
Модификация у бактерий и вирусов
Обновлено: 24.04.2024
Различают два вида изменчивости – фенотипическую и генотипическую.
Фенотипическая изменчивость – модификации – не затрагивает генотип. Модификации затрагивают большинство особей в популяции. Они не передаются по наследству и с течением времени затухают, т. е. возвращаются к исходному фенотипу.
Генотипическая изменчивость затрагивает генотип. В основе ее лежат мутации и рекомбинации.
Мутации – изменение генотипа, сохраняющееся в ряду поколений и сопровождающееся изменением фенотипа. Особенностями мутаций у бактерий является относительная легкость их выявления.
По локализации различают мутации:
1) генные (точечные);
По происхождению мутации могут быть:
1) спонтанными (мутаген неизвестен);
2) индуцированными (мутаген неизвестен).
Рекомбинации – это обмен генетическим материалом между двумя особями с появлением рекомбинантных особей с измененным генотипом.
У бактерий существует несколько механизмов рекомбинации:
2) слияние протопластов;
Конъюгация – обмен генетической информацией при непосредственном контакте донора и реципиента. Наиболее высокая частота передачи у плазмид, при этом плазмиды могут иметь разных хозяев. После образования между донором и реципиентом конъюгационного мостика одна нить ДНК-донора поступает по нему в клетку-реципиент. Чем дольше этот контакт, тем большая часть донорской ДНК может быть передана реципиенту.
Слияние протопластов – механизм обмена генетической информацией при непосредственном контакте участков цитоплазматической мембраны у бактерий, лишенных клеточной стенки.
Трансформация – передача генетической информации в виде изолированных фрагментов ДНК при нахождении реципиентной клетки в среде, содержащей ДНК-донора. Для трансдукции необходимо особое физиологическое состояние клетки-реципиента – компетентность. Это состояние присуще активно делящимся клеткам, в которых идут процессы репликации собственных нуклеиновых кислот. В таких клетках действует фактор компетенции – это белок, который вызывает повышение проницаемости клеточной стенки и цитоплазматической мембраны, поэтому фрагмент ДНК может проникать в такую клетку.
Трансдукция – это передача генетической информации между бактериальными клетками с помощью умеренных трансдуцирующих фагов. Трансдуцирующие фаги могут переносить один ген или более.
1) специфической (переносится всегда один и тот же ген, трансдуцирующий фаг всегда располагается в одном и том же месте);
2) неспецифической (передаются разные гены, локализация трансдуцирующего фага непостоянна).
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Географическое распространение и изменчивость
Географическое распространение и изменчивость Большинству известно, что колибри водятся только в Америке; менее известно, что они – почти исключительно тропические птицы и что те немногие виды, которые попадаются в умеренных (северных и южных) широтах материка,
7. Виды метаболизма бактерий
7. Виды метаболизма бактерий В процессе метаболизма выделяют два вида обмена:1) пластический (конструктивный):а) анаболизм (с затратами энергии);б) катаболизм (с выделением энергии);2) энергетический обмен (протекает в дыхательных
3. Дополнительные органеллы бактерий
3. Дополнительные органеллы бактерий Ворсинки (пили, фимбрии) – это тонкие белковые выросты на поверхности клеточной стенки. Функционально они различны. Различают комон-пили и секс-пили. Комон-пили отвечают за адгезию бактерий на поверхности клеток макроорганизма. Они
ЛЕКЦИЯ № 3. Физиология бактерий
ЛЕКЦИЯ № 3. Физиология бактерий 1. Рост и размножение бактерий Рост бактерий – увеличение бактериальной клетки в размерах без увеличения числа особей в популяции.Размножение бактерий – процесс, обеспечивающий увеличение числа особей в популяции. Бактерии
1. Рост и размножение бактерий
1. Рост и размножение бактерий Рост бактерий – увеличение бактериальной клетки в размерах без увеличения числа особей в популяции.Размножение бактерий – процесс, обеспечивающий увеличение числа особей в популяции. Бактерии характеризуются высокой скоростью
2. Питание бактерий
2. Питание бактерий Под питанием понимают процессы поступления и выведения питательных веществ в клетку и из клетки. Питание в первую очередь обеспечивает размножение и метаболизм клетки.Среди необходимых питательных веществ выделяют органогены – это восемь
Изменчивость.
Изменчивость. Прежде чем применить выработанные в предыдущей главе общие основания к органическим существам в природе, мы должны вкратце обсудить, подвержены ли последние какой-либо вариации. Для надлежащего изложения этой темы потребовалось бы привести длинный
Рояль в кустах, или Скрытая изменчивость
Рояль в кустах, или Скрытая изменчивость Одно из фундаментальных свойств живых существ — помехоустойчивость. Многие случайные помехи (как внутренние — мутации, так и внешние — колебания условий среды) тем или иным способом компенсируются и не приводят к изменению
Глава 5. Изменчивость
Глава 5. Изменчивость Храбреца не разыгрывай перед судьбой, Каждый миг она может покончить с тобой. Твой доверчивый рот, услаждая халвою, Что ей стоит подсыпать отравы любой? Омар Хайям (1048–1123), персидский философ и поэт Всем живым организмам свойственна изменчивость,
Изменчивость
Изменчивость Всем живым организмам свойственна изменчивость, под которой понимают свойство приобретать новые признаки. В природе встречаются различные виды изменчивости.Модификационная изменчивость – это изменения фенотипа под действием факторов внешней среды в
Генетическая изменчивость мужских особей
Тема 5. Изменчивость
Тема 5. Изменчивость Победа какого-нибудь научного взгляда и включение его в мировоззрение не доказывает еще его истинности… Истина нередко в большем объеме открыта научным еретикам, чем ортодоксальным представителям научной мысли. В. И. Вернадский (1863–1945), русский
30. Изменчивость: наследственная и ненаследственная
30. Изменчивость: наследственная и ненаследственная Вспомните!Какие виды изменчивости вам известны?Приведите примеры признаков, изменяющихся под воздействием внешней среды.Что такое мутации?Изменчивость – одно из важнейших свойств живого, способность живых
6. Наследственность и ее изменчивость
6. Наследственность и ее изменчивость Наследственность — это одно из важнейших свойств организмов, которое не присуще телам неживой природы.Под наследственностью часто понимают способность передачи родителями своих свойств потомству.Но это очень узкое и ограниченное
Модификация – фенотипическая изменчивость касающаяся количественных проявлений признаков (уровень, степень выраженности), не затрагивающая генотип.
Мутация – изменчивость затрагивающая генотип. Мутации бывают адекватные и летальные. По направленности индуцибельные (под воздействием мутагена), и спонтанные (природный мутагенез). По локализации мутации бывают точечные (мутация нуклеотидов внутри одного гена), и генные (т.е. целыми генами).
И модификации и мутации создают широкий диапазон изменчивости внутри популяции, что делает её более жизнеспособной и способной к эволюции. Если количество мутаций превышает естественный уровень популяция может погибнуть.
Популяционный анализ: метод определения соотношений количества мутаций в популяции под воздействием определённых факторов внешней среды к фоновому уровню мутаций. Цель: определить мутагенные действия и уровень воздействия факторов внешней среды. Объект: популяция микроорганизмов с устойчивым количеством спонтанных мутаций.
Раздел 2 Инфекция и иммунитет.
1. Понятия: инфекционный процесс (движущие силы), инфекционная болезнь (условия возникновения). Внутрибольничные инфекции.
Инфекционный процесс – взаимодействие восприимчивого макроорганизма с патогенным микроорганизмом в определённых условиях внешней среды.
Движущие силы: -восприимчивый макроорганизм, -патогенный микроорганизм, -внешняя среда.
Формы инфекционного процесса:
1. Болезнь – крайнее выражение инфекционного процесса, протекающее с нарушением гомеостаза и клиническими проявлениями (бывает бессимптомная инфекционная болезнь).
2. Бактерионосительство – носительство патогенных микроорганизмов без нарушения гомеостаза.
3. Персистенция – длительное нахождение микроорганизма в макроорганизме.
Для возникновения инфекционной болезни необходимы условия:
Высокая вирулентность возбудителя.
Ослабление защитных сил организма.
Инфицирующая доза возбудителя.
Условия внешней среды, неблагоприятные для макроорганизма.
Внутрибольничные инфекции – инфекции, возникающие в стационаре при заражении больных от мед. персонала и наоборот.
Особенности внутрибольничных инфекций:
Восприимчивыми организмами являются больные, ослабленные люди.
Возбудителями являются патогенные и условно патогенные микроорганизмы (внутрибольничные штаммы, характеризуются устойчивостью к антибиотикам, УФЛ, бактериофагам, хлору, и т. д.). Эти штаммы может носить мед. персонал. Возбудители передаются через мед. инструментарий, перевязочный, шовный материалы.
2. Патогенность и вирулентность микробов. Определение. Факторы патогенности. Антилизоцимная активность бактерий и другие факторы персистенции.
Патогенные микроорганизмы характеризуются двумя свойствами:
1. Патогенность – видовое, генотипическое, неизменённое свойство – способность вступать в инфекционный процесс с восприимчивым макроорганизмом. Материальная основа патогенности – факторы патогенности, которые обуславливают свойства микроорганизма: инвазивность, токсичность, инфективность.
Токсины делятся на эндо- и экзотоксины. Эндотоксин – липополисахарид (О-антиген) – выделяется только при гибели клетки. Экзотоксин – специфический белок, продуцируемый живой клеткой. Адгезины – пили 1-го порядка, обеспечивающие прикрепление к субстрату. Ферменты защиты (плазмокоагулаза), и агрессии (гиалуронидаза, нейраминидаза). Факторы персистенции: антилизоцимная активность (АЛА), антикомплементарная активность (АКА), антиинтерфероновая активность (АИА) – секрктируемые факторы, направленные на инактивацию естественной защиты организма. Капсула – защита и антифагоцитарная активность. Антибиотикорезистентность. Поверхностные антигены, связывают антитела макроорганизма и препятствуют иммунитету. В зависимости от выраженности факторов патогенности микроорганизмы бывают более или менее вирулентными.
2. Вирулентность – индивидуальное, фенотипическое, вариабельное свойство – меры патогенности микроорганизмов. Вирулентность можно усилить последовательными пассажами через восприимчивые макроорганизмы (используется для изготовления атенуированных вакцин). Меры вирулентности: DLM (Dosis Letalis Minima) – минимальное количество микробов, способное вызвать гибель заражённых опытных животных. DL (Dosis Letalis) – количество микробов, вызывающее гибель 100% животных. DL50 наименьшая доза, убивающая 50% заражённых опытных животных. АЛА – способность инактивации лизоцима в секретах и сыворотке, способствует внутриклеточному паразитированию бактерий, т. к. стабилизирует ферменты лизосом. Не может выживать в условиях лизоцим продуцирующих конкурентов.
АКА – снижение литической активности комплемента.
АИА – инактивирование интерферона.
AIgA – антииммуноглобулиновая активность (блокирует антитела макроорганизма).
Различают генотипическую (наследуемую) изменчивость и фенотипическую (ненаследуемую, модификационную). Наследуемая изменчивость осуществляется в виде мутаций и рекомбинаций.
Мутации - это перегруппировка генов, не связанная с внесением в клетку нового генетического материала, ненаправленное изменение генотипа. Спонтанные мутации возникают без видимых причин, как ошибки репликации; индуцированные - под влиянием мутагенов (УФО, ионизирующей радиации, алкилирующих агентов, азотистой кислоты, аналогов оснований ДНК и др.). Мутации могут проявляться в виде удвоения, выпадения, замены нуклеотидов, вставки (в том числе транспозируемого элемента) и др.
Рекомбинации - это изменения генотипа, связанные с внесением в клетку-реципиент генетического материала от клетки-донора. Различают 3 вида рекомбинаций: трансформацию, трансдукцию, конъюгацию.
Трансформацией называют процесс: поглощения клеткой-реципиентом
изолированной ДНК клетки-донора (или синтетической нуклеиновой кислоты).
Трансдукция - перенос генетической информации (ДНК) при проникновении в клетку умеренного бактериофага (вирусных частиц, паразитирующих на бактериальных клетках).
Конъюгация - перенос генетической информации посредством трансмиссивных плазмид при непосредственном контакте донора и реципиента.
Мутации и рекомендации обеспечивают высокий уровень генетического обмена в различных микробиоценозах, позволяют микробам быстро адаптироваться к меняющимся условиям среды, эволюционировать, нередко приобретая нежелательные свойства - лекарственную устойчивость, повышенный патогенный потенциал. Все формы наследуемой изменчивости активно используются в генно-инженерных исследованиях и биотехнологии.
58. Виды изменчивости микробов. Модификации, виды, примеры
Различают генотипическую (наследуемую) изменчивость и фенотипическую (ненаследуемую, модификационную). Наследуемая изменчивость осуществляется в виде мутаций и рекомбинаций.
В условиях культивирования микробов возможен переход от S-формы к R-форме. При этом кап-сульные бактерии теряют капсулы, лишаются биохимической активности и становятся неполноценными в антигенном отношении, приобретая неспецифические антигены. Подвижные бактерии теряют жгутики.
Переход S-формы в R-форму происходит в основном через промежуточные О и слизистые М-колоиии. В процессе расщепления культур еще наблюдаются карликовые (D-dwarf), G-колонии (gonidial), появляющиеся как дочерние колонии на поверхности или на краю нормально развивающихся.
В условиях нарушения температурного режима, старения культуры, повышенной концентрации солей, применения антибиотиков и'фагов понижается вирулентность, изменяются антигенные и им-муногенные свойства, появляются антибиотикоустойчивые и фаго-устойчивые штаммы, аэробы становятся факультативными анаэробами, утрачивают некоторые имеющиеся ферменты или приобретают новые ферменты. Если культивировать кишечные палочки па среде с добавлением лактозы, то у нее появляется новый, фермепт р-галактоза. Изменение метаболизма у бактерий можно вызвать ультрафиолетовым облучением и рентгеновскими лучами. Изменяя параметры среды обитания, можно установить пределы и границы отклонения микробных клеток. Таким образом, широк диапазон феиотипических изменений. В основе этих изменений лежит прежде всего приспособительная активность обменных функций. Адаптация―это закон живого, и по этому закону живут и развиваются микроорганизмы.

Вирусология занимает важное место среди биологических дисциплин. Современный медицинский или ветеринарный специалист должен знать не только клинико–патологическую сторону заболевания, но и иметь четкое представление о вирусах, их свойствах, методах лабораторной диагностики и свойствах постинфекционного и поствакцинального иммунитета.

Вирус (от лат. virus — яд) является простейшей неклеточной формой жизни в виде микроскопической биологической частицы, представляющей собой молекулы нуклеиновых кислот (ДНК или РНК), заключённых в защитную белковую оболочку (капсид) и способные инфицировать живые организма.
- мутации, то есть изменении последовательности нуклеотидов в определенной области генома вируса, что приводит к фенотипически выраженному изменению свойства;
- рекомбинации, то есть обменом генетическим материалом между двумя вирусами, близкими, но различными по наследственным свойствам.
Мутации у вирусов
- спонтанные;
- индуцированные (вызванные).
Но точечные мутации не всегда приводят к изменению фенотипа. Существует целый ряд причин, по которым такие мутации не могут проявляться. Одна из них - вырождение генетического кода. Код синтеза белка вырождается, что означает, что некоторые аминокислоты могут быть закодированы несколькими триплетами (кодонами). Например, аминокислота лейцин может быть закодирована шестью триплетами. Поэтому, если молекула РНК заменяет триплет ЦУУ на ЦУЦ, ЦУА на ЦУГ, то синтезированная молекула белка все еще будет содержать аминокислоту лейцин.
Поэтому ни структура белка, ни его биологические свойства не нарушаются. Природа использует своего рода синонимичный язык и, заменяя один кодон другим, закладывает в них одно и то же понятие (аминокислоту), тем самым сохраняя естественную структуру и функцию синтезируемого белка.

Другое дело, если аминокислота кодируется только одним триплетом, например, синтез триптофана кодируется и заменяется только триплетом УГГ, то есть синонимом, который отсутствует. В этом случае в белок включается еще одна какаялибо аминокислота, которая может привести к появлению мутантного признака.
Аберрация в фагах вызвана делециями (потерями) различного числа нуклеотидов, от одной пары до последовательности, вызывающей одну или несколько функций вируса. Как спонтанные, так и индуцированные мутации также делятся на прямые и обратные мутации. Мутации могут иметь разные последствия. В некоторых случаях они приводят к изменению фенотипических проявлений в нормальных условиях.

Например, увеличивается или уменьшается размер бляшек под агарным покрытием; увеличивается или ослабевает вирулентность для определенного вида животных; вирус становится более чувствительным к действию химиотерапевтического агента и т. д.
В других случаях мутация является фатальной, поскольку она нарушает синтез или функцию жизненно важного вирусного белка, например, такого как вирусная полимераза. В некоторых случаях мутации являются условно летальными, так как вирусспецифический белок сохраняет свои функции при определенных условиях и теряет эту способность в неразрешающих (непермиссивных) условиях.
Типичным примером таких мутаций являются термочувствительные – ТS-мутации, при которых вирус теряет способность к размножению при повышенных температурах (+39-42°С), сохраняя эту способность при нормальных температурах роста (+36-37°С). Морфологические или структурные мутации могут влиять на размер вириона, первичную структуру вирусных белков и изменения в генах, определяющих ранние и поздние вирусные ферменты, обеспечивающие размножение вируса. Мутации также могут быть различными по своему механизму.
В одних случаях происходит делеция, то есть потеря одного или нескольких нуклеотидов, в других - встраивание одного или нескольких нуклеотидов, а в некоторых случаях один нуклеотид заменяется другим. Мутации могут быть прямыми или обратными. Прямые мутации меняют фенотип, а обратные мутации – реверсии) - восстанавливаются. Реальная реверсия возможна, когда обратная мутация происходит вместе с первичным повреждением, и псевдореверсия, когда мутация происходит в другой области дефектного гена (интрагенное торможение мутации) или в другом гене (экстрагенное подавление мутации).
Реверсия - не редкое явление, потому что ревертанты обычно лучше приспособлены к данной клеточной системе. Поэтому при создании мутантов с определенными свой ствами, например, вакцинных штаммов, следует ожидать возможного превращения их в дикий тип. Вирусы отличаются не только своими небольшими размерами, селективной способностью к размножению в живых клетках, особенностями строения наследственного вещества, но и значительной изменчивостью от других представителей живого мира.
Изменения могут влиять на размер, форму, патогенность, антигенную структуру, тканевую тропность, устойчивость к физико-химическим воздействиям и на другие свойства вирусов. Значение причин, механизмов и характера изменений имеет большое значение при получении необходимых вакцин для вирусных штаммов, а также для разработки эффективных мер борьбы с вирусными эпизодами, в ходе которых, как известно, свойства вирусов могут существенно изменяться.
Мутация вирусов может происходить в результате химических изменений цистронов или нарушения последовательности их расположения в структуре молекулы вирусной нуклеиновой кислоты. В зависимости от условий различают естественную изменчивость вирусов, наблюдаемую в нормальных условиях размножения, и искусственную изменчивость, получаемую в результате многочисленных специальных пассажей или воздействия на вирусы определенных физических или химических факторов (мутагенов). В обычных природных условиях изменчивость проявляется не во всех вирусах одинаково.
Этот признак наиболее заметен у вируса гриппа и вирус ящера. Значительная изменчивость отмечается у вируса гриппа. Об этом свидетельствует большое количество вариантов у разных типов этих вирусов, а также значительные изменения его антигенных свойств в конце почти каждой эпизоотии.
Частота мутаций и механизмы их возникновения
Мутации бактериофагов изучались очень интенсивно не только с целью генетического анализа, но и с целью получения информации о свойствах самих фагов. Частота появления мутантов в потомстве фагов варьируется очень сильно: например, одни мутанты образуются с частотой не более 10, а другие-с частотой 10 и выше. Неблагоприятное воздействие высокочастотных мутаций обычно компенсируется эффектом отбора. Например, мутантный фаг может быть заменен диким типом, что дает более высокий выход фага. Высокая частота вспышек обычно характерна для таких мутаций, которые могут происходить как во многих локусах, так в одном и том же локусе.
В тех случаях, когда нормальный признак соответствует функциональной форме гена, а мутант появляется в результате изменения в любой точке локуса, частота прямых мутаций окажется выше, чем частота обратных мутаций, так как обратные мутации должны приводить к восстановлению нормального состояния. Иногда ревертанты на самом деле являются псевдоревертантами: это происходит либо из-за изменений в другом гене (мутации-супрессоры), либо из-за изменений в том же гене, которые вызывают другую, но также активную форму продукта.
У зрелых фагов частота спонтанных мутаций очень мала, но они могут быть индуцированы под влиянием таких мутагенных факторов, как рентгеновские или ультрафиолетовые лучи, азотистая кислота, гидроксиламин или алкилирующие агенты. Азотистая кислота дезаминирует основания нуклеотидов, а этилметилсульфат их этилирует. Гидроксиламин превращает шитозин в урацил. В результате ошибок, допущенных при репликации химически модифицированной нуклеиновой кислоты, происходят мутации, и потомство фагов, полученное из бактерии, содержит как нормальные, так и мутантные частицы. Однако, как и при обработке мутагенного фага, содержащего одноцепочную ДНК, образуется чистый мутантный клон.
Изучение мутационного процесса, происходящего при размножении фагов, непосредственно связано с анализом развития фагов. Давайте рассмотрим процесс спонтанной мутации. В бактериальной клетке, в которой произошла мутация фага, 6 образуются как нормальный, так и мутировавший фаги. Количество мутантных фаговых частиц, содержащихся в популяции фагов, происходящих из этой отдельной бактериальной клетки, очевидно, определяется характером размножения фагов, поскольку новые гены могут быть сформированы только путем репликации уже существующих. Если вероятность мутации одинакова для каждой репликации, то число мутантов зависит от механизма репликации.
Например, если каждая новая копия гена формируется независимо от других, то распределение мутантных копий в потомках фагов от разных инфицированных бактерий будет случайным. Если же, наоборот, каждая из полученных копий воспроизводится, то в свою очередь мутантные копии будут разделены на группы или клоны, состоящие из мутантных "сибсов".
Индуцированные хозяином модификации бактериофагов
Помимо мутаций, бактериофаги подвержены негенетическим изменениям, в которых главная роль принадлежит клетке-хозяину. Это явление было названо модификациями, вызванными хозяином. Значение этих модификаций для молекулярной биологии состоит в том, что они показали способность внутриклеточной среды вызывать такие изменения в химической структуре генетического материала, которые могут быть использованы для идентификации клеточных линий, синтезирующих ДНК.
Подобные явления были впервые обнаружены на фаговой ДНК, но они также справедливы и для каждой бактериальной клеточной ДНК. Есть также наблюдения, при которых это явление относится и к эукариотическим клеткам. В особых случаях могут возникнуть более сложные ситуации. Двустороннее ограничение фага двумя хозяевами иногда наблюдается, но оно не обязательно. Фаги, отторгнутые клетками, способны адсорбироваться на них и проникать в их ДНК добавляя часть собственной ДНК. Однако последняя часть быстро разрушается, и репликация не происходит.
Деградация ДНК вызывается специфическими эндонуклеазами (рестриктазами или R-нуклеазами), которые могут обнаруживать и расщеплять определенные участки ДНК, если они не были модифицированы под влиянием М-ферментов. После этого ДНК расщепляется экзонуклеазами на отдельные нуклеотиды. Бактериальный штамм может иметь одну или несколько R-нуклеаз и в то же время M-ферменты, которые защищают собственную ДНК клетки. Предложена удобная номенклатура этих ферментов. Согласно ряду данных, области детекции R-нуклеазы не всегда совпадают с областями расщепления ДНК; возможно, что фермент может мигрировать по цепочке до того, как найдет область, где происходит расщепление ДНК. Функциональная роль индуцированных хозяином модификаций неясна.
Как видим, мутирование вирусов проходит достаточно сложный и тернистый путь в приобретении новых вирулентных свойств. Эти свойства могут быть как ослабляющими для развития инфекционного процесса, так и крайне агрессивными в своём новом виде.
Бактериофаги – это вирусы, которые поражают только бактерий. В ходе инфекции они влияют на все процессы жизнедеятельности бактериальной клетки, фактически превращая ее в фабрику по производству вирусного потомства. В конце концов клетка разрушается, а вновь образованные вирусные частицы выходят наружу и могут заражать новые бактерии.
Несмотря на огромное число и разнообразие природных фагов, встречаемся мы с ними редко. Однако бывают ситуации, когда деятельность этих вирусов не остается незамеченной. Например, на предприятиях, где производят сыры, йогурты и другие молочно-кислые продукты, часто приходится сталкиваться с вирусной атакой на бактерии, сбраживающие молоко. В большинстве таких случаев фаговая инфекция распространяется молниеносно, и полезные бактерии гибнут, что приводит к значительным экономическим потерям (Neve et al., 1994).
Именно благодаря прикладным исследованиям в интересах молочной промышленности, направленным на получение устойчивых к бактериофагам штаммов молочно-кислых бактерий, был открыт ряд механизмов, с помощью которых бактерии избегают инфекции. Параллельно были изучены способы, с помощью которых вирусы, в свою очередь, преодолевают бактериальные системы защиты (Moineau et al., 1993).
Кто защищен – тот вооружен
На сегодня известно пять основных, весьма хитроумных механизмов защиты, которые бактерии выработали в непрестанной борьбе с вирусами: изменение рецептора на поверхности клетки; исключение суперинфекции; системы абортивной инфекции; системы рестрикции-модификации и, наконец, системы CRISPR-Cas.
К средствам противовирусной защиты бактерий относятся и системы рестрикции-модификации, в которые входят гены, кодирующие два белка-фермента – рестриктазу и метилазу. Рестриктаза узнает определенные последовательности ДНК длиной 4—6 нуклеотидов и вносит в них двуцепочечные разрывы. Метилаза, напротив, ковалентно модифицирует эти последовательности, добавляя к отдельным нуклеотидным основаниям метильные группы, что предотвращает их узнавание рестриктазой.
Врага нужно знать в лицо
Системы CRISPR-Cas являются уникальным примером адаптивного иммунитета бактерий. При проникновении в клетку ДНК фага специальные белки Cas встраивают фрагменты вирусной ДНК длиной 25—40 нуклеотидов в определенный участок генома бактерии (Barrangou et al., 2007). Такие фрагменты называются спейсерами (от англ. spacer – промежуток), участок, где происходит встраивание, – CRISPR-кассета (от англ. Clustered Regularly Interspaced Short Palindromic Repeats), а сам процесс приобретения спейсеров – адаптацией.
Чтобы использовать спейсеры в борьбе с фаговой инфекцией, в клетке должен происходить еще один процесс, управляемый белками Cas, названный интерференцией. Суть его в том, что в ходе транскрипции CRISPR-кассеты образуется длинная молекула РНК, которая разрезается белками Cas на короткие фрагменты – защитные криспрРНК (крРНК), каждая из которых содержит один спейсер. Белки Cas вместе с молекулой крРНК образуют эффекторный комплекс, который сканирует всю ДНК клетки на наличие последовательностей, идентичных спейсеру (протоспейсеров). Найденные протоспейсеры расщепляются белками Cas (Westra et al., 2012; Jinek et al., 2012).
Системы CRISPR-Cas обнаружены у большинства прокариот – бактерий и архей. Хотя общий принцип действия всех известных систем CRISPR-Cas одинаков, механизмы их работы могут существенно отличаться в деталях. Наибольшие различия проявляются в строении и функционировании эффекторного комплекса, в связи с чем системы CRISPR-Cas делят на несколько типов. На сегодняшний день описаны шесть типов таких неродственных друг другу систем (Makarova et al., 2015; Shmakov et al., 2015).
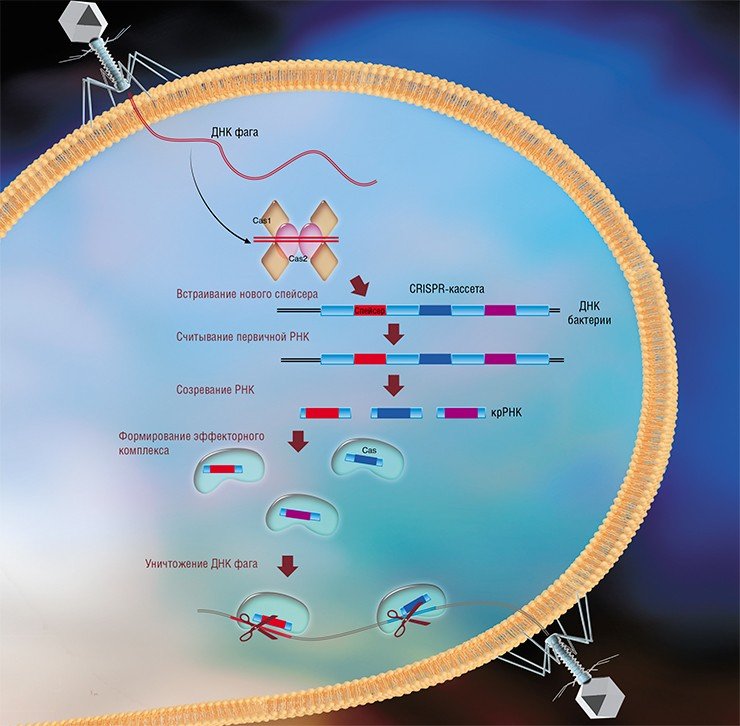
Наиболее изученной является система CRISPR-Cas I типа, которой обладает излюбленный объект молекулярно-биологических исследований – бактерия кишечная палочка (Esсherichia coli). Эффекторный комплекс в этой системе состоит из нескольких небольших белков Cas, каждый из которых отвечает за разные функции: разрезание длинной некодирующей CRISPR РНК, связывание коротких крРНК, поиск, а затем разрезание ДНК-мишени.
Гонка вооружений
Бактериофаги, как факторы среды, вызывают направленные изменения в геноме бактерий, которые наследуются и дают бактериям явное преимущество, спасая от повторных инфекций. Поэтому системы CRISPR-Cas можно считать примером ламарковской эволюции, при которой происходит наследование благоприобретенных признаков (Koonin et al., 2009)
Некоторые бактериофаги реагируют на наличие в бактериальной клетке систем CRISPR-Cas выработкой особых анти CRISPR-белков, способных связываться с белками Cas и блокировать их функции (Bondy-Denomy et al., 2015). Еще одно ухищрение — обмен участков генома вируса, на которые нацелена система CRISPR-Cas, на участки геномов родственных вирусов, отличающихся по составу нуклеотидной последовательности (Paez-Espino et al., 2015).
Благодаря постоянному совершенствованию биоинформатических алгоритмов поиска, а также включению в анализ все большего количества прокариотических геномов, открытие новых типов CRISPR-Cas систем является делом недалекого будущего. Предстоит также выяснить и детальные механизмы работы многих недавно открытых систем. Так, в статье, опубликованной в 2016 г. в журнале Science и посвященной анализу системы CRISPR-Cas VI типа, описан белок С2с2, образующий эффекторный комплекс с крРНК, который нацелен на деградацию не ДНК, а РНК (Abudayyeh et al., 2016). В будущем такое необычное свойство может быть использовано в медицине для регулирования активности генов путем изменения количества кодируемых ими РНК.
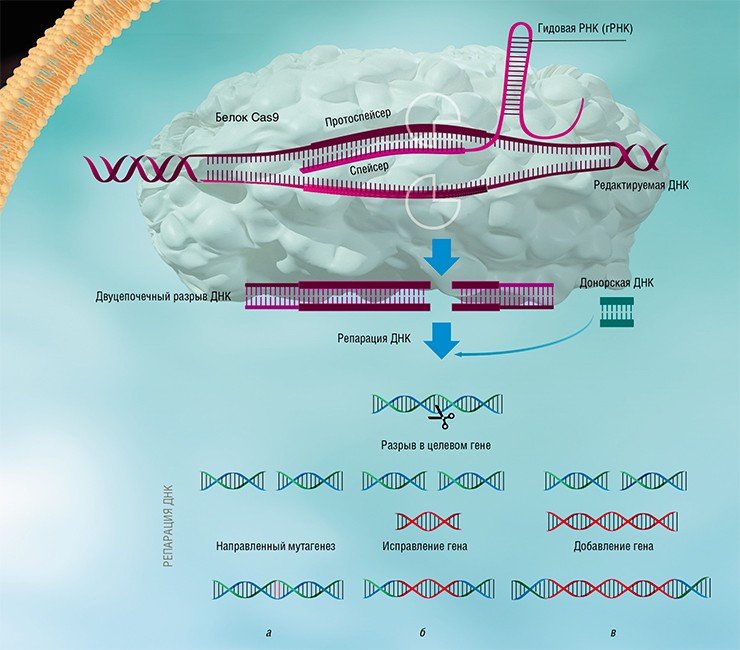
Изучение стратегий борьбы бактерий с бактериофагами, несмотря на свою кажущуюся фундаментальность и отвлеченность от задач практической медицины, принесло неоценимую пользу человечеству. Примерами этого могут служить методы молекулярного клонирования и редактирования геномов – направленного внесения или удаления мутаций и изменения уровня транскрипции определенных генов.
Благодаря быстрому развитию методов молекулярной биологии всего лишь через несколько лет после открытия механизма действия систем CRISPR-Cas была создана работающая технология геномного редактирования, способная бороться с болезнями, ранее считавшимися неизлечимыми. Доступность и простота этой технологии позволяют рассматривать ее как основу для медицины, ветеринарии, сельского хозяйства и биотехнологий будущего, которые будут базироваться на направленных и безопасных генных модификациях.
Нет никаких сомнений, что дальнейшее изучение взаимодействия бактерий и их вирусов может открыть перед нами такие возможности, о которых мы сейчас даже не подозреваем.
Abudayyeh O. O., Gootenberg J. S., Konermann S. et al. C 2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector // Science. 2016. V. 353: aaf5573.
Barrangou R., Fremaux C., Deveau H. et al. CRISPR provides acquired resistance against viruses in prokaryotes // Science. 2007. V. 315. P. 1709–1712.
Bikard D., Marraffini L. A. Innate and adaptive immunity in bacteria: mechanisms of programmed genetic variation to fight bacteriophages // Curr. Opin. Immunol. 2012. V. 1 P. 15–20.
Bondy-Denomy J., Garcia B., Strum S. et al. Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins // Nature. 2015. V. 526. P. 136–139.
Calendar R., Abedon S. T. The Bacteriophages // 2nd Ed., Oxford University Press. 2006.
Datsenko K. A., Pougach K., Tikhonov A. et al. Molecular memory of prior infections activates the CRISPR/Cas adaptive bacterial immunity system // Nat. Commun. 2012. V. 3. P. 945
Jiang W., Marraffini L. A. CRISPR-Cas: New Tools for Genetic Manipulations from Bacterial Immunity Systems // Annu. Rev. Microbiol. 2015. V. 69. P. 209–28.
Jinek M., Chylinski K., Fonfara I., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity // Science. 2012. V. 337. P. 816–821.
Koonin E. V., Wolf Y. I. Is evolution Darwinian or/and Lamarckian? // Biol. Direct. 2009. V. 4. P. 42.
Lopez-Pascua L., Buckling A. Increasing productivity accelerates host-parasite coevolution // J. Evol. Biol. 2008. V. 3. P. 853–860.
Makarova K. S., Wolf Y. I., et al. An updated evolutionary classification of CRISPR-Cas systems // Nat. Rev. Microbiol. 2015. V. 11. P. 722–736.
Moineau, S., Pandian S., Klaenhammer T. R. Restriction/modification systems and restriction endonucleases are more effective on lactococcal bacteriophages that have emerged recently in the dairy industry // Appl. Envir. Microbiol. 1993. V. 59. P. 197–202.
Neve H., Kemper U., et al. Monitoring and characterization of lactococcal bacteriophage in a dairy plant // Kiel. Milckwirtsch. Forschungsber. 1994. V. 46. P. 167–178.
Nuñez J. K., Harrington L. B., et al. Foreign DNA capture during CRISPR-Cas adaptive immunity // Nature. 2015a. V. 527. P. 535–538.
Nuñez J. K., Kranzusch P. J., et al. Cas1-Cas2 complex formation mediates spacer acquisition during CRISPR-Cas adaptive immunity // Nat. Struct. Mol. Biol. 2014. V. 21. P. 528–534.
Nuñez J. K., Lee A. S., Engelman A., Doudna J. A. Integrase-mediated spacer acquisition during CRISPR-Cas adaptive immunity // Nature. 2015b. V. 519. P. 193–198.
Paez-Espino D., Sharon I., et al. CRISPR Immunity Drives Rapid Phage Genome Evolution in Streptococcus thermophilus // MBio. 2015. V. 6: e00262–15.
Shmakov S., Abudayyeh O. O., Makarova K. S., et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. // Mol. Cell. 2015. V. 60. P. 385–397
Tan D., Svenningsen S. L., Middelboe M. Quorum sensing determines the choice of antiphage defense strategy in Vibrio anguillarum. // mBio 2015. V. 6: e00627–15.
Westra E. R., van Erp P. B., Künne T., et al. CRISPR immunity relies on the consecutive binding and degradation of negatively supercoiled invader DNA by Cascade and Cas3 // Mol. Cell. 2012. V. 46. P. 595–605.
Читайте также: