
Отличия вирусов от плазмид
Обновлено: 24.04.2024
• В клетках большинства прокариот присутствует одна кольцевая хромосома
• Генетическая гибкость прокариот и их адаптационные возможности усиливаются при заражении бактериофагами и при переносе плазмид
• Быстрая эволюция генома прокариот обеспечивается транспозонами и другими мобильными элементами
У большинства прокариот присутствует одна кольцевая хромосома, и они являются гаплоидными клетками. В следующем разделе рассмотрены вопросы организации хромосомы в дискретную структуру, нуклеоид. В настоящее время мы располагаем данными секвенирования более 200 геномов прокариот, размеры которых варьируют от 580 кнп (Mycoplasma genitalium) до 9 Мнп (Streptomyces, Myxococcus).
Геном таких хорошо известных бактерий, как Е. coli и В. subtilis, занимает среднее положение (4-5 Мнп). Относительно небольшие размеры прокариотических хромосом, по сравнению с хромосомами высших эукариот, объясняются их компактностью и небольшим количеством некодирующих последовательностей ДНК. В общем, гены, необходимые для постоянного роста и поддержания жизнеспособности прокариот, находятся в хромосоме, а генетическая гибкость микроорганизмов обеспечивается различными мобильными элементами.
У некоторых бактерий хромосома имеет линейную форму, или же в клетке содержится несколько таких хромосом. Встречаются виды, для которых характерны обе эти особенности. Например, клетки Streptomyces содержат линейную хромосому. Концы этой хромосомы замкнуты белковым мостиком, и это объясняет, почему долгое время на генетической карте этих бактерий изображали круговую хромосому.
Бактерии Rhodobacter sphaeroides обладают двумя большими кольцевыми хромосомами (3,0 и 0,9 Мнп), в каждой из которых присутствует много необходимых генов домашнего хояйства. Возбудитель Лайм-боррелиоза, бактерия Воrrelia burgdorferi характеризуется наличием нескольких линейных хромосом.
Стабильные элементы внехромосомной ДНК, которые не несут необходимых генов домашнего хозяйства и поэтому не являются существенными, называются плазмиды. На рисунке ниже перечислены некоторые хорошо изученные плазмиды бактерий. Эти данные дают представление о размерах генов, которые могут нести эти плазмиды. Плазмиды обычно невелики, варьируя по размерам от 2 до 1000 кнп, и обладают кольцевой структурой.
Так же как и в случае хромосом, известны исключения, и некоторые крупные плазмиды достигают размера 1 Мнп или больше, и небольшая их часть обладает линейной структурой. Все плазмиды несут гены, управляющие их репликацией, обычно включающей различные элементы аппарата репликации клетки хозяина. К числу важнейших генов, которые несут плазмиды, относятся гены устойчивости к антибиотикам и обусловливающие патогенные свойства или деградацию необычных источников углерода.
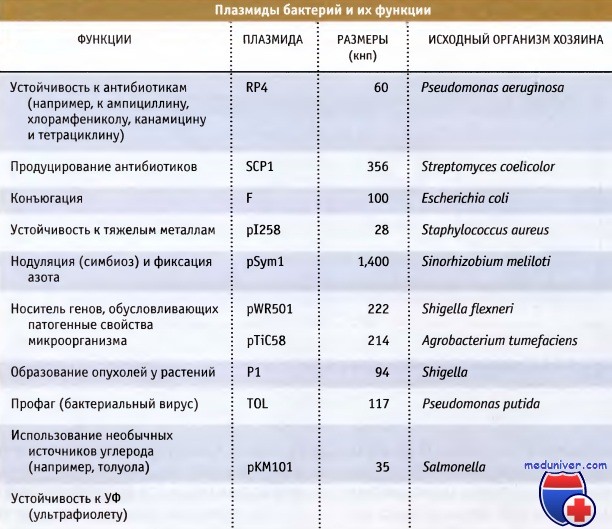
Плазмиды бактерий и их функции.
Плазмиды могут распространяться между организмами различными путями. Конъюгация представляет собой процесс, напоминающий спаривание, при котором присходит прямой перенос ДНК от клетки донора, содержащей плазмиду, в клетку реципиента. В донорской плазмиде закодированы функции, необходимые дляУ большинства прокариот присутствует одна кольцевая хромосома, и они являются гаплоидными клетками. В следующем разделе рассмотрены вопросы организации хромосомы в дискретную структуру, нуклеоид.
В настоящее время мы располагаем данными секвенирования более 200 геномов прокариот, размеры которых варьируют от 580 кнп (Mycoplasma genitalium) до 9 Мнп (Streptomyces, Myxococcus). Геном таких хорошо известных бактерий, как Е. coli и В. subtilis, занимает среднее положение (4-5 Мнп). Относительно небольшие размеры прокариотических хромосом, по сравнению с хромосомами высших эукариот, объясняются их компактностью и небольшим количеством некодирующих последовательностей ДНК. В общем, гены, необходимые для постоянного роста и поддержания жизнеспособности прокариот, находятся в хромосоме, а генетическая гибкость микроорганизмов обеспечивается различными мобильными элементами.
У некоторых бактерий хромосома имеет линейную форму, или же в клетке содержится несколько таких хромосом. Встречаются виды, для которых характерны обе эти особенности. Например, клетки Streptomyces содержат линейную хромосому. Концы этой хромосомы замкнуты белковым мостиком, и это объясняет, почему долгое время на генетической карте этих бактерий изображали круговую хромосому.
Бактерии Rhodobacter sphaeroides обладают двумя большими кольцевыми хромосомами (3,0 и 0,9 Мнп), в каждой из которых присутствует много необходимых генов домашнего хояйства. Возбудитель Лайм-боррелиоза, бактерия Воrrelia burgdorferi характеризуется наличием нескольких линейных хромосом.
Стабильные элементы внехромосомной ДНК, которые не несут необходимых генов домашнего хозяйства и поэтому не являются существенными, называются плазмиды. На рисунке ниже перечислены некоторые хорошо изученные плазмиды бактерий. Эти данные дают представление о размерах генов, которые могут нести эти плазмиды. Плазмиды обычно невелики, варьируя по размерам от 2 до 1000 кнп, и обладают кольцевой структурой.
Так же как и в случае хромосом, известны исключения, и некоторые крупные плазмиды достигают размера 1 Мнп или больше, и небольшая их часть обладает линейной структурой. Все плазмиды несут гены, управляющие их репликацией, обычно включающей различные элементы аппарата репликации клетки хозяина. К числу важнейших генов, которые несут плазмиды, относятся гены устойчивости к антибиотикам и обусловливающие патогенные свойства или деградацию необычных источников углерода.
Плазмиды могут распространяться между организмами различными путями. Конъюгация представляет собой процесс, напоминающий спаривание, при котором присходит прямой перенос ДНК от клетки донора, содержащей плазмиду, в клетку реципиента. В донорской плазмиде закодированы функции, необходимые для обеспечения контакта с клеткой реципиентом, инициации переноса реплицированной ДНК, и самого переноса ДНК в клетку реципиента.
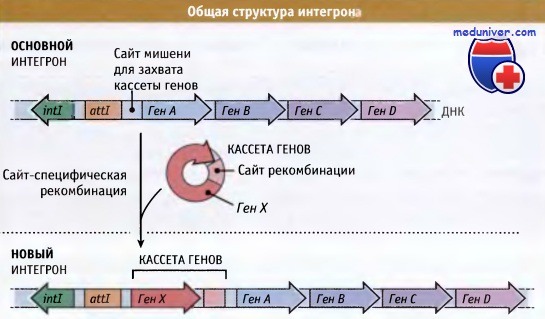
Структура типичного интегрона после накопления нескольких кассет генов.
Плазмиды также могут передаваться при прямом захвате ДНК (трансформация) или с участием бактериофага (трансдукция). Независимо от наличия экстрахромосомных элементов, изменения в бактериальном геноме могут возникать за счет процессов гомологичной и сайт-специфической рекомбинации.
Для многих бактерий важными источниками генетической вариабельности, по-видимому, являются бактериофаги (бактериальные вирусы). Результаты секвенирования показывают, что в геноме многих бактерий присутствуют интегрированные последовательности бактериофага (профаги). Например, у Е. coli их по меньшей мере 9, а у B. subtilis 10. Некоторые из этих профагов обладают дефектами, например у них присутствуют делеции и другие мутации, что делает маловероятным их дальнейшую активацию с образованием инфекционных бактериофагов.
В некоторых случаях профаги несут гены, которые обеспечивают некоторые преимущества клеткам хозяина: это относится к системам рестрикции и модификации, устойчивости к УФ и к таким детерминантом патогенности, как токсины.
Наконец, бактериальный геном также содержит много мобильных генетических элементов, которые распространяются посредством транспозиции. Инсерционные последовательности содержат минимальные элементы, которые, в своей простейшей форме, вместе с фланкирующими последовательностями, обладают только одним геном транспозазы. При инициации транспозиции, фланкирующие последовательности узнаются белком транспозазой.
Затем происходит мобилизация ферментов клетки хозяина, участвующих в репликации и репарации ДНК. Они завершают процесс вставки последовательности в необходимом месте. Более сложные транспозоны несут дополнительные гены, способные обеспечить адаптационные преимущества клеткам хозяина. Наиболее известными примерами транспозонов у бактерий являются гены устойчивости к антибиотикам, однако подобным образом могут переноситься также многие другие гены. Близкие к транспозонам элементы могут катализировать несколько таких типов перегруппировок ДНК, как инверсии и делеции.
Интегроны представляют собой особенно важный инструмент адаптивной перегруппировки генома. Как показано на рисунке ниже, интегроны обычно состоят из гена интегразы, примыкающего к нему сайта мишени для захвата кассеты генов, и сильного промотора, регулирующего экспрессию захваченных генов. В кассетах, часто включающих гены устойчивости к антибиотикам, содержатся последовательности, которые позволяют им включаться на место мишени под действием белка интегразы. Интегроны могут расти за счет последовательного захвата различных кассет генов, что способствует быстрому развитию таких бактерий, которые устойчивы к ряду антибиотиков.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Обзор
Многообразие мобильных генетических элементов
Автор
Редакторы

Организмы и геномы можно таким образом расценивать как секции биосферы, по которым гены глобально циркулируют с различной интенсивностью, и в которые персональные гены и опероны могут включаться, если они предоставляют достаточные преимущества.
K. Jeon и J. Danielli [1]

Основные сокращения
МГЭ мобильные генетические элементы ГГП горизонтальный генетический перенос ОРС открытая рамка считывания, последовательность нуклеотидов между инициирующим и терминирующим кодонами гена ori T место начала переноса цепи плазмидной ДНК при мобилизации ori V место начала репликации (копирования, воспроизводства) плазмидной ДНК rep ген белка-инициатора репликации многих бактериальных плазмид
Вирусы: суперпаразиты и помощники
В 2012 году охарактеризовали интегрированную в ДНК мимивирусов форму вирофага и даже новый класс МГЭ — трансповироны, способные встраиваться в ДНК и мимимирусов, и вирофагов [7]. Подобно бактериофагам, переносящим гены от одних бактерий к другим, вирофаги могут играть важную роль в ГГП между разными группами вирусов и их хозяевами.
Многие профаги кодируют факторы вирулентности, трансформируя нетоксигенные бактерии в агентов-убийц: профаг CTXphi из Vibrio cholerae кодирует холерный токсин, а упомянутый выше P22 — ферменты конверсии О-антигена сальмонеллы, позволяя ей уходить от иммунного надзора.
В составе фагов обнаруживают другие МГЭ (например, транспозоны, несущие гены антибиотикорезистентности) и их модули (системы репликации и переноса конъюгативных плазмид). Самый известный пример слияния модулей фагового и плазмидного происхождения — фаг Р1, способный реплицироваться и длительно поддерживаться вне хромосомы, ничем не отличаясь от плазмиды. Профаги разных бактерий детально рассмотрены в обзоре [8].
Плазмиды — маленькие гиганты больших процессов
Плазмиды — внехромосомные двуцепочечные молекулы ДНК, способные к длительному автономному существованию в клетках прокариот и некоторых эукариот. Чаще всего плазмидные ДНК суперскручены и ковалентно замкнуты в кольцо, однако у актиномицетов и спирохет встречаются и линейные формы, что обычно сочетается с линейной организацией хромосом. Размер плазмид обычно варьирует от 0,85 т.п.н. (pRKU1 из Thermotoga petrophila) до 600 т.п.н., но у бруцелл и ризобий описаны мегаплазмиды размером более 1 млн п.н., что иногда делает вопрос их дифференцировки от дополнительных хромосом риторическим [9]. Элиминация мегаплазмид, в отличие от хромосом, обычно не вызывает фатальных для бактериальной клетки последствий, однако вместе с плазмидами могут утратиться такие важные функции, как способность к фиксации азота и формированию симбиотических клубеньков (у ризобий).
На долю плазмидной ДНК может приходиться 1–15% наследственной информации бактериальной клетки, однако известны случаи, когда плазмидами контролируется до 25% (у некоторых Archaea) и даже до 40% информации (2 мегаплазмиды размером около 1,4 и 1,6 млн п.н. у Sinorhizobium meliloti).
Плазмидами часто мобилизуются сосуществующие с ними в одной клетке неконъюгативные МГЭ (но обладающие как минимум сайтом начала переноса oriT, а чаще и генами mob) и даже хромосомы. Мобилизация генов хромосомы возможна в случае интеграции в неё конъюгативной плазмиды. Это возможно, когда в обеих молекулах присутствуют одинаковые инсерционные последовательности (IS-элементы), обеспечивающие гомологичную рекомбинацию (типичный пример — F-фактор E.coli). Однако интеграция может быть и RecA-независимой, если плазмида кодирует тирозиновую интегразу (плазмиды стрептомицетов pSE101, pSAM2 и псевдомонад pKLK106). В этом случае интеграция чаще происходит в гены тРНК, но в некоторых хозяевах эта закономерность нарушается [3].
Транспозоны — универсальные генетические челноки
Транспозоны — МГЭ, перемещающиеся как в пределах одной молекулы ДНК, так и между разными репликонами одного генома (конъюгативные транспозоны — и между геномами). Фланкированы инвертированными повторами, а в центральной части содержат гены, ответственные за перемещение. Транспозоны прокариот подразделяют на IS-элементы, Tn-элементы и Mu-подобные фаги [10].
Интегроны — природные системы клонирования и экспрессии
В процессе перемещения от одного интегрона к другому или от одного сайта в интегроне к другому сайту, генная кассета существует как автономная и неспособная к репликации двунитевая кольцевая молекула ДНК. Кассеты, захватываемые интегронами и суперинтегронами, могут содержать гены факторов патогенности, метаболических путей, детерминанты антибиотико- и дезинфектантоустойчивости или гены рестрикционных ферментов. Интеграза IntI катализирует сайт-специфическую рекомбинацию между сайтами attI и attC, в результате чего происходит интеграция или вырезание кассеты. Множество событий интеграции ведет к образованию мультикассетных рядов, в которых все кассеты фланкированы attC-сайтами. Известны хромосомные суперинтегроны, включающие до 179 генных кассет (у Vibrio cholerae), однако среди клинически значимых бактерий большинство интегронов содержит до 5–8 генных кассет. Наиболее эффективно экспрессируются кассеты, расположенные ближе к промотору, но изменение селективного давления может способствовать перестройкам в составе интегрона.

Рисунок 3. Интегрон — ДНК, улавливающая генные кассеты и распространяемая в составе более высокоорганизованных МГЭ. а — Структура интегрона класса 1. Pint — промотор интегразы, Pant — промотор кассет антибиотикорезистентности. Остальные элементы объяснены в тексте; б — Иерархическая организация МГЭ.
Все интегроны, несущие кассеты антибиотикорезистентности, разделяют на 5 классов на основании гомологии последовательностей кодируемых ими интеграз. Большинство интегронов антибиотикорезистентности относится к классу 1 (часто ассоциированы с Tn21-семейством). Они включают два концевых невариабельных региона, называемых константными последовательностями (constant sequences, CS), и высоковариабельный центральный участок. В одном конце интегрона (5’-CS), обычно находятся intI, attI и промотор, от которого экспрессируются гены кассеты. В другом конце (3’-CS), находится часть гена qacEΔ1, который, будучи интактным, несёт устойчивость к четвертичным аммониевым соединениям. За ним расположен ген sul, определяющий резистентность микроорганизма к сульфаниламидам, и 1–2 гена с неустановленной функцией — orf5 и иногда orf6 (рис. 3, а). Интегроны класса 2 ассоциированы с Tn7-семейством, классов 3 и 5 — с плазмидами, класса 4 — с конъюгативным геномным островом SXT Vibrio cholerae.

Рисунок 4. Схематическое изображение структуры геномного острова в составе бактериальной хромосомы. DR — прямые повторы ДНК хромосомы, фланкирующие ГО; IS — инсерционные элементы.
Геномные острова гонококков (GGI) кодируют систему секреции типа IV (T4SS), родственную плазмидным системам конъюгационного переноса. Посредством T4SS распространяется не только сам элемент и его продукты, но секретируется в окружающую среду и хромосома Neisseria gonorrhoeae, которая затем может трансформировать другие бактерии и участвовать в рекомбинационных событиях.
Главные нарушители границ и их мекка
Из обитателей отстойников, а что самое опасное — из бактерий вод, уже прошедших очистку, — в большом количестве выделяют плазмиды IncP-1-группы несовместимости (в системе классификации плазмид псевдомонад) [12]. Эти относительно небольшие молекулы дарят хозяевам массу селективных преимуществ (от множественной резистентности до биодеградации хлорорганики) и способны распространяться не только среди псевдомонад, но и практически всех грамотрицательных и некоторых грамположительных бактерий. Кроме того, они способны мобилизовать неконъюгативные R-плазмиды (например, IncP-4) к переносу в ещё более широкий спектр бактерий, а также в дрожжи и клеточные линии млекопитающих [17].
Биология плазмид в России и за рубежом. Что же можно возвести на крепком фундаменте?
Плазмиды. Производными вирусов большинство исследователей в настоящее время признают плазмиды. Тем более, что генно-инженерными манипуляциями можно сконструировать плазмиду из вируса и даже из фрагментов хромосом клетки. В отличие от вирионов плазмиды существуют в виде кольцевых, двунитчатых ДНК, и гены их кодируют разные белки. С эволюционной точки зрения представляют интерес плазмиды, кодирующие ферменты, разрушающие или модифицирующие антибиотики. Именно с ними бывает часто связана лекарственная устойчивость бактерий.
Вироиды. Ещё более своеобразными вирусоподобными структурами являются вироиды – возбудители заразных болезней растений, сходных с обычными их вирусными инфекциями. Эти возбудители представляют относительно небольшие (120000-140000 дальтон) молекулы РНК, не несущие генетического кода, способ воспроизводства которых до сих пор не известен.
Прионы. Это совсем необычные возбудители некоторых медленных инфекций, например болезнь овец скрепи. В их составе не выявлено нуклеиновых кислот, но определены белки, откуда и идёт их название, аналогичное вириону классических вирусов. Одним из возможных толкований природы этих агентов является предположение, что ген приона находится в хромосоме, а сам прион является и индуктором, и продуктом этого гена, и обладателем патологических свойств.
Что же объединяет вирусы и вирусоподобные агенты – плазмиды, вироиды и прионы? Их объединяет то, что все они являются не организмами, не клетками, а автономными генетическими структурами, в разной степени способными к репродукции с помощью либо собственных, либо клеточных, либо тех и других вместе систем синтеза макромолекул. Будучи неклеточными формами жизни, они, сопутствуют клеточным формам жизни, нуждаясь в них и влияя на них.
Вопросы для самоконтроля:
1. Назовите основные характерные черты вирусов?
2. Каковы особенности строения вирусов растений, животных и бактерий?
3. Какие существуют гипотезы происхождения вирусов?
4. Назовите этапы развития вирусов?
5. Какие виды инфекций вызывают вирусы?
6. Какие выделяют группы вирусов в зависимости от генетического материала, содержащегося в них?
7. Приведите примеры вирусных заболеваний растений, животных и человека?
8. Каковы особенности строения, развития вируса иммунного дефицита человека?
Плазмида или R-фактор (от англ. resistance – устойчивость) (экстрахромосомный генетический элемент) представляет собой репликон, который стабильно наследуется в экстрахромосомном состоянии.
Плазмиды имеют собственные гены, которые наделяют их специфическими наследственными признаками и способностью к размножению, должны быть отнесены к живым организмам.
Плазмиды – наипростейшие организмы, лишенные оболочки, собственных систем синтеза белка и мобилизации энергии и представляющие собой особый класс абсолютных внутриклеточных паразитов, наделяющих своих бактерий-хозяев полезными для них свойствами (А. И. Коротяев).
Плазмиды как и вирусы:
* не имеют собственной белоксинтезирующей системы
*Не имеют собственной системы мобилизации энергии;
*не способны к росту и бинарному делению
* размножаются путем воспроизведения себя из собственного генома (путем его саморепликации)
*Являются абсолютными внутриклеточными паразитами
В отличие от вирусов:
* Геном представлен только двунитевой ДНК
* Размножение только путем саморепликации
* Среда обитания - бактерии
* Наделяют хозяина важными свойствами, повышающими жизнеспособность
*F-плазмиды: Донорные функции
*R-плазмиды :Устойчивость к лекарственным препаратам
*Col-плазмиды: Синтез колицинов
*Ent-плазмиды: Синтез энтеротоксннов
*Hly-плазмиды: Синтез гемолизинов
*Биодеградативные плазмиды: Разрушение различных органических и неорганических соединений, в том числе содержащих тяжелые металлы
*Криптические плазмиды: ф-ции неизвестны
Все известные плазмиды представляют собой кольцевидные суперспирализованные молекулы двунитевой ДНК, размеры которых варьируют от 1,5 до 200 МД и более (от 1500 до 400 000 пар нуклеотидов). Чем больше молекулярная масса, тем больше и сложнее набор генов, тем многообразнее функции плазмид.
* Саморегулируемая репликация. Эта функция свойственна всем живым организмам. В составе плазмидных ДНК имеются фиксированная точка ori (точка начала репликации) и соответствующие гены, контролирующие репликацию.
* Явление поверхностного исключения. Этот механизм не позволяет проникнуть в клетку, уже содержащую плазмиду, другой родственной ей плазмиде. Поверхностное исключение обеспечивается синтезом под контролем генов плазмиды особых белков наружной мембраны, которые препятствуют установлению контакта этой клетки с клеткой, несущей такую же плазмиду, или подавляют конъюгативный метаболизм ДНК этой плазмиды
* Явление несовместимости. Суть его заключается в том, что две близкородственные плазмиды не могут стабильно сосуществовать в одной клетке, одна из них подвергается элиминации (удалению).
* Контроль числа копий плазмиды на хромосому клетки. Различают малокопийные (1-4 копии) и многокопийные плазмиды (12-38 копий). Наличие собственных генов позволяет плазмиде осуществлять репликацию независимо от клеточного цикла клетки-хозяина. Информация, необходимая для осуществления репликации плазмиды, обычно заключена в небольшой участок ее ДНК, получивший название основного, или базового репликона.
* Контроль стабильного сохранения плазмид в клетке-хозяине (контроль стабильного поддержания).
* Контроль равномерного распределения дочерних плазмид в дочерние бактериальные клетки. Распределение плазмид между дочерними клетками происходит нерандомически, т. е. не по принципу случайности, а существует генетический механизм контроля равномерного распределения (сегрегации) вновь синтезированных плазмид при клеточном делении. Носительство плазмид для клетки-хозяина становится генетически необходимым, благодаря этому обеспечивается существование плазмид как организмов.
* Способность к самопереносу (у конъюгативных плазмид).
* Способность к мобилизации на перенос (у неконъюгативных плазмид).
* Способность наделять клетку-хозяина дополнительными важными для нее биологическими свойствами, способствующими выживанию бактерий, а следовательно, и плазмид в природе.
Важные св-ва плазмид:
*фертильность (способность женского или мужского организма к участию в оплодотворении);
*устойчивость к тяжелым металлам (кадмий, ртуть);
*способность к продукции бактериоцинов, антибиотиков, токсинов;
*способность к утилизации необычных источников углерода (камфора, октан, октанол).
Плазмиды патогенных микробов контролируют синтез различных факторов патогенности и образование новых способностей у многих видов бактерий:
Плазмида или R-фактор (от англ. resistance – устойчивость) (экстрахромосомный генетический элемент) представляет собой репликон, который стабильно наследуется в экстрахромосомном состоянии.
Плазмиды имеют собственные гены, которые наделяют их специфическими наследственными признаками и способностью к размножению, должны быть отнесены к живым организмам.
Плазмиды – наипростейшие организмы, лишенные оболочки, собственных систем синтеза белка и мобилизации энергии и представляющие собой особый класс абсолютных внутриклеточных паразитов, наделяющих своих бактерий-хозяев полезными для них свойствами (А. И. Коротяев).
Плазмиды как и вирусы:
* не имеют собственной белоксинтезирующей системы
*Не имеют собственной системы мобилизации энергии;
*не способны к росту и бинарному делению
* размножаются путем воспроизведения себя из собственного генома (путем его саморепликации)
*Являются абсолютными внутриклеточными паразитами
В отличие от вирусов:
* Геном представлен только двунитевой ДНК
* Размножение только путем саморепликации
* Среда обитания - бактерии
* Наделяют хозяина важными свойствами, повышающими жизнеспособность
*F-плазмиды: Донорные функции
*R-плазмиды :Устойчивость к лекарственным препаратам
*Col-плазмиды: Синтез колицинов
*Ent-плазмиды: Синтез энтеротоксннов
*Hly-плазмиды: Синтез гемолизинов
*Биодеградативные плазмиды: Разрушение различных органических и неорганических соединений, в том числе содержащих тяжелые металлы
*Криптические плазмиды: ф-ции неизвестны
Все известные плазмиды представляют собой кольцевидные суперспирализованные молекулы двунитевой ДНК, размеры которых варьируют от 1,5 до 200 МД и более (от 1500 до 400 000 пар нуклеотидов). Чем больше молекулярная масса, тем больше и сложнее набор генов, тем многообразнее функции плазмид.
* Саморегулируемая репликация. Эта функция свойственна всем живым организмам. В составе плазмидных ДНК имеются фиксированная точка ori (точка начала репликации) и соответствующие гены, контролирующие репликацию.
* Явление поверхностного исключения. Этот механизм не позволяет проникнуть в клетку, уже содержащую плазмиду, другой родственной ей плазмиде. Поверхностное исключение обеспечивается синтезом под контролем генов плазмиды особых белков наружной мембраны, которые препятствуют установлению контакта этой клетки с клеткой, несущей такую же плазмиду, или подавляют конъюгативный метаболизм ДНК этой плазмиды
* Явление несовместимости. Суть его заключается в том, что две близкородственные плазмиды не могут стабильно сосуществовать в одной клетке, одна из них подвергается элиминации (удалению).
* Контроль числа копий плазмиды на хромосому клетки. Различают малокопийные (1-4 копии) и многокопийные плазмиды (12-38 копий). Наличие собственных генов позволяет плазмиде осуществлять репликацию независимо от клеточного цикла клетки-хозяина. Информация, необходимая для осуществления репликации плазмиды, обычно заключена в небольшой участок ее ДНК, получивший название основного, или базового репликона.
* Контроль стабильного сохранения плазмид в клетке-хозяине (контроль стабильного поддержания).
* Контроль равномерного распределения дочерних плазмид в дочерние бактериальные клетки. Распределение плазмид между дочерними клетками происходит нерандомически, т. е. не по принципу случайности, а существует генетический механизм контроля равномерного распределения (сегрегации) вновь синтезированных плазмид при клеточном делении. Носительство плазмид для клетки-хозяина становится генетически необходимым, благодаря этому обеспечивается существование плазмид как организмов.
* Способность к самопереносу (у конъюгативных плазмид).
* Способность к мобилизации на перенос (у неконъюгативных плазмид).
* Способность наделять клетку-хозяина дополнительными важными для нее биологическими свойствами, способствующими выживанию бактерий, а следовательно, и плазмид в природе.
Важные св-ва плазмид:
*фертильность (способность женского или мужского организма к участию в оплодотворении);
*устойчивость к тяжелым металлам (кадмий, ртуть);
*способность к продукции бактериоцинов, антибиотиков, токсинов;
*способность к утилизации необычных источников углерода (камфора, октан, октанол).
Плазмиды патогенных микробов контролируют синтез различных факторов патогенности и образование новых способностей у многих видов бактерий:
Читайте также: