (человека и животных) либо трансформированные опухолевые клетки. При помещении на соответствующую плоскую поверхность клеточные культуры обычно растут в виде монослоя. Первично-трипсинизированные культуры. Суспензии клеток получают гомогенизированием соответствующих тканей, предварительно обработанных трипсином. Культуры часто представлены клетками смешанного типа и не подлежат повторному культивированию. Жизнеспособность таких культур составляет 2-3 нед.
Полуперевиваемые линии клеток представлены диплоидными клетками человека и животных. Культуры ограниченно пригодны к повторному диспергированию и росту (как правило, не более 20-30 пересевов), сохраняя при этом жизнеспособность и не подвергаясь спонтанной трансформации.
Перевиваемые линии клеток (гетероплоидные культуры) представлены клетками, подвергнутыми длительному культивированию и спонтанным трансформациям. Культуры способны к многократному диспергированию и перевиванию. Работа с ними менее трудоёмка по сравнению с приготовлениями первичных культур; перевиваемые клетки относительно одинаковы по своей морфологии и стабильны по свойствам.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Животные модели для обнаружения вирусов. Идентификация вирусов. Качественное определение вирусов. Цитопатические эффекты вирусов. Бляшкообразование вируса. Тельца включений вирусов.
При невозможности выделить и идентифицировать вирус стандартными методами in vitro инфекционный материал вводят чувствительным к возбудителю животным, и после развития типичного инфекционного процесса проводят повторное заражение чувствительных клеточных культур. Наиболее часто используют мышей, кроликов и обезьян; для выделения некоторых вирусов (например, вирусов Коксаки) заражают мышат-сосунков. Вследствие дороговизны и сложности содержания лабораторных животных, практически повсеместно их вытеснили клеточные культуры. Тем не менее животные модели активно используют для изучения особенностей патогенеза и формирования иммунных реакций при вирусных инфекциях. j
Идентификация вирусов
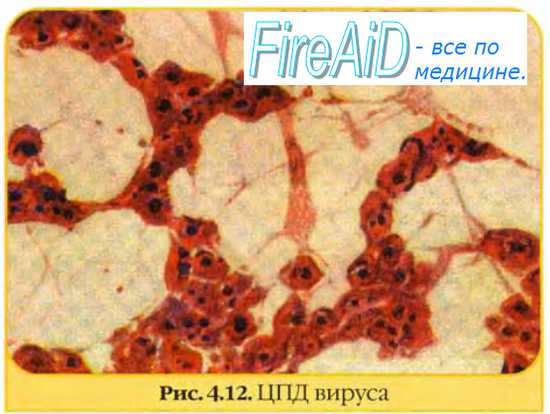
Цитопатические эффекты вирусов оценивают при микроскопии клеточных культур. По степени поражения клеток выделяют вирусы с высокой или умеренной цитопатогенностью. Размножение вирусов в культурах клеток сопровождается нарушениями морфологии клеток монослоя. Некоторые вирусы вызывают характерные цитопатические изменения, что (с учётом клинической картины заболевания) позволяет быстро поставить предварительный диагноз. Например, размножение парамиксовирусов (вирусы кори, паротита, PC-вирус) сопровождается появлением характерных гигантских многоядерных клеток; аденовирусы вызывают образование скоплений больших круглых клеток, а при репродукции герпесвирусов клетки округлой формы диффузно располагаются по всему монослою.
Бляшкообразование вирусов
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: