
Особенности морфологии биологии и классификации вирусов
Обновлено: 24.04.2024
Систематика вирусов. Особенности классификации вирусов. Основные критерии таксономической классификации вирусов.
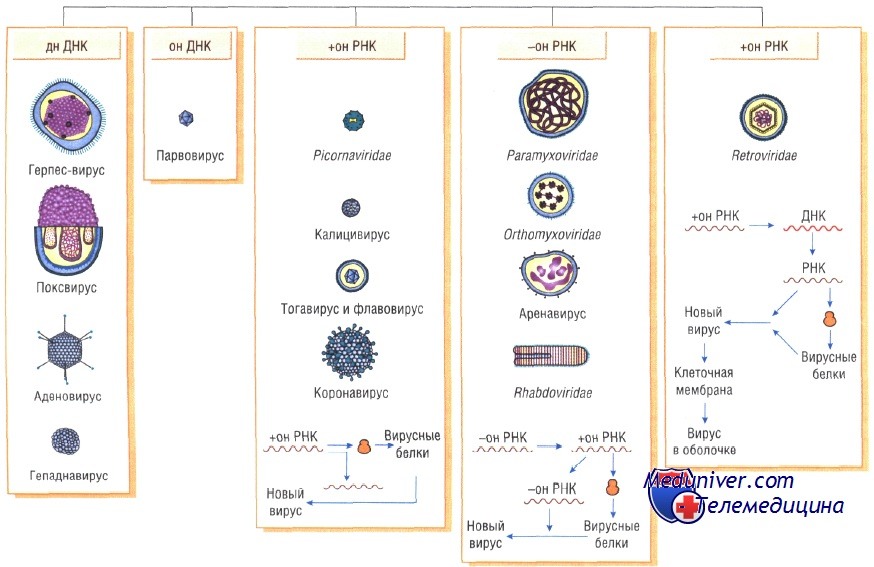
Вирусы отнесены к царству Vira. В основу их классификации положен тип нуклеиново кислоты, образующей геном. Соответственно выделяют рибовирусы (РНК-вирусы) и дезоксирибовирусы (ДНК-вирусы).
Для вирусов предложены следующие таксономические категории (по восходящей): Вид (Species) —> Род (Genus) —> Подсемейство (Subfamilia) —> Семейство (Familia). Но категории подсемейств и родов разработаны не для всех вирусов. Видовые названия вирусов обычно связывают с вызываемыми ими заболеваниями (например, вирус бешенства) либо по названию места, где они были впервые выделены (например, вирусы Коксаки, вирус Эбола). Если семейство включает большое количество видов, то видовые названия дают в соответствии с антигенной структурой и разделяют их на типы (например, аденовирус 32 типа или вирус герпеса 1 типа). Реже используют фамилии учёных, впервые их выделивших (например, вирус Эпстайна-Барр или вирус саркомы Рауса). Иногда используют устаревшие названия групп вирусов, отражающих их уникальные эпидемиологические характеристики (например, арбовирусы). Классификационные признаки патогенных для человека вирусов приведены в табл. 3-1.
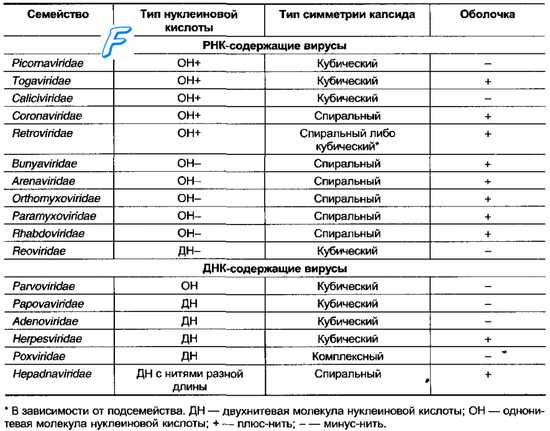
Таблица 3-1. Семейства вирусов, включающие возбудителей инфекций человека
К вирусам отнесены вироиды [от virus и греч. eidos, сходство] — мелкие кольцевые однонитевые суперспирализованные молекулы РНК (аналогичную организацию имеет геном вируса гепатита D). Поскольку у вироидов нет белковой оболочки, они не проявляют выраженных иммуногенных свойств, и поэтому их нельзя идентифицировать серологическими методами. Вироиды вызывают заболевания у растений. В качестве безымянного таксона в царство Vira также включены и прионы.
Основные критерии таксономической классификации вирусов
При систематизировании вирусов выделяют следующие основные критерии: сходство нуклеиновых кислот, размеры, наличие или отсутствие суперкапсида, тип симметрии нуклеокапсида, характеристика нуклеиновых кислот (молекулярная масса, тип кислоты (ДНК или РНК), полярность [плюс или минус], количество нитей в молекуле либо наличие сегментов, наличие ферментов), чувствительность к химическим агентам (особенно к эфиру), антигенная структура и иммуногенность, тропизм к тканям и клеткам, способность образовывать тельца включений.
Дополнительный критерий — симптоматология поражений: способность вызывать генерализованные поражения либо инфекции с первичным поражением определённых органов (нейроинфекции, респираторные инфекции и др.).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царствуVira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом.
Морфологию и структурувирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Формавирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы.
Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид.
Сложные, или оболочечные, вирусы снаружи капсида окружены ли-попротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок.
Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии.Икосаэдрическийтип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита).Спиральныйтип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа).
Включения — скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Вирус натуральной оспы образует цитоплазмати-ческие включения — тельца Гварниери; вирусы герпеса и аденовирусы — внутриядерные включения.
Размерывирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм).
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Вирусы поражаютпозвоночных и беспозвоночных животных, а также растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы также участвуют в процессах канцерогенеза, могут передаваться различными путями, в том числе через плаценту (вирус краснухи, цитомега ловирус и др.), поражая плод человека. Они могут приводить к постинфекционным осложнениям — развитию миокардитов, панкреатитов, иммунодефицитов и др.
Кроме обычных вирусов, известны и так называемые неканонические вирусы — прионы — белковые инфекционные частицы, являющиеся агентами белковой природы, имеющие вид фибрилл размером 10—20x100—200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтц-фельдта—Якоба, куру и др.).
Другими необычными агентами, близкими к вирусам, являются вироиды — небольшие молекулы кольцевой, суперспи-рализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Открытие вирусов Д.И.Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому ее развитию способствовали: изобретение электронного микроскопа, разработка метода культивирования микроорганизмов в культурах клеток.
Слово “вирус” в переводе с латинского- яд (животного происхождения). Этот термин применяют для обозначения уникальных представителей живой природы, не имеющих клеточного (эукариотического или прокариотического) строения и обладающих облигатным внутриклеточным паразитизмом, т.е. которые не могут жить без клетки.
В настоящее время вирусология- бурно развивающаяся наука, что связано с рядом причин:
- ведущей ролью вирусов в инфекционной патологии человека (примеры- вирус гриппа, ВИЧ- вирус иммунодефицита человека, цитомегаловирус и другие герпесвирусы) на фоне практически полного отсутствия средств специфической химиотерапии;
- использованием вирусов для решения многих фундаментальных вопросов биологии и генетики.
Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого мира.
1.Ультрамикроскопические размеры (измеряются в нанометрах). Крупные вирусы (вирус оспы) могут достигать размеров 300 нм, мелкие- от 20 до 40 нм. 1мм=1000мкм, 1мкм=1000нм.
2.Вирусы содержат нуклеиновую кислоту только одного типа- или ДНК (ДНК- вирусы) или РНК (РНК- вирусы). У всех остальных организмов геном представлен ДНК, в них содержится как ДНК, так и РНК.
3.Вирусы не способны к росту и бинарному делению.
4.Вирусы размножаются путем воспроизводства себя в инфицированной клетке хозяина за счет собственной геномной нуклеиновой кислоты.
5.У вирусов нет собственных систем мобилизации энергии и белок- синтензирующих систем, в связи с чем вирусы являются абсолютными внутриклеточными паразитами.
6.Средой обитания вирусов являются живые клетки- бактерии (это вирусы бактерий или бактериофаги), клетки растений, животных и человека.
Все вирусы существуют в двух качественно разных формах: внеклеточной- вирион и внутриклеточной- вирус. Таксономия этих представителей микромира основана на характеристике вирионов- конечной фазы развития вирусов.
Строение (морфология) вирусов.
1.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК- вирусов) или двухцепочечными молекулами ДНК (у большинства ДНК- вирусов).
2.Капсид - белковая оболочка, в которую упакована геномная нуклеиновая кислота. Капсид состоит из идентичных белковых субъединиц- капсомеров. Существуют два способа упаковки капсомеров в капсид- спиральный (спиральные вирусы) и кубический (сферические вирусы).
При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также по спирали, уложена геномная нуклеиновая кислота (нитевидные вирусы). При кубическом типе симметрии вирионы могут быть в виде многогранников, чаще всего- двадцатигранники - икосаэдры.
3.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и называются “голыми”.
4. У других вирусов поверх капсида есть дополнительная мембраноподобная оболочка, приобретаемая вирусом в момент выхода из клетки хозяина- суперкапсид. Такие вирусы называют “одетыми”.
Кроме вирусов, имеются еще более просто устроенные формы способных передаваться агентов - плазмиды, вироиды и прионы.
Основные этапы взаимодействия вируса с клеткой хозяина.
1.Адсорбция- пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина (у вируса гриппа- гемагглютинин, у вируса иммунодефицита человека- гликопротеин gp 120).
2.Проникновение- путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза).
3.Освобождение нуклеиновых кислот- “раздевание” нуклеокапсида и активация нуклеиновой кислоты.
4.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса.
5.Сборка вирионов- ассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком.
6.Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами.
Исходы взаимодействия вирусов с клеткой хозяина.
1.Абортивный процесс- когда клетки освобождаются от вируса:
- при инфицировании дефектным вирусом, для репликации которого нужен вирус- помощник, самостоятельная репликация этих вирусов невозможна ( так называемые вирусоиды). Например, вирус дельта (D) гепатита может реплицироваться только при наличии вируса гепатита B, его Hbs - антигена, аденоассоциированный вирус- в присутствии аденовируса);
- при инфицировании вирусом генетически нечувствительных к нему клеток;
- при заражении чувствительных клеток вирусом в неразрешающих условиях.
2.Продуктивный процесс- репликация (продукция) вирусов:
- гибель (лизис) клеток (цитопатический эффект)- результат интенсивного размножения и формирования большого количества вирусных частиц - характерный результат продуктивного процесса, вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит достаточно узнаваемый специфический характер;
- стабильное взаимодействие, не приводящее к гибели клетки (персистирующие и латентные инфекции) - так называемая вирусная трансформация клетки.
3.Интегративный процесс- интеграция вирусного генома с геномом клетки хозяина. Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК- геном хозяина могут только ДНК- вирусы (принцип “ДНК- в ДНК”). Единственные РНК- вирусы, способные интегрироваться в геном клетки хозяина- ретровирусы, имеют для этого специальный механизм. Особенность их репродукции- синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы с последующим встраиванием ДНК в геном хозяина.
Основные методы культивирования вирусов.
1.В организме лабораторных животных.
2.В куриных эмбрионах.
3.В клеточных культурах - основной метод.
Типы клеточных культур.
1.Первичные (трипсинизированные) культуры- фибробласты эмбриона курицы (ФЭК), человека (ФЭЧ), клетки почки различных животных и т.д. Первичные культуры получают из клеток различных тканей чаще путем их размельчения и трипсинизации, используют однократно, т.е. постоянно необходимо иметь соответствующие органы или ткани.
2.Линии диплоидных клеток пригодны к повторному диспергированию и росту, как правило не более 20 пассажей (теряют исходные свойства).
3.Перевиваемые линии (гетероплоидные культуры), способны к многократному диспергированию и перевиванию, т.е. к многократным пассажам, наиболее удобны в вирусологической работе- например, линии опухолевых клеток Hela, Hep и др.
Специальные питательные среды для культур клеток.
Используются разнообразные синтетические вирусологические питательные среды сложного состава, включающие большой набор различных факторов роста- среда 199, Игла, раствор Хэнкса, гидролизат лактальбумина. В среды добавляют стабилизаторы рН (Hepes), различные в видовом отношении сыворотки крови (наиболее эффективной считают эмбриональную телячью сыворотку), L-цистеин и L-глютамин.
В зависимости от функционального использования среды могут быть ростовые (с большим содержанием сыворотки крови) - их используют для выращивания клеточных культур до внесения вирусных проб, и поддерживающие (с меньшим содержанием сыворотки или ее отсутствием)- для содержания инфицированных вирусом клеточных культур.
Выявляемые проявления вирусной инфекции клеточных культур.
2.Выявление телец включений.
3. Выявление вирусов методом флюоресцирующих антител (МФА), электронной микроскопией, авторадиографией.
4.Цветная проба. Обычный цвет используемых культуральных сред, содержащих в качестве индикатора рН феноловый красный, при оптимальных для клеток условиях культивирования (рН около 7,2)- красный. Размножение клеток меняет рН и соответственно- цвет среды с красного на желтый за счет смещения рН в кислую сторону. При размножении в клеточных культурах вирусов происходит лизис клеток, изменения рН и цвета среды не происходит.
5.Выявление гемагглютинина вирусов- гемадсорбция, гемагглютинация.
6.Метод бляшек (бляшкообразования). В результате цитолитического действия многих вирусов на клеточные культуры образуются зоны массовой гибели клеток. Выявляют бляшки- вирусные “ клеточно- негативные” колонии.
Название семейства вирусов заканчивается на “viridae”, рода- “virus”, для вида обычно используют специальные названия, например - вирус краснухи, вирус иммунодефицита человека- ВИЧ, вирус парагриппа человека типа 1 и т.д.
Вирусы бактерий (бактериофаги).
Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например, в сточных водах). Фагам присущи биологические особенности, свойственные и другим вирусам.
Наиболее морфологически распространенный тип фагов характеризуется наличием головки- икосаэдра, отростка (хвоста) со спиральной симметрией (часто имеет полый стержень и сократительный чехол), шипов и отростков (нитей), т.е. внешне несколько напоминают сперматозоид.
Взаимодействие фагов с клеткой (бактерией) строго специфично, т.е. бактериофаги способны инфицировать только определенные виды и фаготипы бактерий.
Основные этапы взаимодействия фагов и бактерий.
1.Адсорбция (взаимодействие специфических рецепторов).
2.Внедрение вирусной ДНК (инъекция фага) осуществляется за счет лизирования веществами типа лизоцима участка клеточной стенки, сокращения чехла, вталкивания стержня хвоста через цитоплазматическую мембрану в клетку, впрыскивание ДНК в цитоплазму.
4.Выход дочерних популяций.
Основные свойства фагов.
Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие редуктивную фаговую инфекцию (редукцию фага). В последнем случае геном фага в клетке не не реплицируется, а внедряется (интегрируется) в хромосому клетки хозяина (ДНК в ДНК), фаг превращается в профаг. Этот процесс получил название лизогении. Если в результате внедрения фага в хромосому бактериальной клетки она приобретает новые наследуемые признаки, такую форму изменчивости бактерий называют лизогенной (фаговой) конверсией. Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг при нарушении синтеза особого белка- репрессора может перейти в литический цикл развития, вызвать продуктивную инфекцию с лизисом бактерии.
Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями- в трансдукции (одна из форм генетического обмена). Например, способностью вырабатывать экзотоксин обладают только возбудитель дифтерии, в хромосому которого интегрирован умеренный профаг, несущий оперон tox, отвечающий за синтез дифтерийного экзотоксина. Умеренный фаг tox вызывает лизогенную конверсию нетоксигенной дифтерийной палочки в токсигенную.
По спектру действия на бактерии фаги разделяют на :
- поливалентные (лизируют близкородственные бактерии, например сальмонеллы);
- моновалентные (лизируют бактерии одного вида);
- типоспецифические (лизируют только определенные фаговары возбудителя).
На плотных средах фаги обнаруживают чаще с помощью спот (spot) - теста (образование негативного пятна при росте колоний) или методом агаровых слоев (титрования по Грациа).
Практическое использование бактериофагов.
1.Для идентификации (определение фаготипа).
2.Для фагопрофилактики (купирование вспышек).
3.Для фаготерапии (лечение дисбактериозов).
4.Для оценки санитарного состояния окружающей среды и эпидемиологического анализа.
Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных.
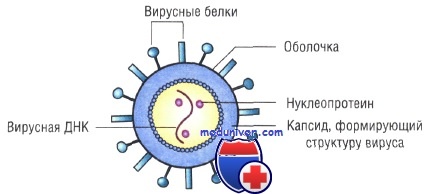
Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
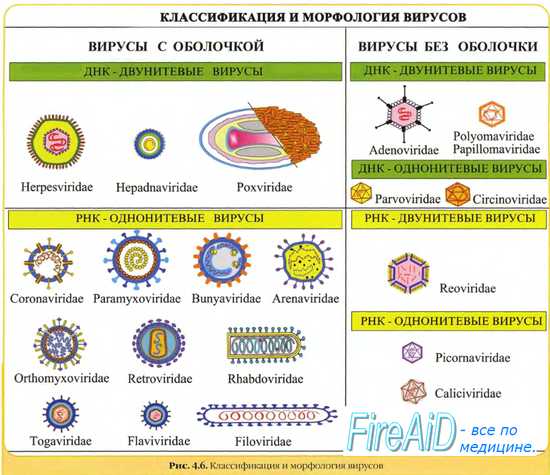
Морфология вирусов. Размеры вирусов.
Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки.
Нуклеиновые кислоты вирусов
Вирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине.
Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180") повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
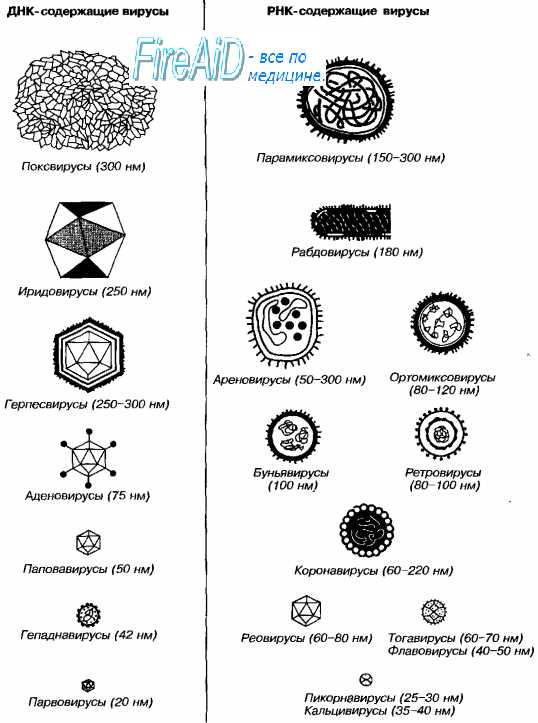
Рис. 2-1. Размеры и морфология основных возбудителей вирусных инфекций человека.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК.
Инфекционность нуклеиновых кислот вирусов
Многие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Строение вирусов. Классификация вирусов
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: