
Особенности строения генетического аппарата у бактерий и вирусов
Обновлено: 18.04.2024
В состав нуклеотида бактерий входят ДНК, РНК и белки. Число нуклеотидов в бактериальной клетке может варьировать от одного (в культурах, находящихся в стационарной фазе роста) до двух (в стадии задержки размножения после переноса клеток в свежую среду) и четырех (в культурах с постоянной скоростью роста). Каждый нуклеотид содержит двухцепочечную замкнутую в кольцо молекулу ДНК. В молекуле ДНК нуклеотида закодирована вся генетическая информация, необходимая для жизнедеятельности клетки, поэтому нуклеотид рассматривают как бактериальную хромосому. Хромосомы имеют кольцевое строение. Гигантская молекула ДНК бактериальной хромосомы поддерживается связанными с ней молекулами РНК и белка в форме компактной структуры, свернутой в отдельные сверхспирализованные петли (домены), число которых колеблется от 12 до 80.
Помимо хромосомной ДНК в состав генома многих прокариот входят также сверхскрученные, ковалентно-замкнутые кольцевые молекулы внехромосомной, или плазмидной, ДНК.
Способы передачи наследственной информации у бактерий:
1. трансформация – перенос изолированных фрагментов молекулы ДНК из одного организма к другому.
2. трансдукция это способность переносить наследственную информацию от одного организма к другому при помощи вирусов.
3. конъюгация – обмен наследственной информацией.
Генетический материал вируса представлен одной молекулой нуклеиновой кислоты, ДНК или РНК, не связанной с белком. В связи с этим вирусы подразделяются на ДНК- и РНК-содержащие. Вирусы бактерий чаще содержат ДНК, а почти все вирусы растений и подавляющее большинство вирусов человека – РНК.
Нуклеиновая кислота вируса бывает одно- или двухцепочечной и может иметь кольцевую или линейную форму. Кольцевая форма ДНК более стабильна и свойственна большинству вирусов. Кольцо ДНК (РНК) обычно бывает перекручено, поэтому она имеет суперспирализованный вид.
Генетическое вещество у вирусов заключено в белковую оболочку, которая вместе с нуклеиновой кислотой образует так называемый капсид или нуклеокапсид. Большинство вирусов растений и РНК-содержащих бактериальных фагов состоит только из нуклеиновой кислоты и белка.
29. Определение понятия "фенотип" и формы фенотипической изменчивости. Фенотипическая изменчивость у эукариот, формы проявления.
Фенотип бактерии – результат взаимодействия между бактерией и окружающей средой, который контролирует геном.
Фенотипическая изменчивость — модификация — не затрагивает генотип, но затрагивает большинство особей популяции. Модификации не передаются по наследству и с течением времени затухают, т. е. возвращаются к исходному фенотипу через большее (длительные модификации) или меньшее (кратковременные модификации) число поколений.
Они могут возникать в популяции любого вида, и их проявления в мире бактерий наблюдают довольно часто. Они в целом контролируются генофором бактерий, но (в отличие от мутаций) не сопровождаются изменениями кодирующей структуры и утрачиваются при прекращении действия вызвавших их факторов. У бактерий наблюдают морфологические (приводящие к обратимым изменениям формы) и биохимические (приводящие к синтезу некоторых продуктов, чаще ферментов) модификации. По существу, модификации возникают как адаптивные реакции бактериальных клеток на изменения окружающей среды, что позволяет им быстро приспосабливаться благодаря чему сохраняется определённая численность популяции. После устранения соответствующего воздействия, вызвавшего их образование, бактерии возвращаются к исходному фенотипу. Примером адаптации микроорганизмов может служить способность патогенных микроорганизмов образовывать под действием пенициллина L-формы, у которых отсутствует клеточная стенка, служащая мишенью для пенициллина.
• В клетках большинства прокариот присутствует одна кольцевая хромосома
• Генетическая гибкость прокариот и их адаптационные возможности усиливаются при заражении бактериофагами и при переносе плазмид
• Быстрая эволюция генома прокариот обеспечивается транспозонами и другими мобильными элементами
У большинства прокариот присутствует одна кольцевая хромосома, и они являются гаплоидными клетками. В следующем разделе рассмотрены вопросы организации хромосомы в дискретную структуру, нуклеоид. В настоящее время мы располагаем данными секвенирования более 200 геномов прокариот, размеры которых варьируют от 580 кнп (Mycoplasma genitalium) до 9 Мнп (Streptomyces, Myxococcus).
Геном таких хорошо известных бактерий, как Е. coli и В. subtilis, занимает среднее положение (4-5 Мнп). Относительно небольшие размеры прокариотических хромосом, по сравнению с хромосомами высших эукариот, объясняются их компактностью и небольшим количеством некодирующих последовательностей ДНК. В общем, гены, необходимые для постоянного роста и поддержания жизнеспособности прокариот, находятся в хромосоме, а генетическая гибкость микроорганизмов обеспечивается различными мобильными элементами.
У некоторых бактерий хромосома имеет линейную форму, или же в клетке содержится несколько таких хромосом. Встречаются виды, для которых характерны обе эти особенности. Например, клетки Streptomyces содержат линейную хромосому. Концы этой хромосомы замкнуты белковым мостиком, и это объясняет, почему долгое время на генетической карте этих бактерий изображали круговую хромосому.
Бактерии Rhodobacter sphaeroides обладают двумя большими кольцевыми хромосомами (3,0 и 0,9 Мнп), в каждой из которых присутствует много необходимых генов домашнего хояйства. Возбудитель Лайм-боррелиоза, бактерия Воrrelia burgdorferi характеризуется наличием нескольких линейных хромосом.
Стабильные элементы внехромосомной ДНК, которые не несут необходимых генов домашнего хозяйства и поэтому не являются существенными, называются плазмиды. На рисунке ниже перечислены некоторые хорошо изученные плазмиды бактерий. Эти данные дают представление о размерах генов, которые могут нести эти плазмиды. Плазмиды обычно невелики, варьируя по размерам от 2 до 1000 кнп, и обладают кольцевой структурой.
Так же как и в случае хромосом, известны исключения, и некоторые крупные плазмиды достигают размера 1 Мнп или больше, и небольшая их часть обладает линейной структурой. Все плазмиды несут гены, управляющие их репликацией, обычно включающей различные элементы аппарата репликации клетки хозяина. К числу важнейших генов, которые несут плазмиды, относятся гены устойчивости к антибиотикам и обусловливающие патогенные свойства или деградацию необычных источников углерода.
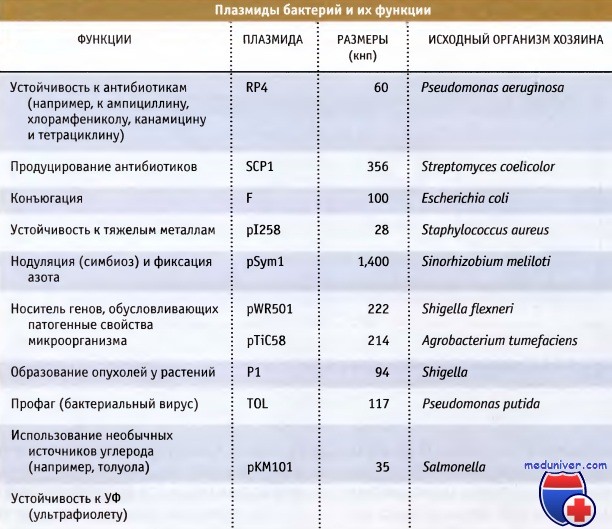
Плазмиды бактерий и их функции.
Плазмиды могут распространяться между организмами различными путями. Конъюгация представляет собой процесс, напоминающий спаривание, при котором присходит прямой перенос ДНК от клетки донора, содержащей плазмиду, в клетку реципиента. В донорской плазмиде закодированы функции, необходимые дляУ большинства прокариот присутствует одна кольцевая хромосома, и они являются гаплоидными клетками. В следующем разделе рассмотрены вопросы организации хромосомы в дискретную структуру, нуклеоид.
В настоящее время мы располагаем данными секвенирования более 200 геномов прокариот, размеры которых варьируют от 580 кнп (Mycoplasma genitalium) до 9 Мнп (Streptomyces, Myxococcus). Геном таких хорошо известных бактерий, как Е. coli и В. subtilis, занимает среднее положение (4-5 Мнп). Относительно небольшие размеры прокариотических хромосом, по сравнению с хромосомами высших эукариот, объясняются их компактностью и небольшим количеством некодирующих последовательностей ДНК. В общем, гены, необходимые для постоянного роста и поддержания жизнеспособности прокариот, находятся в хромосоме, а генетическая гибкость микроорганизмов обеспечивается различными мобильными элементами.
У некоторых бактерий хромосома имеет линейную форму, или же в клетке содержится несколько таких хромосом. Встречаются виды, для которых характерны обе эти особенности. Например, клетки Streptomyces содержат линейную хромосому. Концы этой хромосомы замкнуты белковым мостиком, и это объясняет, почему долгое время на генетической карте этих бактерий изображали круговую хромосому.
Бактерии Rhodobacter sphaeroides обладают двумя большими кольцевыми хромосомами (3,0 и 0,9 Мнп), в каждой из которых присутствует много необходимых генов домашнего хояйства. Возбудитель Лайм-боррелиоза, бактерия Воrrelia burgdorferi характеризуется наличием нескольких линейных хромосом.
Стабильные элементы внехромосомной ДНК, которые не несут необходимых генов домашнего хозяйства и поэтому не являются существенными, называются плазмиды. На рисунке ниже перечислены некоторые хорошо изученные плазмиды бактерий. Эти данные дают представление о размерах генов, которые могут нести эти плазмиды. Плазмиды обычно невелики, варьируя по размерам от 2 до 1000 кнп, и обладают кольцевой структурой.
Так же как и в случае хромосом, известны исключения, и некоторые крупные плазмиды достигают размера 1 Мнп или больше, и небольшая их часть обладает линейной структурой. Все плазмиды несут гены, управляющие их репликацией, обычно включающей различные элементы аппарата репликации клетки хозяина. К числу важнейших генов, которые несут плазмиды, относятся гены устойчивости к антибиотикам и обусловливающие патогенные свойства или деградацию необычных источников углерода.
Плазмиды могут распространяться между организмами различными путями. Конъюгация представляет собой процесс, напоминающий спаривание, при котором присходит прямой перенос ДНК от клетки донора, содержащей плазмиду, в клетку реципиента. В донорской плазмиде закодированы функции, необходимые для обеспечения контакта с клеткой реципиентом, инициации переноса реплицированной ДНК, и самого переноса ДНК в клетку реципиента.
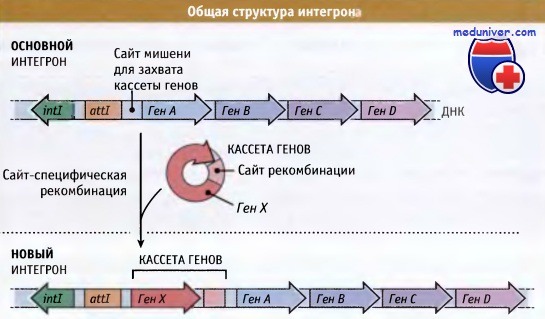
Структура типичного интегрона после накопления нескольких кассет генов.
Плазмиды также могут передаваться при прямом захвате ДНК (трансформация) или с участием бактериофага (трансдукция). Независимо от наличия экстрахромосомных элементов, изменения в бактериальном геноме могут возникать за счет процессов гомологичной и сайт-специфической рекомбинации.
Для многих бактерий важными источниками генетической вариабельности, по-видимому, являются бактериофаги (бактериальные вирусы). Результаты секвенирования показывают, что в геноме многих бактерий присутствуют интегрированные последовательности бактериофага (профаги). Например, у Е. coli их по меньшей мере 9, а у B. subtilis 10. Некоторые из этих профагов обладают дефектами, например у них присутствуют делеции и другие мутации, что делает маловероятным их дальнейшую активацию с образованием инфекционных бактериофагов.
В некоторых случаях профаги несут гены, которые обеспечивают некоторые преимущества клеткам хозяина: это относится к системам рестрикции и модификации, устойчивости к УФ и к таким детерминантом патогенности, как токсины.
Наконец, бактериальный геном также содержит много мобильных генетических элементов, которые распространяются посредством транспозиции. Инсерционные последовательности содержат минимальные элементы, которые, в своей простейшей форме, вместе с фланкирующими последовательностями, обладают только одним геном транспозазы. При инициации транспозиции, фланкирующие последовательности узнаются белком транспозазой.
Затем происходит мобилизация ферментов клетки хозяина, участвующих в репликации и репарации ДНК. Они завершают процесс вставки последовательности в необходимом месте. Более сложные транспозоны несут дополнительные гены, способные обеспечить адаптационные преимущества клеткам хозяина. Наиболее известными примерами транспозонов у бактерий являются гены устойчивости к антибиотикам, однако подобным образом могут переноситься также многие другие гены. Близкие к транспозонам элементы могут катализировать несколько таких типов перегруппировок ДНК, как инверсии и делеции.
Интегроны представляют собой особенно важный инструмент адаптивной перегруппировки генома. Как показано на рисунке ниже, интегроны обычно состоят из гена интегразы, примыкающего к нему сайта мишени для захвата кассеты генов, и сильного промотора, регулирующего экспрессию захваченных генов. В кассетах, часто включающих гены устойчивости к антибиотикам, содержатся последовательности, которые позволяют им включаться на место мишени под действием белка интегразы. Интегроны могут расти за счет последовательного захвата различных кассет генов, что способствует быстрому развитию таких бактерий, которые устойчивы к ряду антибиотиков.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Клетка прокариот бактерий и архей
Основные положения:
• В прокариотической клетке плазматическая мембрана окружает один компартмент
• Во всем компартменте присутствует одинаковая водная среда
• В клетке генетический материал занимает компактную область
• Бактерии и археи относятся к прокариотам, однако различаются по своим структурным особенностям
Прокариоты подразделяются на два царства. Раньше считали, что все прокариоты представлены бактериями, но сейчас часть их мы причисляем к археям. Как бактерии, так и археи существуют в форме только одноклеточных организмов (хотя некоторые бактерии в популяции проявляют способность к агрегации).
Область, ограниченная плазматической мембраной, называется цитоплазмой. У прокариот мембрана окружена клеточной стенкой, жесткая структура которой обеспечивает защиту клетки от физических воздействий внешней среды.
На рисунке ниже показано, что в компартменте бактериальной клетки генетический материал расположен компактно, однако не отделен мембраной от содержимого цитоплазмы. К простейшим формам бактерий относится микоплазма, которая, однако, не способна к самостоятельному существованию, поскольку не может производить многие из жизненно необходимых продуктов.
У бактерий существует один компартмент, хотя внутренние области могут отличаться друг от друга.
Поэтому микоплазма существует внутри других организмов, в которых эти продукты образуются. В геноме микоплазмы содержится всего лишь около 500 генов, которые кодируют лишь минимальное количество продуктов, необходимых для построения клетки. Геном свободноживущих бактерий содержит более 1500 генов и кодирует синтез ферментов метаболизма, необходимых для превращения небольших молекул, а также обеспечивает функционирование более сложного аппарата регуляции экспрессии генов.
Бактерии подразделяются на две группы, дивергенция между которыми произошла, вероятно, около двух миллиардов лет назад. Эти группы называются грамположительные и грамотрицательные, в зависимости от того, приобретают ли клетки окраску при прокрашивании по Граму. К числу наиболее полно охарактеризованных грамотрицательных бактерий относится Escherichia coli, а из грамположительных бактерий наиболее изучена Bacillus subtilis. Окраска развивается при взаимодействии красителя с клеточной стенкой.
У грамположительных бактерий клеточная стенка окружает плазматическую мембрану, и краситель непосредственно взаимодействует с компонентами стенки. У грамотрицательных бактерий существует вторая мембрана, окружающая клеточную стенку. Наличие этой мембраны и различия в составе клеточной стенки препятствуют развитию окраски. Область, находящаяся между наружной и внутренней мембранами, называется периплазматическим пространством. В этом пространстве находятся специфические белки и другие компоненты. Если за критерий компартмента принимать область, ограниченную мембранами, то можно считать, что грамотрицательные бактерии имеют два компартмента.
Однако периплазматическое пространство следует рассматривать как компартмент лишь в аспекте взаимодействия между клеткой и окружающей средой. Это никак не сказывается на основополагающем факте, что синтетическая активность бактериальной клетки сосредоточена в том же компартменте, где находится генетический материал.
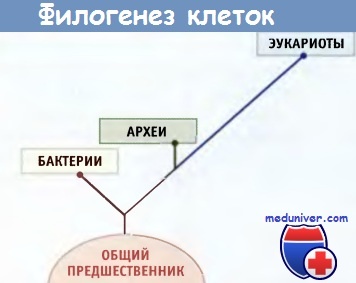
Данные филогенетического анализа с использованием современных молекулярных методов позволяют считать,
что организмы можно подразделить на три царства.
Некоторые бактерии могут развиваться, давая начало определенному типу специализированных клеток, что напоминает процесс развития у высших организмов.
Известно много различных видов бактерий, которые возникли на ранних этапах эволюции. Установить их филогенетические взаимоотношения достаточно сложно, поскольку, в отличие от эукариот, ископаемых остатков не сохранилось. Однако современные молекулярные методы, основанные на секвенировании рибосо-мальных РНК, и недавно разработанные приемы полного секвенирования генома привели к революционным выводам относительно происхождения прокариот. Как отдельное царство прокариот были идентифицированы археи.
По виду и строению археи напоминают бактерии: они характеризуются небольшими размерами и представляют собой одноклеточные организмы. Обычно они существуют в экстремальных условиях (например, при высоких температурах), и раньше их ошибочно принимали за бактерии, которые приспособились к таким условиям существования. Как и клетки бактерий, археи представляют собой клетки с одним компартментом и не имеют внутренних мембран.
У них могут проявляться такие же морфологические признаки, как у бактерий, например наличие жесткой стенки или капсулы, окружающей плазматическую мембрану, а также жгутиков, направленных в окружающую среду. Основные отличия наблюдаются на молекулярном уровне, и компоненты клетки археев отличаются от таковых у бактерий. Аппарат, осуществляющий экспрессию генов у археев, больше напоминает аналогичный аппарат клеток эукариот, чем клеток бактерий. Клеточная стенка у них построена из субъединиц, отличающихся от субъединиц клеточной стенки бактерий или растений. Существуют отличия в составе мембранных липидов. По генетической сложности археи больше напоминают свободно-живущих бактерий.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Структуры передачи наследственной информации клеток
Основные положения:
• ДНК переносит генетическую информацию, в которой закодирована последовательность всех клеточных белков.
• Информация может переноситься и в клеточных структурах которые также наследуются.
В каждой живой клетке двойная спираль ДНК несет основную наследственную информацию. У бактерий и архей все информационные последовательности обычно расположены в одной хромосоме. У эукариот почти все гены находятся в ядерных хромосомах, и небольшое количество информационных последовательностей локализовано в митохондриях и хлоропластах (у растений).
ДНК также может являться генетическим материалом вирусов, однако у некоторых вирусов таковым служит РНК. У всех вирусов генетический материал окружен белковой оболочкой. Конечно, вирусы не являются живыми организмами, однако при инфицировании клетки их генетический аппарат функционирует таким же образом, как и у клетки-хозяина.
Клетки также способны хранить информацию, которая не закодирована в последовательностях ДНК. Такой способ наследования называется эпигенетической наследственностью. Формально этим термином описывается ситуация, когда две клетки имеют различный фенотип, хотя последовательности их ДНК в локусе, ответственном за проявление фенотипа, идентичны.
Примером эпигенетического эффекта является поведение белка, вызывающего заболевание коровьего бешенства. На рисунке ниже показано, что белок PrP может или находиться в простой растворимой форме, или принимать конформацию, при которой он образует крупные агрегаты. Образование этих агрегатов является причиной развития заболевания.
Они способны к самовоспроизведению, поскольку под их воздействием вновь синтезированные молекулы PrP принимают конформацию, способствующую агрегации. Аналогичные эффекты обнаружены в дрожжах, и, таким образом, эпигенетическое наследование является достаточно широко распространенной формой передачи наследственной информации. Во всех этих случаях фенотип определяется скорее наличием предсуществующих белковых агрегатов, а не последовательностью гена, кодирующего данный белок.
Мы не знаем, нужна ли клетке информация, заложенная в форме, отличной от последовательности нуклеотидов в ДНК Если бы смогли считать последовательности ДНК, кодирующие все белки, оказались бы они способны к взаимодействию с образованием всех клеточных структур и функций? Если нет, то какова природа информации, которая позволяет клетке образоваться только из уже существующей клетки?
Необходимы ли некие предсуществующие матрицы, на которых в дальнейшем собираются все структуры клетки (Важным фактором является локализация клеточных структур)?
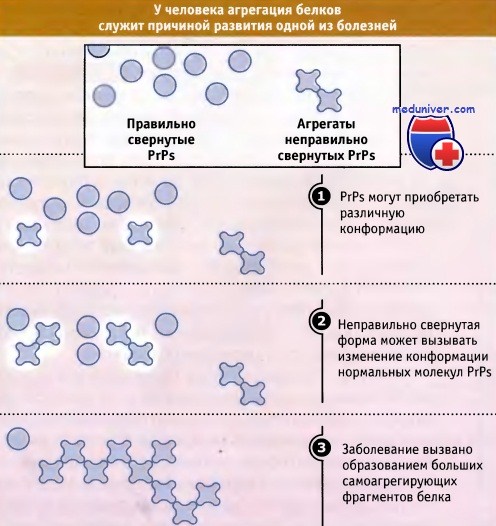
PrP существуют в растворимой форме или приобретают альтернативную конформацию, способную к агрегации.
Под влиянием агрегатов вновь синтезированнные молекулы PrP начинают приобретать конформацию, благоприятствующую агрегации.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Вернуться в содержание раздела "генетика" на нашем сайте
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
БАКТЕРИИ. Химический состав клеток бактерий в основном такой же, как и у клеток высокоорганизованных организмов. Они окружены оболочкой, внутри которой находится цитоплазма, ядерный аппарат, рибосомы, ферменты и другие включения. В отличие от клеток эукариот в клетках бактерий отсутствуют митохондрии, аппарат Гольджи, эндоплазматическая сеть. Цитоплазма бактерий представляет собой коллоидную часть клетки с гранулярной (зернистой) структурой. Основную массу гранул составляют рибосомы с константой седиментации 70S, участвующие в синтезе белка. В центральной части цитоплазмы бактерий расположен ядерный аппарат - нуклеотид и плазмиды. Ядро прокариот называется нуклеотидом по той причине, что оно в отличии от эукариот не изолировано от цитоплазмы мембраной и представлено одной очень длинной молекулой ДНК (хромосомой). Хромосома бактерии E. coli включает около 5*10 6 пар оснований, имеет имеет молекулярную массу 3*10 9 Д.В хромосоме кишечной палочки ДНК замкнута в кольцо и состоит из дискретно расположенных генов. Длина молекулы ДНК в расправленном состоянии достигает 1 мм, что значительно превышает среднюю длину самой бактерии.
ДНК бактерий не отличается по строению от ДНК высших организмов. Она содержит те же четыре нуклеотида, в состав каждого из которых входят дезоксирибоза, остаток фосфорной кислоты и пуриновое (аденин или гуанин) или пиримидиновое (тимин или цитозин) основание. Соединение мононуклеотидных остатков осуществляется сложноэфирными мостиками.
ВИРУСЫ. Вирусная частица содержит в своём составе одну из нуклеиновых кислот, которая окружена белковой оболочкой (капсидом). Геном вирусов может быть представлен двухцепочной или одноцепочной ДНК, одноцепочной или двухцепочной РНК. Типичная частица состоит из головки и хвостового отростка. В головке плотно упакована ДНК. Головку и хвостовой отросток покрывает белковый чехол. На конце хвостового отростка имеются специальные волоконца, облегчающие прикрепление фага к оболочке бактерий.
7. Внехромосомные факторы наследования.
Наследование признаков через цитоплазму называется внехромосомной. Цитоплазма влияет на активность проявления гена. Некоторые её органоиды, имеющие свою систему белкового синтеза (митохондрии, пластиды), могут влиять на развитие определённых признаков. В процессе развития наблюдается сложное взаимодействие ядра и цитоплазмы. Но определяющую роль всё-таки играет ядро.
8. Роль плазмид в определении у бактерий свойств устойчивости к антибиотическим лекарственным веществам.
У некоторых непатогенных и патогенных видов бактерий обнаружены факторы резистентности к лекарственным веществам – R-факторы. Это довольно крупные плазмиды. Они, присутствуя в цитоплазме бактерий и передаваясь их поколения в поколение, обуславливают устойчивость бактерий к действию одного или нескольких лекарственных веществ.
9. Способы передачи наследственной информации у микроорганизмов: трансформация, трансдукция и конъюгация.
В процессе трансформации принимают участие две бактериальные клетки – донор и реципиент. Трансформирующий агент представляет собой часть молекулы ДНК донора, которая внедряется в геном реципиента, изменяя его фенотип. В процессе трансформации клетки донора и реципиента не соприкасаются друг с другом. Механизм переноса генетического материала заключается в том, что из клеток донора выделяются в окружающую среду молекулы или фрагменты молекул ДНК. Сначала ДНК адсорбируется на оболочке клетки реципиента. Затем через определённые рецепторные участки её стенки при помощи специальных клеточных белков ДНК втягивается внутрь клетки. Проникающая донорская ДНК должна быть двухцепочной. В реципиентной клетке она становиться одноцепочной.
Трансдукция – это перенос генов из одной бактериальной клетки в другую при помощи умеренных фагов. При переносе генетического материала происходит замена участка молекулы ДНК фага. Фаг при это теряет свой собственный фрагмент и становиться дефектным. Различают три вида трансдукции: общую, или неспецифическую, специфическую и абортивную.
Конъюгация – это передача генетического материала от одних бактерий другими при их скрещивании.
Читайте также: