
Первичные культуры клеток для выделения вирусов
Обновлено: 12.05.2024
Лабораторные исследованияпри проведении идентификации вирусов и диагностике вирусных инфекций включают следующие этапы: выделение, культивирование, индикация (выявление) и идентификация вирусов.
2.3.1 Культивирование вирусов
Вирусы не растут на искусственных питательных средах, а размножаются только внутриклеточно. Крупным достижением было предложение Р. Гудпасчура в 1932 г. использовать для культивирования вирусов куриные эмбрионы. Окончательное решение проблемы культивирования вирусов оказалось возможным лишь после того, как были разработаны основные способы культивирования клеток вне организма.
Использование куриных эмбрионов. Куриные эмбрионы – практически идеальные модели для культивирования некоторых вирусов (например, гриппа и кори). Замкнутая полость эмбриона препятствует проникновению микроорганизмов извне, а также развитию спонтанных вирусных инфекций. Эмбрионы применяют для первичного выделения вирусов из патологического материала; для пассирования и сохранения их, а также для получения необходимых количеств вируса. Некоторые возбудители (например, герпесвирусы) вызывают характерные изменения (по ним можно распознавать заболевание).
Для заражения обычно используют куриные эмбрионы 7–12-дневного возраста. Перед заражением определяют жизнеспособность эмбриона путем овоскопирования (просматривают в проходящем свете). Живые эмбрионы при овоскопировании проявляют двигательную активность, хорошо виден сосудистый рисунок. Простым карандашом очерчивают границы воздушной камеры.
Куриные эмбрионы заражают вируссодержащим материалом в асептических условиях стерильными инструментами, предварительно обработав скорлупу над воздушным пространством йодом и спиртом. Заражение проводят на хорион-аллантоисную оболочку, в амниотическую или аллантоисную полость, либо в желточный мешок (рисунок 29). Выбор метода заражения зависит от биологических свойств вируса.
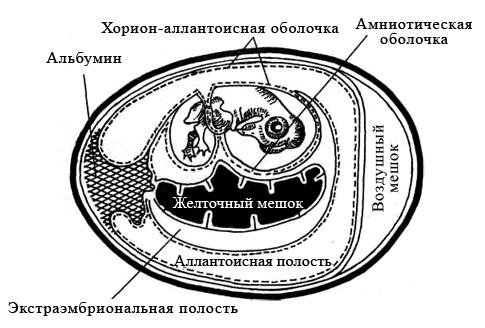
Рисунок 29 – Схематическое изображение развивающегося куриного эмбриона
Культура клеток. Вначале был использован метод переживающих тканей. Он заключался в том, что в колбу, содержащую питательную среду, вносили кусочек ткани. Клетки некоторых тканей в таких условиях могут переживать (но не размножаться) до 30 дней, а в них могут размножаться вирусы. Однако этот способ давал очень небольшой выход вирусов. Необходимо было разработать условия, при которых клетки ткани могли бы свободно размножаться.
Для получения культур клеток необходимо было решить четыре главных задачи:
– получить в необходимом количестве свободные (т. е. изолированные друг от друга) клетки;
– создать такие питательные среды и условия, в которых клетки могли бы активно размножаться;
– обеспечить условия, при которых в культурах клеток не могли бы размножаться бактерии;
– определить методы, с помощью которых можно было бы распознавать рост вируса в культуре клеток и идентифицировать его.
Для выделения изолированных (разобщенных), но жизнеспособных клеток из разрушенных тканей, стали использовать обработку их слабым раствором трипсина, разрушающего межклеточные мостики. Для культивирования клеток были предложены различные среды, содержащие все необходимые для размножения клеток питательные вещества (аминокислоты, основания, витамины и другие), минеральные соли, имеющие оптимальную рН и т. д. К питательным средам добавляли индикатор, по изменению цвета которого можно было судить о метаболизме клеток и их размножении. Было установлено, что в качестве основы, на которой клетки размножаются и образуют монослой, может быть использовано хорошо обработанное стекло пробирок и колб. Для подавления возможного роста бактерий вируссодержащий материал перед посевом его в культуры клеток стали обрабатывать антибиотиками.
В 1949 г. Дж. Эндерс, Т. Веллер и Ф. Роббинс показали, что вирус полиомиелита хорошо размножается в первично-трипсинизированных культурах клеток, полученных из почек обезьян. Основной недостаток первично-трипсинизированных клеток заключается в том, что после нескольких пересевов они перестают размножаться. Поэтому предпочтением стали пользоваться культуры таких клеток, которые способны размножаться in vitro бесконечно долго. Такие перевиваемые культуры клеток (клеточные линии характеризуются бессмертием и гетероплоидным кариотипом) получают из опухолевых тканей (HeLa получена из карциномы шейки матки, НЕр-2 – из карциномы гортани; Детройт-6 – из метастаза рака легкого в костный мозг; RН – из опухоли почки человека) или из мутантных клеток с полиплоидным набором хромосом. Однако опухолевые клетки нельзя применять для получения вакцин. Для этих целей используют только культуры таких клеток, которые не содержат никаких контаминантных вирусов и не обладают злокачественностью. Лучше всего этим требованиям отвечают культуры диплоидных клеток.

Полуперевиваемые (диплоидные) культуры клеток – клетки одного генотипа, способные in vitro выдерживать 50–100 пассажей, сохраняя при этом свой исходный диплоидный набор хромосом. Диплоидные линии фибробластов эмбриона человека используются как для диагностики вирусных инфекций, так и при производстве вирусных вакцин. Как оказалось, вирусы могут размножаться не только в культурах клеток, образующих монослой на стекле пробирок, но и в суспензиях живых клеток.
Для обеспечения жизнедеятельности культивируемых клеток необходимы питательные среды. По назначению они делятся на ростовые и поддерживающие. В ростовых питательных средах должно содержаться больше питательных веществ, обеспечивающих активное размножение клеток и формирование монослоя. Поддерживающие среды обеспечивают переживание клеток в уже сформированном монослое в период размножения в них вирусов.
2.3.2 Выделение вирусов
Выделение вирусов в культурах клеток. При выделении вирусов из различных инфекционных материалов (кровь, моча, слизистые отделяемые, смывы из органов) применяют культуры клеток, обладающих наибольшей чувствительностью к предполагаемому вирусу. Для заражения используют культуры в пробирках с хорошо развитым монослоем клеток. Перед заражением клеток питательную среду удаляют и в каждую пробирку вносят по 0,1–0,2 мл взвеси исследуемого материала, предварительно обработанного антибиотиками для уничтожения бактерий и грибов. После 30-60 мин контакта вируса с монослоем клеток удаляют избыток материала, в культуру вносят поддерживающую среду и пробы оставляют в термостате до выявления признаков размножения вируса.
Выделение вирусов на лабораторных животных. При невозможности выделить и идентифицировать вирус стандартными методами in vitro инфекционный материал вводят чувствительным к возбудителю животным, и после развития типичного инфекционного процесса проводят повторное заражение чувствительных клеточных культур. Наиболее часто используют мышей, кроликов и обезьян; для выделения некоторых вирусов (например, вирусов Коксаки) заражают мышат-сосунков. Вследствие дороговизны и сложности содержания лабораторных животных, практически повсеместно их вытеснили клеточные культуры. Тем не менее животные модели активно используют для изучения особенностей патогенеза и формирования иммунных реакций при вирусных инфекциях.
Таким образом, для выделения чистых культур вирусов в лабораторных условиях в настоящее время используются следующие живые объекты (биологические модели): 1) культура клеток (тканей, органов); 2) куриные эмбрионы; 3) лабораторные животные.
2.3.3 Индикация вирусов
Индикация вирусов в культурах клеток. Индикатором наличия вируса в зараженных культурах клеток может служить:
1) развитие специфической дегенерации клеток – цитопатическое действие вируса (ЦПД), имеющее три основных типа: крупно- или мелкоклеточная дегенерация; образование многоядерных гигантских клеток (симпластов); развитие очагов клеточной пролиферации, состоящих из нескольких слоев клеток (гроздевидная дегенерация клеток).
Различают два механизма гибели клеток, вызываемой вирусами, – некроз и апоптоз. Некроз происходит из-за необратимых нарушений целостности клеточных мембран, апоптоз – вследствие фрагментации ядерной ДНК под действием клеточной эндонуклеазы.
Цитопатические эффектыоценивают при микроскопии клеточных культур. По степени поражения клеток выделяют вирусы с высокой или умеренной цитопатогенностью:
2) обнаружение внутриклеточных включений, располагающихся в цитоплазме и/или в ядрах пораженных клеток;
3) положительная реакция гемагглютинации (РГА) или гемадсорбции (РГАдс). Некоторые вирусы, в частности, вирус гриппа, обладают особыми рецепторами (гемагглютининами), с помощью которых они адсорбируются на эритроцитах и вызывают их склеивание (гемагглютинацию). Такие вирусы легко обнаруживаются с помощью реакции гемагглютинации или гемадсорбции (эритроциты адсорбируются на инфицированных вирусами клетках культуры тканей);
4) феномен бляшкообразования. Широкое распространение получил предложенный в 1952 г. Р. Дюльбекко метод бляшек (негативных колоний), позволяющий производить количественное определение вирусов. Для выделения вирусов монослой клеток после удаления питательной среды заражают вируссодержащим материалом и покрывают слоем агара, содержащего индикатор нейтральный красный. Чашки (флаконы) инкубируют при 37 °С. Через 48–96 ч выявляются пятна – бляшки. Они имеют диаметр 1–3 мм и выглядят неокрашенными на розовом фоне. Пятна возникают за счет цитопатического действия вируса;
5) цветная реакция Солка. О росте вирусов в клетках можно судить с помощью индикатора, добавляемого к питательной среде. Если клетки активно осуществляют метаболизм, рН среды сдвигается в кислую сторону, и среда окрашивается в желтый цвет. В случае размножения вируса клетки погибают, рН среды мало меняется, и она сохраняет первоначальный (малиновый) цвет или (при нейтральной рН) приобретает оранжевый;
6) реакция интерференции (используется при отсутствии ЦПД, гемагглютинации и гемадсорбции): исследуемая культура повторно заражается вирусом, вызывающим ЦПД. В положительном случае ЦПД будет отсутствовать (реакция интерференции положительна). Если в исследуемом материале вируса не было, наблюдается ЦПД.
Кроме того, для обнаружения вируса в культурах клеток могут быть использованы различные серологические реакции.
Индикация вирусов на лабораторных животных. Индикация вируса основана на обнаружении у животных признаков инфекционного заболевания, регистрации их гибели, изучении характера патоморфологических и патогистологических изменений в тканях и органах, выявлении положительной реакции гемагглютинации.
2.3.4 Методы идентификации вирусов
Определение типа вируса (его идентификация) основано на нейтрализации биологической активности вируса с помощью типоспецифических сывороток. Конечный результат ее может быть установлен на основании следующих признаков:
1) нейтрализация цитопатического действия: в культуральную среду, содержащую изучаемый вирус, вносят коммерческую сыворотку (например, к вирусу краснухи при подозрении на неё), инкубируют и заражают вторую культуру; через 1–2 дня в неё вносят известный цитопатогенный вирус. При наличии цитопатогенного эффекта делают вывод о том, что первая культура была заражена вирусом, соответствовавшим антителам примененной сыворотки;
2) нейтрализация реакции гемадсорбции;
3) изменение проявления цветной пробы;
4) задержка (торможение) реакции гемагглютинации: смешивают культуральную среду, содержащую возбудитель, с известной коммерческой антисывороткой и вносят в культуру клеток. После инкубации определяют способность культуры к гемагглютинации и при её отсутствии делают заключение о несоответствии вируса антисыворотке.
5) нейтрализация в опытах на животных.
Таким образом РН (реакция нейтрализации) основана на подавлении соответствующей реакции, феномена, развития инфекционного процесса после внесения в культуру или введения в организм животного смеси вируса со специфичными AT, содержащимися в диагностической сыворотке.
Вопросы для самоконтроля
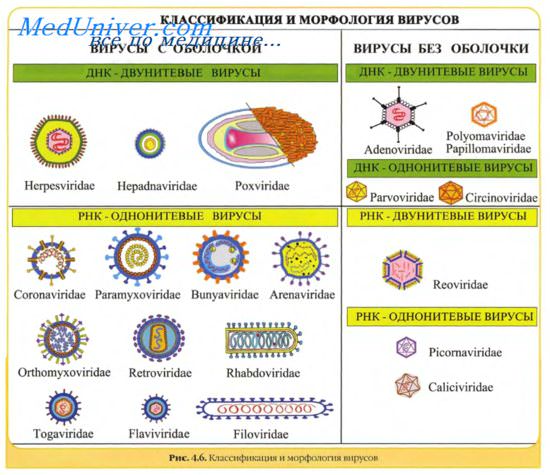
1 Назовите основные принципы классификации вирусов.
2 Приведите русские и латинские названия основных семейств вирусов человека и животных.
3 Назовите типовых представителей основных семейств вирусов и заболевания, вызываемые ими.
4 Каковы особенности морфологии и ультраструктуры вирусов человека и животных (основных семейств)?
5 Назовите РНК-геномные и ДНК-геномные фитовирусы.
6 Какие этапы включают в себя лабораторные исследования при идентификации вирусов и диагностике вирусных инфекций?
7 Какие биологические модели используются для выделения и культивирования вирусов человека и животных?
8 Как происходит заражение куриных эмбрионов в лабораторных условиях?
9 Какие методы получения культуры клеток вы знаете?
10 Как проводят идентификацию вирусов в курином эмбрионе и на лабораторных животных?
11 Какие существуют методы индикации вирусов на культуре клеток?
12 В чем заключается назначение и сущность реакций нейтрализации вирусов?
Культивирование вирусов. Биотехнологии в вирусологии.
Массовое выращивание клеток в культуре является центральным звеном любого технологического процесса, основанного на использовании клеток животных, и, в первую очередь, производства вирусных препаратов. Эта стадия определяет массу и качество клеток и, тем самым, в целом технологию получения вирусного сырья. Выбор способа культивирования вируса в значительной мере определяется способностью клеток размножаться на поверхности плотного субстрата или в суспензионной культуре. Трудно определить верхние и нижние границы крупномасштабного или промышленного выращивания вирусов в клеточных культурах. Все зависит от масштабов производства вирусных препаратов. В одних случаях речь идет о получении сотен литров, в других — десятков и даже сотен тысяч литров культурального вируса в год. Это зависит от вида вакцин и масштабов их применения. Изготовление живых вакцин при прочих равных условиях всегда требует меньших объемов вирусного сырья, нежели приготовление инактивированных, в особенности концентрированных вакцин.
Отсутствие эффективных вакцин для профилактики некоторых заболеваний объясняется, прежде всего, отсутствием экономичного способа получения иммуногенного материала в достаточном количестве.
В отличие от инактивированных вакцин против ящура и полиомиелита, выпускаемых в больших количествах, при многих заболеваниях человека и животных применяют живые вакцины, для изготовления которых не требуется большого количества вирусного сырья. Это, прежде всего, относится к тем вакцинам, которые перед применением разводят. Ежегодно производство таких вакцин, связанное с получением сотен литров культурального вируса, удовлетворяется использованием статических или вращающихся культуральных сосудов. Однако такие методы культивирования вирусов не могут удовлетворить крупномасштабное производство ряда вирусных вакцин. Например, при изготовлении противоящурной вакцины перевиваемые клетки выращивают в суспензии в реакторах с рабочим объемом более 1000 л. Крупные научно-производственные центры Южной Америки ежегодно вырабатывали до 600 млн. доз моновалентной инактивированной противоящурной вакцины. Для этой цели необходимо еженедельно получать около 20 000 л суспензионной культуры клеток ВНК-21.
До недавнего времени производство большинства вирусных препаратов основывалось на использовании первичных культур клеток из нормальных тканей различных видов домашних и лабораторных животных. Кроме того, в качестве клеточных субстратов для производства вакцин, применяемых в медицине, использовали немногие линии диплоидных клеток с ограниченной жизненной потенцией. Широкое применение таких препаратов в медицине и ветеринарной практике дало возможность достичь больших успехов в борьбе со многими опасными вирусными болезнями человека и животных. Однако первичные культуры клеток во многих отношениях не являются перспективными клеточными субстратами. Их приготовление связано с периодическим убоем животных и необходимостью выделения клеток из тканей.
Первичные культуры отличаются нестандартностью, таят в себе опасность в отношении эндогенной контаминации различными вирусами и микроорганизмами. Наконец, их сложно выращивать в условиях крупносерийного производства. Отмеченные трудности значительно возрастают в связи с тенденцией постоянного увеличения масштабов изготовления противовирусных препаратов. Кроме того, в последнее время все более широкое развитие получает разработка концентрированных и субъединичных вакцин, при изготовлении которых требуется большое количество вирусного материала. Естественно, что стоящая задача может быть решена лишь путем использования постоянных (перевиваемых) линий клеток, отличающихся способностью к бесконечной пересеваемости вне организма, высокой стандартностью, низкой стоимостью, относительной простотой трансфекции рекомбинантной ДНК и последующего клонирования высокоэффективных продуцентов, высокой вероятностью правильного посттрансляционного процессинга вновь синтезируемых белков, кодируемых трансфецирующей ДНК.
Ветеринарная наука в течение последней четверти века накопила большой опыт в изготовлении вирусных вакцин с использованием в качестве субстрата для размножения вирусов культур постоянных линий клеток животных. Особый успех достигнут в изготовлении инактивированной противоящурной вакцины. Производство вакцин против ящура имеет наиболее цитируемую технологию. Она основана на использовании линии трансформированных клеток новорожденного хомяка, выращиваемых в суспензии. Эта технология достаточно экономична, ее выполняют в биореакторах большой емкости. Накоплены определенные доказательства безопасности некоторых постоянных клеточных линий, используемых в качестве субстрата в производстве ряда биологических препаратов. Например, инактивированную противоящурную вакцину готовят из вируса, выращенного в культуре постоянной линии клеток почки новорожденного хомяка (линия ВНК-21). Более чем за 20-летний период этой вакциной привито свыше 100 млн. голов крупного рогатого скота и не обнаружено каких-либо нарушений у привитых животных, по крайней мере, в течение 2—4 лет после введения вакцины. Имеется много других примеров безопасности применения инактивированных и даже живых вакцин против ряда болезней животных, приготовленных из вирусов, размноженных в культурах различных постоянных линий клеток. Применение биологических препаратов, полученных на основе перевиваемых клеточных линий, в медицине началось намного позже, чем в ветеринарной практике.
Несмотря на очевидные преимущества постоянных линий клеток, медицинская практика до недавнего времени воздерживалась от их применения в производстве вирусных вакцин. Причина заключалась в том, что, согласно существовавшему мнению, для изготовления медицинских вирусных вакцин можно было использовать клетки из тканей только клинически здоровых животных. Производство таких вакцин ограничивалось использованием первичных и диплоидных культур клеток. В диплоидных линиях клеток человека никогда не были обнаружены латентные вирусы или спонтанная трансформация клеток. Основное возражение против использования постоянных клеточных линий для репликации вирусов и векторных рекомбинатов в производстве вирусных вакцин медицинского назначения заключалось в их возможной онкогенности из-за контаминации вакцин клеточной ДНК или генными продуктами (регуляторными белками). Интеграция гетерогенной ДНК может привести к предзлокачественным изменениям в результате активации протоонкогенов, запуску онкогенов и инактивации генов опухолевой супрессии. В процессе репродукции вакцинных штаммов для живых вакцин с использованием клеточных линий, латентно контаминированных другими вирусами, могут появляться вирусные гибриды с неожиданными свойствами.
Этот вопрос рассматривался неоднократно на различных научных форумах в Европе и Северной Америке. Ценность постоянных клеточных линий в качестве субстратов стала особенно очевидной благодаря успехам, достигнутым в последнее время в области фундаментальных биологических исследований, а также в связи с перспективой их использования в рекомбинантной ДНК-технологии и получении генно-инженерных биопрепаратов. Общая тенденция к применению постоянных клеточных линий в производстве медицинских иммунобиологических препаратов наметилась на рубеже 70—80-х годов. Так, в 1978 г. в Лейк-Плесиде (США) было предложено использовать лимфобластоидные клетки человека для крупномасштабного производства альфа-интерферона. В 1981 г. Комитет экспертов ВОЗ по стандартизации биологических препаратов одобрил применение неопухолевых и неконтаминированных вирусами постоянных клеточных линий для производства инактивированной полиомиелитной вакцины, а затем также для инактивированной вакцины против бешенства. Такое решение дало возможность в короткий срок разработать методы крупномасштабного выращивания вирусов с использованием микроносителей и создать высокоэффективные вирусные вакцины.
В истории создания биологических препаратов ключевая роль всегда принадлежала выбору приемлемо безопасных вариантов. Решение о применении людям биопрепаратов, полученных с использованием постоянных клеточных линий, основывалось на оценке различными комитетами выгод и риска, связанных с созданием новых препаратов, по сравнению с существующими. Важнейшие потенциальные факторы риска, связанные с биологическими препаратами, производимыми на постоянных клеточных линиях, можно разделить на три категории: примесь гетерогенной ДНК, вирусы и трансформирующие белки.
Одним из основных вопросов, требующих самого пристального внимания, является потенциальная долгосрочная опасность, связанная с присутствием в препаратах примесей гетерогенных ДНК, особенно в тех случаях, когда последние могли содержать потенциально онкогенные кодирующие или регуляторные последовательности.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Среды для выращивания вирусов. Клеточные субстраты в вирусологии.
В течение многих лет разрабатывались методы выращивания небольших количеств клеток животных в лабораторных условиях. Однако наладить массовое культивирование таких клеток оказалось не простой задачей. Способ выращивания, характер используемой среды, методы управления и контроля в значительной степени зависят от типа выращиваемых клеток. Все культивируемые клетки первоначально получают от животных путем механической или ферментативной дезагрегации нормальных или малигинизированных тканей либо с помощью перфузии in vivo.
Первичные культуры обычно получают трипсинизацией тканей куриных эмбрионов или тканей (чаще всего почек), взятых от других видов здоровых животных. Возраст используемых эмбрионов может значительно различаться и влиять на выход и жизнеспособность клеток. Для более полного контроля доноров ткани их следует брать из SPF-хозяйств.
Первичные культуры клеток почек зеленых мартышек обычно используют для приготовления полиовирусной вакцины. Поскольку обезьяны являются очень дорогим объектом для увеличения выхода клеток почки дезагрегируют методом перфузионной трипсинизации in vivo, а затем первичную культуру выращивают на микроносителях. Для изготовления других медицинских вакцин широко применяют первичную культуру клеток почки кроликов, телят и других животных. Для изготовления вакцин, используемых в ветеринарной медицине, чаще всего применяют первичные культуры клеток куриных эмбрионов и почек естественного хозяина или восприимчивых животных.
Первичные культуры в зависимости от типа ткани и условий выращивания значительно различаются. Некоторые из них погибают через несколько дней после посева, тогда как другие могут длительно сохраняться в культуре без заметных морфологических и биохимических изменений. С момента приготовления первичной культуры до остановки роста клеток и гибели культуры после серийных пересевов проходит от нескольких недель до нескольких месяцев. Клетки эмбрионального происхождения при культивировании обычно сохраняют жизнеспособность более продолжительное время.
Линия клеток - первичная культура, со времени получения субкультуры клеточные линии могут иметь ограниченный срок жизни in vitro (например, диплоидные фибробласты человека и животных), а могут размножаться in vitro неограниченно долго. В последнем случае их называют постоянными, стабильными или непрерывными клеточными линиями.
В связи с трудностями получения соответствующих количеств первичных культур клеток почки зеленой мартышки (по причине стоимости) и преимуществами использования предварительно изученных клеточных культур для производства вакцины против полиомиелита и других вакцин были лицензированы диплоидные линии клеток WI-38 и MRC-5. Существуют сомнения в том, что ростовые свойства и продуктивность диплоидных клеток варьируют. Даже хорошо охарактеризованные диплоидные линии клеток ведут себя неодинаково при работе с ними различных исследователей.
Штамм клеток — популяция однородных клеток (по одному или нескольким маркерам), сохраняющая специфические свойства в течение ограниченного периода культивирования. Штамм клеток может происходить из первичной культуры либо от линии клеток, получающих специфические свойства путем селекции или клонирования.
Линия гетероплоидных клеток. Этот термин означает, что клеточная линия имеет менее 75% клеток с диплоидным набором хромосом. Этот термин не означает, что клетки являются злокачественными или способны расти до бесконечности.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Для культивирования вирусов в лабораторных условиях используются следующие живые объекты: 1) культуры клеток (тканей, органов); 2) куриные эмбрионы; 3) лабораторные животные.
I. Культуры клеток
Наибольшее распространение имеют однослойные культуры клеток, которые можно разделить на первичные (первично трипсинизированные), полуперевиваемые (диплоидные), перевиваемые, трансфецированные.
По происхождению они подразделяются на эмбриональные, опухолевые и из взрослых организмов; по морфогенезу — на фибробластные, эпителиальные и др.
Первичные культуры клеток — это клетки какой-либо ткани человека или животного, способные культивироваться в виде монослоя на пластмассовой или стеклянной поверхности в специальной питательной среде, но не способные к длительному размножению. Срок жизни таких культур ограничен. В каждом конкретном случае их получают из ткани после механического измельчения, обработки протеолитическими ферментами и стандартизации количества клеток. Первичные культуры, полученные из почек обезьян, почек эмбриона человека, амниона человека, куриных эмбрионов, широко используются для выделения и накопления вирусов, а также для производства вирусных вакцин.
Полуперевиваемые (диплоидные) культуры клеток — клетки одного генотипа, способные in vitro выдерживать до 50100 пассажей, сохраняя при этом свой исходный диплоидный набор хромосом. Диплоидные линии фибробластов эмбриона человека используются как для диагностики вирусных инфекций, так и при производстве вирусных вакцин.
Перевиваемые клеточные линии характеризуются бессмертием и гетероплоидным кариотипом. Источником перевиваемых линий могут быть первичные клеточные культуры (например, СОЦ — из сердца обезьяны циномольгус, ПЭС — из почек эмбриона свиньи, ВНК-21 — из почек однодневных сирийских хомяков; ПМС — из почки морской свинки и др.), отдельные клетки которых обнаруживают тенденцию к бесконечному размножению in vitro. Совокупность изменений, приводящих к появлению в клетках таких свойств, называют трансформацией, а клетки перевиваемых тканевых культур — трансформированными.
Другой источник перевиваемых клеточных линий — злокачественные новообразования. В этом случае трансформация клеток происходит in vivo. Получены и наиболее широко в вирусологической практике применяются следующие линии перевиваемых клеток: HeLa — получена из карциномы шейки матки; Hep-2 — из карциномы гортани; Детройт-6 — из метастаза рака легкого в костный мозг; RH — из опухоли почки человека.
Трансфецированные культуры клеток. Разработаны экспериментальные линии культур клеток методом трансфекции (переноса) генов вирусов, контролирующих биосинтез поверхностных антигенов. Такие культуры клеток экспрессируют поверхностный белок определенного вируса (HBs-антиген, gp120 и др.) на мембране клеток культуры. Такие культуры клеток используются с целью изучения иммунологических механизмов патогенеза вирусных инфекций, разработки химиотерапевтических и иммунобиологических препаратов.
Для обеспечения жизнедеятельности культивируемых клеток необходимы питательные среды. По назначению они делятся на ростовые и поддерживающие. В ростовых питательных средах должно содержаться больше питательных веществ, обеспечивающих активное размножение клеток и формирование монослоя. Поддерживающие среды обеспечивают переживание клеток в уже сформированном монослое в период размножения в них вирусов.
Широкое применение находят стандартные синтетические среды, например, синтетическая среда 199 и среда Игла. Независимо от назначения все питательные среды для культур клеток конструируются на основе сбалансированного солевого раствора. Чаще всего им является раствор Хенкса. Неотъемлемый компонент большинства ростовых сред — сыворотка крови животных (телячья, бычья, лошадиная), без наличия 510% которой размножение клеток и формирование монослоя не происходит. В состав поддерживающих сред сыворотка не входит. С целью предотвращения возможного роста микроорганизмов в питательные среды вносят антибиотики.
Выделение вирусов в культурах клеток и методы их индикации
При выделении вирусов из различных инфекционных материалов от больного (кровь, моча, фекалии, слизистые отделяемые, смывы из органов) применяют культуры клеток, обладающие наибольшей чувствительностью к предполагаемому вирусу. Для заражения используют культуры в пробирках с хорошо развитым монослоем клеток. Перед заражением клеток питательную среду удаляют и в каждую пробирку вносят по 0,10,2 мл взвеси испытуемого материала, предварительно обработанного антибиотиками для уничтожения бактерий и грибов. После 3060 мин. контакта вируса с монослоем клеток удаляют избыток материала, в культуру клеток вносят поддерживающую среду и пробы оставляют в термостате до выявления признаков размножения вируса.
Индикатором наличия вируса в зараженных таким образом культурах клеток может служить:
развитие специфической дегенерации клеток — цитопатическое действие вируса (ЦПД), имеющее три основных типа: кругло- или мелкоклеточная дегенерация; образование многоядерных гигантских клеток (симпластов); развитие очагов клеточной пролиферации, состоящих из нескольких слоев клеток;
обнаружение внутриклеточных включений, располагающихся в цитоплазме и/или в ядрах пораженных клеток;
положительная реакция гамагглютинации (РГА) или гемадсорбции (РГАдс);
при отсутствии ЦПД, ГА или ГАдс. можно использовать реакцию интерференции: исследуемая культура повторно заражается вирусом, вызывающим ЦПД. В положительном случае ЦПД будет отсутствовать (реакция интерференции положительная). Если в исследуемом материале вируса не было, наблюдается ЦПД.
Читайте также: