
Особенности структуры рнк и днк вирусного происхождения
Обновлено: 26.04.2024
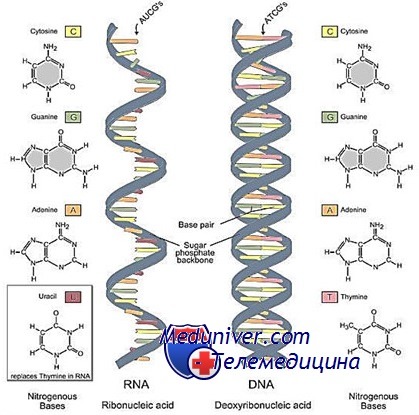
Структура и типы РНК человека: транспортная, ядерная, рибосомальная, митохондриальная и т.д.
Гетерогенная ядерная и информационная РНК
ГяРНК и её производное — информационная (или матричная) РНК переносят генетическую информацию от ядерной ДНК к цитоплазме.
Количество видов гяРНК равно количеству генов, так как она служит прямой копией кодирующих последовательностей генома. В процессе транскрипции РНК с ДНК ключевую роль играет фермент РНК-полимераза II. Информационная РНК образуется в результате процессинга гяРНК, при котором происходят вырезание некодирующих участков (интронов) и склеивание кодирующих экзонов. Таким образом, в состав иРНК входят кодирующая информация соответствующих видов гяРНК, а также фланкирующий лидерный и трейлерный участки, по этой причине она значительно короче.
Транспортная РНК
Так, например, тРНК, антикодон которой имеет последовательность 5'-ЦЦА-3', может нести только аминокислоту триптофан. Следует отметить, что данная зависимость лежит в основе передачи генетической информации, носителем которой выступает тРНК.
Транскрипция молекул тРНК происходит с кодирующих её последовательностей в ДНК при участии фермента РНК-полимеразы III. Различают более 40 семейств тРНК, которые, в свою очередь, подразделяют на несколько видов.
Рибосомальная РНК
Существует несколько субъединиц рРНК, которые различаются по коэффициенту седиментации (осаждения), измеряемому в единицах Сведберга (S). Данный коэффициент зависит от скорости осаждения субъединиц при центрифугировании в насыщенной водной среде.
Траскрипция рРНК с ДНК происходит при помощи двух дополнительных РНК-полимераз. РНК-полимераза I транскрибирует 5S, 5,8S и 28S в виде одного длинного 45S-тpaнскрипта, который затем разделяется на необходимые части. Таким образом обеспечивается равное количество молекул. В организме человека в каждом гаплоидном геноме присутствует примерно 250 копий последовательности ДНК, кодирующей 45S-транскрипт. Они расположены в пяти кластерных тандемных повторах в коротких плечах хромосом 13, 14, 15, 21 и 22.
Данные участки известны как ядрышковые организаторы, так как их транскрипция и последующий процессинг 45S-транскрипта происходят внутри ядрышка.
Не менее чем в трёх кластерах хромосомы 1 существует 2000 копий 5S-pPHK гена. Их транскрипция протекает в присутствии РНК-полимеразы III снаружи ядрышка. Затем они доставляются к местам сборки рибосом при помощи рибосомальных белков.
В рРНК насчитывают около 95 псевдоуридиновых участков, образованных посредством изомеризации уридина малой ядрышковой РНК.
Малая ядерная РНК. Превращение гяРНК в иРНК путём удаления интронов проходит в ядерном комплексе РНК-белков, называемом сплайсомой. У каждой сплайсомы есть ядро, состоящее из трёх малых (низкомолекулярных) ядерных рибонуклео-протеинов, или снурпов. Каждый снурп содержит хотя бы одну малую ядерную РНК и несколько белков. Существует несколько сотен различных малых ядерных РНК, транскрибируемых в основном РНК-полимеразой II.
Считают, что их основная функция — распознавание специфических рибонуклеиновых последовательностей посредством спаривания оснований по типу РНК—РНК. Для процессинга гяРНК наиболее важны Ul, U2, U4/U6 и U5.
Малая ядрышковая РНК. Малая (низкомолекулярная) ядрышковая РНК в основном участвует в направлении или проведении модификаций оснований в рРНК и малой ядерной РНК, таких, как, например, метилирование и псевдоуридинизация. Большинство малых ядрышковых РНК находятся в интронах других генов.
Сигналраспознающая РНК. Сигналраспознающая РНК распознаёт сигнальную последовательность белков, предназначенных для экспрессии, и участвует в их переносе через цитоплазматическую мембрану.
Митохондриальная РНК
Митохондриальная ДНК представляет собой непрерывную петлю и кодирует 13 полипептидов, 22 тРНК и 2 рРНК (16S и 23S). Большинство генов находятся на одной (тяжёлой) цепи, однако некоторое их количество расположено и на комплементарной ей лёгкой. При этом обе цепи транскрибируются в виде непрерывных транскриптов при помощи митохондриоспецифической РНК-полимеразы. Данный фермент кодируется ядерным геном. Длинные молекулы РНК затем расщепляются на 37 отдельных видов, а мРНК, рРНК и тРНК совместно транслируют 13 мРНК. Большое количество дополнительных белков, которые поступают в митохондрию из цитоплазмы, транслируются с ядерных генов.
У пациентов с системной красной волчанкой обнаруживают антитела к снурп-белкам собственного организма. Кроме того, считают, что определённый набор генов малой ядерной РНК хромосомы 15q играет важную роль в патогенезе синдрома Прадера—Вилли (наследственное сочетание олигофрении, низкого роста, ожирения, гипотонии мышц).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Типы вирусных геномов. Вирусная нуклеиновая кислота представлена только одной нуклеиновой кислотой (ДНК или РНК). Каждая из них является геномом. В разных по величине вирионах в геноме насчитывают от нескольких до многих десятков генов. Геномные нуклеиновые кислоты вирусов отличаются большим разнообразием структуры и формы.
Геном вируса является гаплоидным [греч. Haploos, одиночный и eidos, вид], т. е. представлен одним набором генов. Частично диплоидны [греч. Diploos, двойной] ДНК-содержащие вирусы, в ДНК которых встречаются повторяющиеся нуклеотидные последовательности. Полностью диплоидны ретровирусы, геном которых представлен двумя идентичными молекулами РНК.
По форме молекула ДНК может быть: 1) линейной, 2) кольцевой (циркулярно-замкнутой), 3) ковалентно-сцепленные суперспирализованные (например, у паповавирусов).
В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развернутые на 180°) повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо.
Молекулярная масса вирусных ДНК в 10–100 раз меньше массы бактериальных ДНК.
Особенности строения вирусной РНК.Вирусные РНК различаются в еще большей степени. Молекулы РНК по структуре могут быть:
1) одно- и двухцепочечные (диплоидный геном); 2) цельные (сплошные) и фрагментированные (сегментированные) на 2–3 . 8–12 сегментов.
Наличие сегментов ведет к увеличению кодирующей ёмкости генома.
По форме РНК различают: 1) линейные, 2) кольцевые.
Среди РНК-геномных вирусов с одноцепочечной линейной молекулой РНК различают вирусы с +РНК (позитивным) и -РНК (негативным) геномом.
-РНК не способны транслировать генетическую информацию непосредственно на рибосомах, то есть они не могут функционировать как иРНК. Синтез иРНК у РНК-негативных вирусов осуществляется в зараженной клетке на матрице -РНК с помощью вирусоспецифического фермента транскриптазы.
Структурно-функциональная организация вирусного генома. ДНК-содержащие вирусы также, как прокариоты и эукариоты, имеют структурные гены, кодирующие белки-ферменты, и регуляторные гены, детерминирующие образование репрессоров, подавляющих, в частности, функцию структурных.
Считывание информациисоперонов контролируется энхансером [англ. enhancer] или усилителем транскрипции; промотором [лат. promotum, продвигать], ответственным за ее инициацию (начало), с которым связывается фермент РНК-полимераза, осуществляющая транскрипцию ДНК; оператором[от лат. работник], регулирующим транскрипцию оперона (или отдельных генов) и терминатором [лат. terminare, ограничивать], прекращающим ее. При этом регуляторные участки оперона представляют собой короткие последовательности нуклеотидов ДНК; энхансер, промотор и оператор расположены в его начале (перед структурными генами), а терминатор – в конце.
В структурных генах вирусных оперонов, как и в клетках эукариот, имеются кодируемые участки нуклеотидных последовательностей, несущих информацию (экзоны), и некодируемые вставочные последовательности (интроны), которые после транскрипции в процессе созревания (процессинга) иРНК вырезаются с одновременным считыванием экзонов, что называется сплайсингом [англ. splice, соединять, сращивать].
Кодирующая способность вирусного генома.Число генов в вирусных геномах колеблется от 3–4 у самых простых вирусов до многих десятков у сложно устроенных. Увеличение генетической информации при минимальном содержании генетического материала происходит за счет того, что:
1) вирусные иРНК в отличие от иРНК про- и эукариот могут направлять синтез не одного, а двух-трех белков. Достигается это двухкратным считыванием одной и той же иРНК с находящихся в ней в разных участках двух-трех инициирующих АУГ-кодонов. Образующиеся полипептиды с разных инициирующих кодонов будут копиями, отличающимися только длиной;

2) при сдвиге рамки считывания на один или два нуклеотида и появлении нового генетического кода молекула иРНК может транслироваться с образованием таких полипептидов, у которых нет идентичных аминокислотных последовательностей. Такие белки называют уникальными белками;
3) нередко у вирусов происходит трансляция гигантских полипептидов-предшественников с последующим нарезанием их на более мелкие;
4) относительно невысокий уровень генетической информации вирусов компенсируется исключительно точным механизмом переключения с репликации на транскрипцию и наоборот, что особенно ярко проявляется при репродукции РНК-содержащих вирусов.
Из нескольких сотен известных в настоящее время вирусов человека и животных РНК-геном содержит около 80% вирусов. Способность РНК хранить наследственную информацию является уникальной особенностью вируса.
У просто организованных и некоторых сложно организованных вирусов вирусная РНК в отсутствие белка может вызвать инфекционный процесс. Впервые инфекционная активность РНК вируса табачной мозаики была продемонстрирована X. Френкель-Конратом и соавт. в 1957 г. и А. Гирером и Г. Шраммом в 1958 г. Впоследствии положение об инфекционной активности РНК было перенесено на все РНК-содержащие вирусы, однако долголетние усилия доказать это для таких вирусов, как вирусы гриппа, парамиксовирусы, рабдовирусы (так называемые минус-нитевые вирусы), оказались бесплодными: у этих вирусов инфекционной структурой являются не РНК, а комплекс РНК с внутренними белками. Таким образом, геномная РНК может обладать инфекционной активностью в зависимости от своей структуры.
Структура вирусных РНК чрезвычайно разнообразна. У вирусов обнаружены однонитчатые и двунитчатые, линейные, фрагментированные и кольцевые РНК. РНК-геном в основном является гаплоидным, геном ретровирусов — диплоидный, т. е. состоит из двух идентичных молекул РНК.
В основном однонитчатые РНК являются линейными молекулами, однако РНК-фрагменты буньявирусов обнаружены в виде кольцевой формы. Кольцевая форма возникает за счет образования водородных связей между концами молекул.
Двунитчатые РНК.Этот необычный для клетки тип нуклеиновой кислоты, впервые обнаруженный у реовирусов, широко распространен среди вирусов животных, растений и бактерий. Вирусы, содержащие подобный геном, называют диплорнавирусы.
Общей особенностью диплорнавирусов является фрагментированное состояние генома. Так, геном реовирусов состоит из 10 фрагментов, ротавирусов — из 11 фрагментов.
Молекулярная масса РНК таких вирусов варьирует в широких пределах.
БЕЛКИ
В зараженной клетке вирусный геном кодирует синтез двух групп белков: 1) структурных, которые входят в состав вирусных частиц потомства, и 2) неструктурных, которые обслуживают процесс внутриклеточной репродукции вируса на разных его этапах, но в состав вирусных частиц не входят.
Структурные белки делятся на 2 группы:
1) капсидные белки, образующие капсид, т. е. футляр для нуклеиновой кислоты вируса (от лат. capsa — вместилище), и входящие в состав капсида геномные белки, и ферменты;
2) суперкапсидные белки, входящие в состав суперкапсида, т. е. наружной вирусной оболочки.
Просто организованные вирусы, представляющие собой нуклеокапсид, содержат только капсидные белки. Сложно организованные вирусы содержат капсидные и суперкапслдные белки.
В составе капсида некоторых вирусов (пикорнавирусы, паповавирусы, аденовирусы) содержатся белки, ковалентно связанные с вирусным геномом (геномные белки). Эти белки являются терминальными, т. е. соединенными с концом вирусной нуклеиновой кислоты. Функции их неразрывно связаны с функциями генома и их регуляцией.
У ряда сложно организованных вирусов в составе капсида имеются ферменты, осуществляющие транскрипцию и репликацию вирусного генома — РНК и ДНК (РНК-и ДНК-полимеразы), а также ферменты, модифицирующие концы иРНК. Если ферменты и геномные белки представлены единичными молекулами, то капсидные белки представлены множественными молекулами. Эти белки и формируют капсидную оболочку, в которую у сложно организованных вирусов вставлены молекулы белков с другими функциями.
Основным принципом строения капсидной оболочки вирусов является принцип субьединичности, т. е. построение капсидной оболочки из субъединиц-капсомеров, образованных идентичными полипептидными цепями. Правильно построенные белковые субъединицы — капсомеры возникают благодаря способности вирусных капсидных белков к самосборке. Самосборка объясняется тем, что упорядоченная структура — капсид имеет наименьшую свободную энергию по сравнению с неупорядоченными белковыми молекулами. Сборка капсидной оболочки из субъединиц запрограммирована в первичной структуре белка и происходит самопроизвольно или при взаимодействии с нуклеиновой кислотой.
Принцип субъединичности в строении вирусного капсида является универсальным свойством капсидных белков и имеет огромное значение для вирусов. Благодаря этому свойству достигается огромная экономия генетического материала. Если бы капсидная оболочка была построена из разных белков, то на кодирование ее потребовалась бы основная часть генетической информации, заложенной в вирусном геноме. В действительности на кодирование, например, одной полипептидной цепи вируса табачной мозаики, расходуется менее 10% генома. Далее, в механизме самосборки заложена возможность контроля за полноценностью вирусных полипептидов: дефектные и чужеродные полипептидные цепи при таком способе сборки вирионов будут автоматически отбрасываться.
Описанная способность к самосборке в пробирке и в зараженной клетке характерна только для простых вирусов. Сборка сложно организованных вирусов является гораздо более сложным многоступенчатым процессом, хотя отдельные ее этапы, например формирование капсидов и нуклеокапсидов, также основаны на самосборке.
Суперкапсидные белки. Гликопротеиды. Суперкапсидные белки, или пепломеры, располагаются в липопротеидной оболочке (суперкапсиде или пеплосе) сложно устроенных вирусов. Они либо пронизывают насквозь липидный бислой как, например, гликопротеиды альфа-вирусов (вируса леса Семлики), либо не доходят до внутренней поверхности. Эти белки являются типичными внутримембранными белками и имеют много общего с клеточными мембранными белками. Как и последние, суперкапсидные белки обычно гликозилированы. Углеводные цепочки прикреплены к молекуле полипептида в определенных участках. Гликозилирование осуществляют клеточные ферменты, поэтому один и тот же вирус, продуцируемый разными видами клеток, может иметь разные у углеводные остатки: может варьировать как состав углеводов, так и длина углеводной цепочки и место прикрепления ее к полипептидному остову.
Основной функцией гликопротеидов является взаимодействие со специфическими рецепторами клеточной поверхности. Благодаря этим белкам осуществляется распознавание специфических клеточных рецепторов и прикрепление к ним вирусной частицы, т. е. адсорбция вируса на клетке. Поэтому гликопротеиды, выполняющие эту функцию, называют вирусными прикрепительными белками.
Другой функцией гликопротеидов является участие в слиянии вирусной и клеточной мембран, т. е. в событии, ведущем к проникновению вирусных частиц в клетку. Вирусные белки слияния ответственны за такие процессы, как гемолиз и слияние плазматических мембран соседних клеток, приводящие к образованию гигантских клеток, синцитиев и симпластов.
Неструктурные белки. Неструктурные белки изучены гораздо хуже, чем структурные, поскольку их выделяют не из очищенных препаратов вирусов, а из зараженных клеток, и возникают трудности в их идентификации и очистке от клеточных белков.
К неструктурным белкам относятся:
1) предшественники вирусных белков, которые отличаются от других неструктурных белков нестабильностью в зараженной клетке в результате быстрого нарезания на структурные белки;
2) ферменты синтеза РНК и ДНК (РНК- и ДНК- полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома;
4) ферменты, модифицирующие вирусные белки, например протеиназы и протеинкиназы.
Однако многие неструктурные белки при ряде вирусных инфекций еще не идентифицированы и функции их не определены.
Строение вирусов. Классификация вирусов
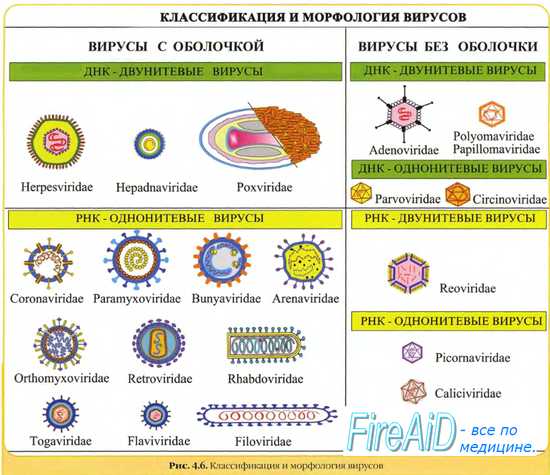
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
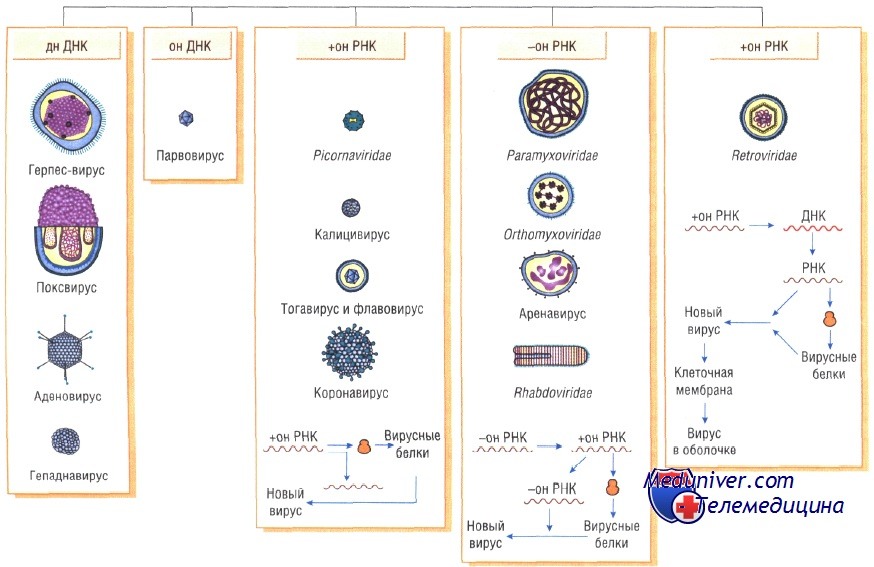
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
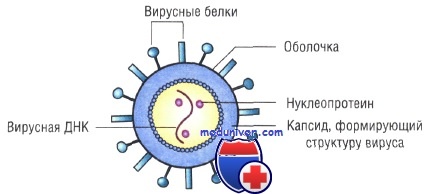
Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных.
Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
Морфология вирусов. Размеры вирусов.
Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки.
Нуклеиновые кислоты вирусов
Вирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине.
Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180") повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
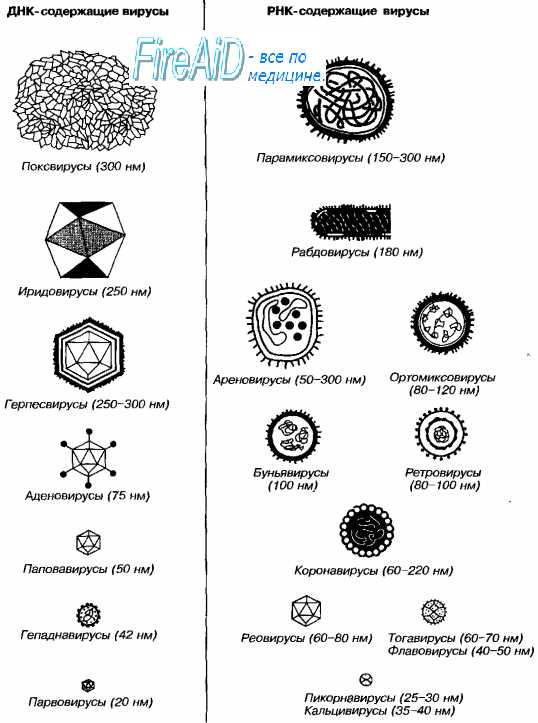
Рис. 2-1. Размеры и морфология основных возбудителей вирусных инфекций человека.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК.
Инфекционность нуклеиновых кислот вирусов
Многие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: