
Размер молекулярная масса вируса осповакцины
Обновлено: 18.04.2024
Нуклеоид содержит ДНК, ассоциированную с белками. Способ ее упаковки неясен, хотя из комплекса ДНК— белок выделены 3 белка с молекулярной массой 33 000, 28 000 и 12 000, последний белок богат аргинином. Напрашивается аналогия с упаковкой ДНК хромосом эукариотов гистоновыми и негистоновыми белками. Нуклеоид окружен внутренними оболочками. Внешние оболочки состоят из липидов и углеводов. Оболочки построены сложно и состоят из слоев трубчатых или глобулярных структур. Разные роды вирусов различаются преимущественно деталями строения внешних оболочек.
Геном вирионов представляет собой линейную двунитевую ДНК с молекулярной массой 85x10 6 —250x10 6 , концы которой соединены ковалентно, образуя терминальные петли и инвертированные тандемные повторы. Молекулярная масса генома не может служить таксономическим признаком, так как у серологически родственных вирусов она колеблется от 85x10 6 (вирус паравакцины) до 200x10 6 (вирус оспы птиц). Содержание Г+Ц у вирусов оспы позвоночных составляет 5— 7,5% от массы вириона; состав нуклеотидов: А — 29,5%, Ц — 20%, Г — 20,6%, Т — 29,9%. Геном вируса осповакцины содержит всего 240 000 пар нуклеотидов и имеет длину 82 мкм. Необычная прочность ДНК этого вируса и других вирусов данной группы обусловлена ковалентными связями комплементарных нитей на обоих концах молекулы ДНК, примерно на расстоянии 50 нуклеотидов от ее концов [см. Слепушкин А. Н., 1982]. Как было показано при изучении вируса осповакцины, ее ДНК, подобно ДНК многих других вирусов, содержит терминальные инвертированные повторы, которые, как предполагается, играют важную роль в репликации ДНК. У вируса осповакцины эти повторы весьма велики — молекулярная масса около 7х10 6 , или около 10 000 пар нуклеотидов. Они могут быть частично транскрибированы, кодируя синтез ранних мРНК [Wittek R. et al., 1980].
На геноме вирусов оспы закодировано несколько сотен белков (глико- и фосфопротеидов) с молекулярной массой 80 000— 200 000. По данным разных авторов, для различных вирусов число вирусспецифйческих белков варьирует. Ранее указывалось, что вирионы осповакцины содержат более 80 белков, суммарная молекулярная масса которых составляет около 60% массы генома [Fenner F., 1979]. У вируса оспы Яба обнаружено 37 структурных белков с молекулярной массой 10 000—220 000 [Fenger.T., Rouhandch Н., 1976]. Более поздние данные свидетельствуют, что вирус осповакцины имеет 279 белков, в том числе 13 гликопротеидов [Carrasco L., Bravo R., 1985]. Пока идентифицированы лишь немногие вирионные и и невирионные белки. Нейтрализация вируса связана с поверхностным белком (58 000), гемагглютинация — с гликопротеидом (85 000). Другие мажорные белки обозначают символами,2Ь, 4а, 4Ь, 6а, 6b, 8, 11b; два белка (4а, 4b) составляют 50% суммарных белков сердцевины, масса которой в свою очередь составляет около 50% массы вириона. В наружных оболочках вириона найдено 8 белков, а на самой поверхности — 5 белков. В составе вирионов имеется или в процессе репродукции образуется не менее 10 ферментов, в том числе РНК-полимераза (вирионная транскриптаза), ферменты, катализирующие образование и метилирование кэп-структур, которые обеспечивают синтез и процессинг мРНК, протеин- и тимидинкиназа. По-видимому, и синтез вирусной ДНК катализируется в основном вирусспецифическими ферментами [Slabaugh М., Mattews С., 1984].
Репликация вирусов оспы (рис. 61) происходит в цитоплазме ступенчато, включая первичное раздевание (uncoating I), «синтез ранних мРНК и белков, вторичное раздевание (uncoating II), репликацию ДНК, синтез поздних мРНК и белков, входящих в состав вирионов, их гликозилирование и фосфорилирование, образование предшественников вирионов, созревание их и выход из клетки [Shida Н., 1986].
Хотя имеются данные об участии клеточного ядра в репродукции вирусов оспы [Miningan Н. et al., 1985], но оно не обязательно, так как весь цикл репродукции вирусов оспы можно воспроизвести в безъядерных клетках [Villareal Е. et al., 1984]. Вместе с тем репродукция этих вирусов, если даже абсолютно исключить клеточно-зависимые синтезы белка, тесно связана с клеточным метаболизмом. Об этом свидетельствуют возможность репродукции штаммов вируса оспoвакцины, дефектных по некоторым генам (например, по гену тимидинкиназы), возможность замены этого гена геном герпесвируса, а также наличие в геноме вируса достаточно обширных нереплицирующихся областей. Именно на использовании последних в значительной мере основаны генно-инженерные исследования, в результате которых вирус осповакцины со встроенными в него чужеродными генами становится экспрессирующим их вектором (см. ниже).

Рис. 61. Цикл репликации вируса осповакцины (схема).
Места, в которых происходят репликация ДНК и формирование вирионов, являются структурами клеточной цитоплазмы, модифицированными вирусом; их обозначают как вирусные фабрики. Более 100 белков, синтезирующихся в первые часы инфекции, обеспечивают вторую стадию раздевания, репликацию вирусной ДНК и транскрипцию поздних генов. Среди ферментов, синтезируемых в этой стадии, следует назвать тимидинкиназу, ДНК-полимеразу, полинуклеотидлигазу, ферменты синтеза ДНК.
Репликация вирусной ДНК связана с образованием разрывов терминальных петель, самопраймированием 3'-концов: вирусной ДНК, но в общем многие детали синтеза вирусной ДНК мало изучены, и поэтому предложено несколько схем ее репликации. Поздняя транскрипция охватывает большую часть генома. На этой стадии транслируются структурные белки, входящие в состав вирионов. Подсчитано, что 7з генома вирусов оспы кодирует ранние, 1/3 — структурные и 1/3 — поздние неструктурные белки [Pennington Т., 1976]. В транскрипции поздних генов, по-видимому, участвует клеточная РНК-полимераза II. Часть белков, синтезируемых на этой стадии, подвергается протеолитическому расщеплению, которое связано, со сборкой вирионов и их предшественников.
Вирусы оспы вызывают глубокое угнетение синтеза клеточных макромолекул, причем эти синтезы блокируются как вирионными белками, так и белками, синтезирующимися в ходе репродукции вируса. Некоторые представители семейства вирусов оспы вызывают пролиферацию зараженных клеток и их: неопластическую трансформацию (вирусы фибромы, Яба, контагиозный вирус моллюска). Для вирусов оспы характерны такие генетические процессы, как рекомбинация внутри родов, спасение маркера [Nakano Е. et al., 1982] и негенетическая реактивация между родами вирусов оспы позвоночных. Вирусы оспы, циркулирующие в природе, подвергаются процессам рекомбинации. Одним из таких рекомбинантов является злокачественный вирус кроликов, явившийся рекомбинантом 10% генома вируса фибромы кроликов и 90% генома вируса миксомы кроликов [Block W. et al., 1985].
Более подробно описание особенностей генетики вирусов оспы можно найти в монографиях и обзорах [см. Гендон Ю. З., 1975]. Здесь же мы хотели бы отметить некоторые важные особенности, характерные, впрочем, не только для вирусов оспы, но и для других, ранее упоминавшихся ДНК-содержащих вирусов (герпесвирусы, бакуловирусы, аденовирусы и др.). Во-первых, ДНК вирусов оспы, близких между собой, или мутантов, легко рекомбинируются, образуя как гомологичные, так и гетерологичные рекомбинанты. В последнем случае образующиеся гетеродуплексы могут включать в себя чужеродные гены. Во-вторых, рекомбинация может происходить не только между полными геномами двух вирусов оспы, но и между полным геномом и сравнительно небольшим его фрагментом (4х10 6 ), включенными в плазмиду, при котрасфекции. В-третьих, в геноме вирусов оспы имеются большие участки, которые условно обозначают как несущественные для репликации области генома. Они имеют размер около 25 000 пар нуклеотидов, т. е. более 10% всего генома [Smith G., Moss. В., 1983]. По-видимому, этой областью кодируется синтез белков репликации и других, которые могут быть заменены клеточными белками, как например, тимидинкиназа, а может быть, и белки, обеспечивающие вирулентность вируса. Все эти особенности делают вирус осповакцины весьма удобным вектором для включения в него чужеродных генов. Мы не будем здесь цитировать уже многочисленную литературу, посвященную использованию вируса оспованцины для генно-инженерных операций; сошлемся лишь на меморандум экспертов ВОЗ по этому вопросу [WHO, 1985]. Важно подчеркнуть другое: при генно-инженерных операциях, т. е. искусственных манипуляциях, используют естественные существующие генетические взаимосвязи у этой и других групп вирусов.
Кстати, именно эта особенность вирусов оспы послужила препятствием для применения рекомбинантного вируса осповакцины, содержащего экспрессируемый ген протективного гликопротеида вируса бешенства, для иммунизации лисиц. При поедании мяса, смоченного суспензией такого вируса, у лисиц и других хищников развивается осповакцинный стоматит, сопровождающийся выработкой иммунитета одновременно против оеповакцины и бешенства. Однако комитет экспертов ВОЗ решительно возразил против такого проекта, намечавшегося к проведению в Аргентине. Имелась в виду опасность появления в природе рекомбинантов вирусов оспы с повышенной нейровирулентностью, которая вызвана или связана с геном вируса бешенства, введенного в ДНК вируса осповакцины.
Вирусы оспы вызывают разнообразные заболевания, и все же наиболее выраженно они проявляются в виде высыпаний на коже и слизистых оболочках. Соответственно у животных заболевания передаются при контакте, через корм и питье, а у человека также воздушно-капельным путем. Особое место занимают неопластические процессы, поражающие либо наружные покровы, либо более глубоко лежащие ткани (фиброма). В последнем случае возможно участие в передаче кровососущих насекомых.
К сожалению, еще не сделана проверка эволюционных соотношений вируса оспы с прокариотами (эукариотами и/или архебактериями), так как все эти проверки, построение филогенетических древ и т. п. делались чаще всего на основе анализа рибосомных РНК (265, 165, 55). Последние имеются даже у потомков далеко эволюционировавших эндосимбионтов (митохондрии, пластиды). А этого как раз и нет у оспенных вирусов.

Рис. 62. Эволюционное древо белка вируса осповакцины сходных доменов у млекопитающих. Обведенные пунктиром овалы обозначают неопределенность ветвей древа.
С геномами эукариотических клеток геномы оспенных вирусов сближает наличие тандемных повторов в инвертированных терминальных повторах, что было показано при изучении вируса осповакцины. Предполагают, что они облегчают циклизацию однонитевых ДНК во время репликации. Эти повторы имеют размеры 70—150 пар нуклеотидов и повторяются тандемно 13—30 раз [Wittek R., Moss В., 1980].
Таким образом, происхождение вирусов оспы остается неясным. С бактериями его сближают громадный геном, независимая от клетки репликация и отсутствие сплайсинга. Однако структура их генома сходна и со структурой генома эукариотов, а некоторые гены имеют явно эукариотическое происхождение, если не считать их результатом далеко зашедшей молекулярной конвергенции.
Несмотря на большое сходство плана строения и морфологии вирионов оспы позвоночных и оспы насекомых, наличие по крайней мере 4 ферментов с одинаковыми функциями, размножение в цитоплазме и ряд других свойств, указывающих на общность происхождения обеих групп вирусов оспы, в эволюции всей группы вирусов остается много неясного. По-видимому, более древнее происхождение имеют вирусы оспы насекомых уж по одному тому, что насекомые — гораздо более древние формы организмов, нежели теплокровные позвоночные. Вирусы оспы не обнаружены у морских беспозвоночных, и если это факт, а не недостаток знаний, то можно предположить, что вирусы оспы появились не ранее силура, т. е. около 400 млн лет назад, или еще позже, в карбоне.
Освоение ими новой экологической ниши можно связать с развитием кровососущих насекомых, так как некоторые вирусы оспы позвоночных до сих пор могут передаваться кровососущими насекомыми. Кстати, все до сих пор известные вирусы оспы позвоночных поражают теплокровных — млекопитающих или птиц, а вирусы насекомых — как некровососущих (чешуекрылые), так и кровососущих (двукрылые), в том числе комаров-кровососов теплокровных животных. Поэтому образование вирусов оспы позвоночных следует отнести к сравнительно позднему периоду — юрскому или меловому, т. е. 120—150 млн лет назад или еще позже. Именно этим молено объяснить умеренную дивергенцию, существующую между 6 группами (родами) вирусов оспы позвоночных, вплоть до сохранения общих антигенов и еще более широких антигенных связей между вирусами одного и того же рода.
Эволюция этих вирусов происходила путем заполнения ими новых экологических ниш. Она сопровождалась затем, узкой специализацией вплоть до поражения соответствующим вирусом только одного вида. Так появились и эволюционировали вирусы оспы буйволов, верблюдов, кроликов, мышей (группа вирусов осповакцины), узелков доильщиц, контагиозной эктимы, пустулярного стоматита коров (группа вирусов, паравакцины), оспы канареек, голубей, перепелок, ласточек, воробьев, индюшек (группа вирусов оспы птиц), оспы овец, коз, фибромы и миксомы кроликов, оспы свиней. Приручение человеком животных, с одной стороны, усилило специализацию вирусов (оспа овец, коз, верблюдов), с другой — интенсифицировало распространение вирусов, в результате чего одни и те же виды домашних животных стали поражаться разными вирусами оспы. Например, коровы поражаются вирусами осповащины, коровьей оспы, паравакцины, узелков доильщиц; кролики — вирусами кроличьей оспы, фибромы и миксомы. Впрочем, вопрос об узкой специализации не столь уж прост. Мы говорим об оспе коров, но на самом деле речь идет обоспе, которой болеют коровы, но резервуаром которой являются, вероятно, грызуны. Вирус оспы обезьян выделен от белок [Khodakevich L., 1985]. При анализе вируса оспы коров была установлена широкая шкала патогенности, включающая 9 порядков млеко, питающих (рогатый скот, кенгуру, носороги, дельфины, кошачьи, грызуны). Все эти данные позволяют заключить, что грызуны являются основным резервуаром возбудителя в природе и источником заражения коров и других млекопитающих [Berkovitz Е., Pogo В., 1984]. По данным ВОЗ, оспа коров на самом деле является оспой у коров, а оспа обезьян — оспой у обезьян. В последнем случае установлено, что резервуаром оспы обезьян являются белки, а обезьяны — вторичные источники инфекции [WHO, 1986].
При молекулярно-биологическом изучении вируса фибромы Шоупа было показано, что этот вирус наиболее близок к ортопоксвирусам и в то же время имеет часть генома, близкую к геному лепорипоксвирусов [Berkovitz Е., Pogo В., 1985]. Таким образом, рекомбинации вирусов оспы имеют место в природе и некоторые вирусы имеют, несомненно, рекомбинантное происхождение [Berkovitz Е., Pogo В., 1985].
Но вернемся к исходным и последующим этапам эволюции вирусов оспы животных [Жданов В. М., Львов Д. К., 1984].
При оспенных заболеваниях птиц и млекопитающих поражаются кожа и слизистые оболочки. Заболевание сопровождается вирусемией и длительным носительством. В этом отношении характерна эктромелия мышей, при которой носительство может быть пожизненным. Все эти заболевания передаются либо при контакте, либо алиментарным путем, либо посредством кровососущих насекомых. Заболевание обычно поражает молодых животных, у которых инфекция может протекать остро. Выжившие животные становятся длительными, нередко, как это имеет место при эктромелии, пожизненными носителями и заражают затем свое потомство.
Приручение домашних животных и появление больших стад привели к тому, что болезни типа тех, которые вызывают параоспенные вирусы, эволюционировали по пути становления инфекций с более острым течением, что обеспечивало более быстрое распространение возбудителей в стаде. Более острое течение болезни стимулировало выработку более совершенного иммунитета, исключавшего возможность длительной латентной инфекции. Такого типа инфекция не могла бы существовать среди диких животных, но в больших стадах домашних животных при выраженном контакте между ними существование острой инфекции без длительного носительства было возможным. При наличии многих видов животных, прирученных человеком, создавалась возможность взаимного обмена паразитами (вирусами), образования экологических разновидностей, а также самостоятельных заболеваний в результате адаптации вирусов оспы к организму домашних животных. Следует подчеркнуть, что различные оспенные заболевания могли возникнуть среди стадных животных и менее вероятным являлось приспособление вирусов к тем видам животных, которые не разводились в стадах. Не удивительно потому, что известные до сих пор оспенные заболевания типа выраженной острой инфекции не поражают таких домашних животных, как кошки и собаки. Они распространены только среди стадных животных, домашней птицы (осподифтерит кур), а также среди мышевидных грызунов (эктромелия).
Итак, уже на ранних стадиях развития общества (поздний период варварства), когда появились большие стада домашних животных, создались условия для возникновения оспенных заболеваний этих животных с острым и подострым течением. Заражение животных происходило преимущественно алиментарным путем, в связи с чем вирусы вызывают поражение не только кожи, но и слизистых оболочек. Поскольку заражение происходило алиментарным путем, вирусы могли сохраниться лишь при условии их высокой устойчивости к неблагоприятным влияниям факторов внешней среды. И действительно, возбудители оспенных заболеваний диких животных отличаются устойчивостью и способностью длительно сохраняться во внешней среде.
Оспенные заболевания возникли в странах Старого Света, так как в Америке до открытия ее европейцами, кроме лам, не было других видов прирученных животных, а ламы не болеют естественными оспенными заболеваниями. Впервые большие стада домашних животных появились в Юго-Западной и Средней Азии, а также в Северной Африке (Египет). Здесь, по всей вероятности, и возникли оспенные заболевания домашних животных.
Такие же варианты изолированы и от диких грызунов в тех же районах Экваториальной Африки (табл. 21).
На основании данных табл. 21 можно предположить, что источником происхождения вируса оспы человека могли быть белые варианты вирусов оспы обезьян или грызунов Экваториальной Африки. Известно, что вирус оспы обезьян патогенен для человека, он вызывает генерализованный процесс, но в обычных условиях не передается от человека к человеку. Однако нельзя исключить, что болезнь может передаваться от человека к человеку контактным путем, такой путь передачи заразного начала вполне возможен в примитивных бытовых условиях.
Генерализация процесса резко повышает заразность больного, так как в этом случае наличие сыпи на слизистых оболочках полости рта и зева (энантема) и мацерация их создают новые дополнительные возможности для передачи инфекции воздушно-капельным путем. Заразность больного резко возрастает, становится возможным развитие самостоятельных вспышек среди людей. Надо полагать, что такого рода вспышки возникали не раз, однако в условиях первобытно-общинного строя они не могли широко распространиться и ограничились пределами рода или племени.
Таблица 21. Сравнительные свойства некоторых вирусов оспы хFenner F., 19771
Натуральная оспа (variola vera) — особо опасная вирусная инфекция, проявляющаяся интоксикацией, лихорадкой и пустулёзно-папулёзной сыпью.
Оспа одна из древнейших инфекционных болезней человека. Первые упоминания об натуральной оспе относятся к 3730-3710 гг. до н.э. (папирус Аменофиса I). В Европу возбудитель проник в VI веке, в Россию был занесён в XV-XVI веках, в Америку — в XVI веке.
До открытия Дженнером метода вакцинации европейские врачи были бессильны перед натуральной оспой. В отдельные годы смертность от оспы достигала 1,5 млн человек.
Возбудитель оспы — самый крупный вирус (220x300 нм), имеет кирпичеобразную форму с закруглёнными углами.
В состав вируса натуральной оспы входят 30 белков, более 10 из них — ферменты, катализирующие синтез нуклеиновых кислот. Вирус натуральной оспы проявляет гемагглютинирующие свойства; гемагглютинин состоит из трёх гликопротеинов.
Важнейшие Аг вируса оспы — нуклеопротеид NP (общий для всего семейства), термостабильный и термолабильный Аг, а также группа растворимых Аг. Размеры вирионов позволяют обнаружить их световой микроскопией в форме телец включений.
Внутриклеточные (точнее околоядерные) включения в клетках роговицы заражённого кролика впервые обнаружил итальянский патолог Д. Гварнери (1892). ЭТ возбудителя натуральной оспы в отделяемом везикул открыл немецкий бактериолог Э. Пашен (1906). Позднее было установлено наличие двух штаммов вируса натуральной оспы.
Первый вызывает классическую оспу (variola major) с летальностью, превышающей 50%, второй — алястрим (variola minor) с более лёгким течением и летальностью, не превышающей 1%. Возбудители идентичны по основным свойствам. Отличительные особенности — способность вируса алястрима размножаться в куриных эмбрионах при более низкой температуре (37,5 °С), образовывать бляшки и проявлять цитопатичес-кий эффект в культуре куриных фибробластов при температуре 37 °С.
К вирусу натуральной оспы чувствителен человек и приматы; экспериментальное внутримозговое заражение новорождённых мышат приводит к развитию генерализованной инфекции. Взрослые мыши нечувствительны к вирусу.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Геном вирусов оспы. Антигены вирусов оспы.
Геном вирусов оспы представлен одной линейной молекулой двуцепочечной ДНК с ковалентно замкнутыми концами размером 130 тпн (парапоксвирусы) — 280 тпн (авипоксвирусы). На обоих концах генома имеются идентичные, но противоположно ориентированные тандемы повторяющихся нуклеотидных последовательностей. Геномы вирусов оспы способны кодировать около 200 белков, из которых не менее 100 входят в структуру вириона. Однако функциональные особенности определены лишь у небольшого количества вирусных белков. Наиболее важными из них являются ферменты, участвующие в синтезе вирусных нуклеиновых кислот и структурных компонентов вирионов. Например, синтез ДНК-полимеразы, ДНК-лигазы, РНК-полимеразы, энзимов, связанных с кэппированием и полиаденилированием мРНК и тимидинкиназы.
Инфекционные вирусные частицы содержат системы транскрипции, которые могут выполнять in vitro синтез РНК, а также способны полиаденилировать, кэппировать и метилировать. В вирусных частицах содержится большое количество кодируемых вирусом энзимов и других биологически активных факторов.
Некоторые гены вирусов оспы кодируют белки, которые секретируются инфицированными клетками и вызывают ответ организма на инфекцию, в том числе и формирование иммунитета.
К таким вирокинам относятся гомологичный эпидермальный фактор роста, белок, снижающий активность комплемента, вирокины, обеспечивающие устойчивость к интерферону, и другие супрессоры иммунного ответа, подавляющие действие некоторых цитокинов организма хозяина.
Вирусы оспы обычно характеризуются узким спектром хозяев. Они передаются чаще респираторным путем и реже через поврежденную кожу. Вирусы оспы овец, свиней, птиц и миксоматоза передаются также через укус членистоногими. Вирусы оспы устойчивы в окружающей среде и могут сохраняться годами в высохших струпьях кожи или других вируссодержащих материалах.
Большинство вирусов оспы хорошо размножаются в культуре клеток. Исключение составляют парапоксвирусы, вирус оспы свиней и вирус контагиозного моллюска. Однако они, так же как и ортопоксвирусы, легко образуют оспины на хориоал-лантоисной оболочке куриных эмбрионов.
Образование вириона связано с вхождением ДНК внутрь незрелой сердцевинной структуры, которое затем завершается включением наружных покрывающих слоев. Репликация и сборка вирионов происходят в разных местах цитоплазмы в так называемых виропластах или вирусных фабриках. Вирионы выходят из клетки почкованием (оболочечные вирионы), или путем экзоцитоза, или при лизисе клеток (вирионы без оболочки). Большинство вирионов освобождаются при цитолизе и не имеют оболочки. Вирионы с оболочкой и без нее обладают инфекционностью, но первые, вероятно, играют более значительную роль в возникновении и распространении заболевания, а также в создании иммунитета.
В очищенном вирусе осповакцины (ВОВ) выявлены белки с молекулярной массой 10-250 кД. Многие из них сосредоточены в сердцевине вириона. Два структурных гликопротеина располагаются между оболочкой и сердцевиной. В оболочке ВОВ содержится около 10 белков, из которых иммунологически наиболее активны крупномолекулярные белки с молекулярной массой 58—32 кД (VP4c, VP6a, VP6b и VP7a). Белок 32 кД определяет круг хозяев и важен для репликации вируса.
В составе очищенного вируса оспы птиц обнаружено 29 полипептидов с молекулярной массой 14-138 кД. Наивысшей антигенной и иммуногенной активностью обладают полипептиды с молекулярной массой 35 и 37 кД. За индукцию вируснейтрализующих антител ВО ответственны антигены, расположенные на поверхности наружной оболочки вириона, и прежде всего белок 58 кД (VP4c), являющийся основным структурным компонентом трубочек (ворсинок). Антисыворотка к этому белку нейтрализовала инфекционность вируса и предотвращала образование синцития в культуре клеток. Этот белок ответственен за выработку иммунитета.
Внеклеточные вирионы покрыты дополнительной наружной оболочкой, отсутствующей у внутриклеточных вирионов. Она играет важную роль в индукции синтеза ВН-антител. Инфекционность ВОВ и ВО крупного рогатого скота, имеющих наружную оболочку, нейтрализовалась антисывороткой к имеющему эту оболочку ВОВ, но не нейтрализовалась антисывороткой к ВОВ, лишенному наружной оболочки.
За гемагглютинирующую активность ортопоксвирусов ответственны полипептид 85 кД и гликопротеид 41 кД. Во внеклеточном оболочечном ВОВ гемагглютинирующие свойства связаны с полипептидом 85 кД. Внутриклеточные вирионы (без дополнительной оболочки) практически не содержали этого полипептида. Неструктурный гемагглютинин формируется на цитоплазматических мембранах. С его образованием инфицированные клетки приобретают способность адсорбировать эритроциты. Вирусспецифические белки с молекулярной массой 32 и 37 кД, экспрессируемые на поверхности клеток, инфицированных ВОВ, делают их мишенями для специфических цитотоксических Т-лимфоцитов. МАТ-реактивные против каждого из пяти (54; 34; 32; 29 и 17—25 кД) белков наружного слоя поверхности вируса осповакцины нейтрализовали его инфекционность. В структуре полипептида 54 кД обнаружено два нейтрализующих эпитопа(Аи В). Анализ антигенных детерминант поверхностных полипептидов, проведенный с помощью МАТ, выявил в составе ортопоксвирусов, наряду с видоспецифическими, группоспецифические эпитопы.
Связывание ВОВ с нейтрализующими МАТ не препятствовало его прикреплению к клеткам-мишеням, но блокировало депротеинизацию вирионов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вирусы оспы. Виды вирусов оспы. Строение вирусов оспы.
Вирусы оспы (ВО) — самые крупные вирусы, содержащие ДНК, молекулярная масса которой больше, чем у любого другого вируса животных. Они широко распространены в природе, вызывают заболевания позвоночных и беспозвоночных с летальным исходом или в виде легко протекающей продолжительной инфекции с образованием доброкачественных опухолей. ВО передаются членистоногими при прямом контакте, аэрозольно или механически. ВО обычно имеют узкий круг хозяев, хотя некоторые из них представляют исключения, в том числе вирусы вакцины и оспы птиц. Вирусы оспы позвоночных включают восемь родов (орто-, пара-, ави-, капри-, лепори-, суи-, моллюсци- и ятапоксвирусы).
Разделение вирусов оспы позвоночных на роды проведено с учетом массы и структуры генома, размера и формы вириона, способности к генетической рекомбинации, антигенной связи и спектра патогенности.
Ортопоксвирусы. Типичный представитель - вирус осповакцины (ВОВ). Спектр естественных хозяев узкий, обычно ограничен одним видом животных: вирусы агглютинируют эритроциты цыплят; варьируют по вирулентности для естественных хозяев и лабораторных животных. Другие виды: вирусы оспы буйволов, верблюдов, крупного рогатого скота, мышей, обезьян и вирус натуральной оспы.
Парапоксвирусы. Типичный представитель — вирус контагиозного пустулёзного дерматита (эктимы) овец и коз (вирус орф). Представители рода серологически связаны между собой, но отличаются от представителей других родов; гемагглютинин не образуют. Другие виды: вирусы пустулёзного стоматита крупного рогатого скота, контагиозной эктимы серн и сыпи доярок.
Авипоксвирусы. Естественные хозяева — птицы; типичный представитель — вирус оспы кур. Члены рода серологически связаны между собой. Другие виды: вирусы оспы канареек, голубей, перепелов, воробьев, скворцов, индеек и оспы Юнко. Передаются членистоногими.
Каприпоксвирусы. Естественные хозяева — парнокопытные; типичный представитель — ВО мелких жвачных (овец и коз). В данный род входит также вирус кожной бугорчатки крупного рогатого скота.
Лепорипоксвирусы. Типичный представитель — вирус миксомы кроликов. Передается механически членистоногими. Другие виды: вирусы фибромы зайцев, кроликов (вирус Шоупа) и белок. Вирус злокачественной фибромы кроликов — летальный туморогенный поксвирус, возникший, по-видимому, вследствие рекомбинации между вирусами фибромы и миксомы кроликов.
Суипоксвирусы. Типичный представитель — вирус оспы свиней. В инфицированных клетках образуются несколько типов цитоплазматических включений и наблюдается вакуолизация ядра. Другие члены рода не определены.
Моллюсципоксвирусы. Типичный представитель — вирус контагиозного моллюска. Возможные члены рода — вирусы однокопытных и обезьян.
Ятапоксвирусы включают вирус опухолей обезьян Яба и вирус оспы Тана.
Вирусы оспы — наиболее крупные из всех вирусов животных. Под электронным микроскопом они выглядят как большие овальные (кирпичеобразные) частицы размером около 250-350x200-270 нм.
Орто-, ави-, лепорипоксвирусы более вытянуты, а вирус оспы свиней шире, чем другие ВО. Парапоксвирусы имеют овальную (коконообразную) форму и размер 260x160 нм.
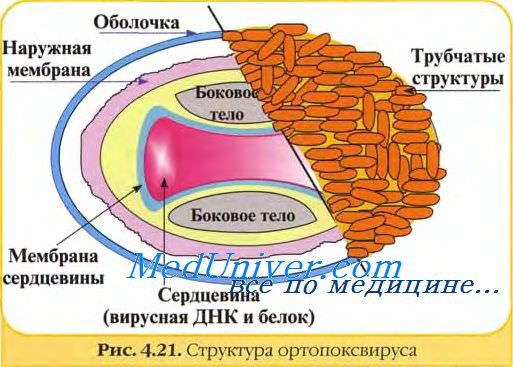
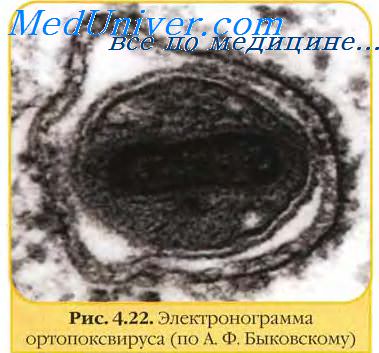
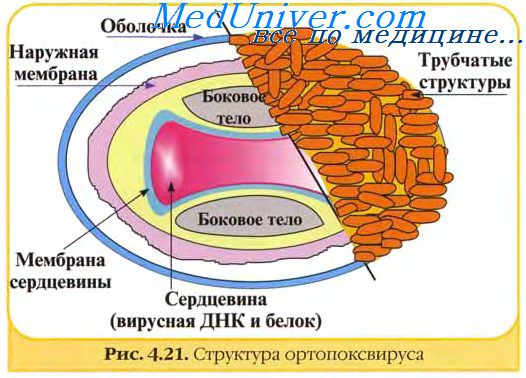
В структуре вирусов оспы различают три основных компонента: двояковогнутую сердцевину, овальные боковые тела и оболочку вириона. Сердцевину вириона составляют ДНК и связанные с нею белки. Сердцевина окружена гладкой мембраной (толщиной около 5 нм), снаружи покрытой слоем вертикально уложенных и плотно прилегающих друг к другу цилиндрических субъединиц (5x10 нм). Вогнутость сердцевины с обеих сторон занята овальными образованиями (неизвестной природы), называемыми боковыми телами. Они как бы сдавливают сердцевину, придавая ей форму двояковогнутого диска, имеющего на разрезе вид гантели.
Вирионы заключены в липопротеиновую супероболочку, которая имеет толщину 20—30 нм и содержит липиды клетки и вирусспецифические белки.
Вирионы большинства вирусов оспы окружены слоем беспорядочно расположенных трубчатых структур, придающих им характерный вид. Эти структуры состоят из сферических субъединиц диаметром около 5 нм. Субъединицы построены из молекул протеина или гликопротеина. В состав ворсинок длиной 20 нм покрывающих поверхность вируса осповакцины (ВОВ), входит белок с молекулярной массой 58 кД, относящийся к главным полипептидам вириона. Поверхность парапоксвирусов покрыта длинными нитеподобными трубочками, уложенными крест-накрест, напоминающими клубок пряжи.
Вирус осповакцины содержит белки, липиды и ДНК, которые соответственно составляют 90, 5 и 3,2% массы вириона (5х10~15 г). В вирусе оспы птиц около 1/3 массы составляют липиды.
Вирион образуется включением ДНК внутрь незрелых вирусоподобных частиц, которые затем созревают, покрываясь дополнительно наружными оболочками. Репликация и сборка вирионов происходят в разных местах цитоплазмы (в виропластах или вирусных фабриках), и вирионы освобождаются почкованием или при лизисе клеток.
Вирионы, освободившиеся из клетки почкованием до ее разрушения, покрыты оболочкой, которая содержит клеточные липиды и несколько вирусспецифических белков.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Оспа птиц (лат. – Variola avium; англ. – Pox; оспа-дифтерит) – контагиозная болезнь птиц отряда куриных, голубиных, воробьиных, характеризующаяся поражением эпителия кожи, дифтеритическим и катаральным воспалением слизистых оболочек ротовой полости и верхних дыхательных путей [3].
Возбудитель по номенклатуре ICTV 2009/2010 (МКТВ):
Виды: Canarypox virus, Fowlpox virus, Juncopox virus, Mynahpox virus, Pigeonpox virus, Psittacinepox virus, Quailpox virus, Sparrowpox virus, Starlingpox virus, Turkeypox virus [5].
Морфология и химический состав. Вирионы овальной или прямоугольной формы, размером в среднем 140—450 нм и со сложным типом симметрии. Состоят из нуклеоида, содержащего геном, боковых тел и наружной липопротеидной оболочки, образующей выступы. Геном представлен двуспиральном линейной ДНК [2].
Антигенная структура, антигенная активность, антигенные свойства. Вирионы вируса оспы содержат наружный протеидный антиген в составе поверхностных ворсинок (филаментов). Наружный антиген индуцирует образование антигемагглютинирующих вируснейтрализующих антител, которые обеспечивают развитие иммунитета. Внутренние полипептиды индуцируют комплементсвязывающие и преципитирующие антитела [2].
В составе очищенного вируса ОП обнаружено 29 полипептидов с молекулярной массой 14-138 кД. Наивысшей антигенной и иммуногенной активностью обладают полипептиды с молекулярной массой 35 и 37 кД. За индукцию ВНА вируса ОП ответственны АГ, расположенные на поверхности наружной оболочки вириона и прежде всего белок 58 кД, являющийся основным структурным компонентом трубочек (ворсинок). Антисыворотка к этому белку нейтрализует инфекционность вируса и предотвращает образование синцития в культуре клеток. Этот белок ответственен за выработку иммунитета. Выявлен 21 белок, кодирующий вирус оспы птиц. Эти белки можно разделить на пререпликативные и пострепликативные по аналогии с вирусом осповакцины. В первую группу входит белок 70 кД. В сыворотках крови больных и экспериментально зараженных кур с 4-7-го дня появляются ГА в титре 1:10 - 1:80 и достигающие к 22-30 дню титра 1:160 - 1:640, далее идет снижение титра. РЗГА специфична, ее можно использовать для диагностики оспы. У гипериммунизированных вирусом оспы кур выявляются специфические АТ-преципитины. Преципитирующая сыворотка при 4°С сохраняет активность до 300 дней. АТ в сыворотке крови кур после однократного введения им оспенного вируса реагируют только с полипептидами, молекулярная масса которых 37 и 35 кД, тогда как АТ после вторичной инокуляции курам вируса оспы реагируют еще с 6-ю другими структурными белками вируса. Иногда полевые вирулентные изоляты вызывают образование анти-ГА в высоком титре, но ВНА - в низком [4].
Устойчивость к физико-химическим факторам. Вирус чувствителен к высокой температуре. Высушивание и холод консервируют его. В оспенных корочках при температуре -15°С он выживает до 2 лет и более. В лиофилизированном виде при хранении под вакуумом оспенный материал остается вирулентным до 8 лет. Чувствителен к этиловому спирту, быстро погибает в гниющем субстрате [1].
Культивирование, экспериментальная инфекция. Для культивирования вируса используют куриные эмбрионы, у которых он поражает ХАО и обнаруживается в тканях и органах зародыша. Кроме того, применяют культуры клеток куриных фибробластов, где вирус вызывает округление, вакуолизацию и образование клеток с 5-7 ядрами. Вирус хорошо репродуцируется в перевиваемой линии клеток QT-35.
Экспериментальная инфекция легко воспроизводится на курах и цыплятах. У кур материал втирают в перьевые фолликулы, скарифицированную кожу гребня, бородки и сережек, где развивается доброкачественный фолликулит. Некоторыми штаммами можно вызвать заражение ворон, уток, ястребов и канареек [1].
Библиографический список:
Барышников П.И. Ветеринарная вирусология [Текст]: учебное пособие / П.И. Барышников. Барнаул: Изд-во АГАУ, 2006.– 14 с.
Р.В. Белоусова, Э.А. Преображенская, И.В. Третьякова. Ветеринарная вирусология. Под ред. проф. Р. В. Белоусовой. — М.: КолосС, 2007. — 424 с.
Бессарабов, Б.Ф. Инфекционные болезни животных [Текст] : учебник / Б. Ф. Бессарабов, А. А. Вашутин, Е. С. Воронин и др.; Под ред. А. А. Сидорчука. — М.: КолосС, 2007. — 448 с.
Сюрин, В. Н. Ветеринарная вирусология [Текст] : учебник / В. Н. Сюрин, Р. В. Белоусова, Н. В. Фомина. – М. : Агропромиздат, 2003. – 439 с.
Читайте также: