
Рецепторы клеток для вирусов
Обновлено: 18.04.2024
Мы кратко рассмотрели структуру вирионов гриппа и процесс кодирования вирусными РНК одного и более белков. Теперь обратимся к размножению вирусов гриппа.
Вирусы – это исключительно внутриклеточные паразиты, они не могут воспроизводиться вне клетки. Появление новых инфекционных частиц должно протекать внутри клетки. Войдя в клетки, вирусы начинают паразитировать в структуре хозяина для воспроизводства вирусного потомства. Все события, происходящие в инфицированной вирусом клетке, называются инфекционным циклом, или вирусной репликацией. Вирусологи искусственно разделяют инфекционный цикл на несколько стадий, чтобы проще было его изучить. Эти стадии включают прикрепление и вхождение вириона, трансляцию мРНК в белок, репликацию генома (создание большего количества РНК или ДНК), скопление новых частиц и выход частиц из клетки. Мы рассмотрим каждую из названных стадий, а затем обсудим, как вирус гриппа инфицирует нас и вызывает болезнь.
Остановимся на первой стадии – прикреплении вириона к клетке. Возьмем обычную клетку, с которой, я уверен, знаком каждый. Но не будет лишним повториться.
Рисунок 10.
Слева показан клеточный белок, прикрепленный к цитоплазматической мембране. Снизу изображена внутренняя часть клетки – цитоплазма. Часть белка пересекает мембрану, и также видны части цитоплазматической и внеклеточной сторон. Сферы – это сахара, прикрепленные ко множеству белков (белок + сахар = гликопротеин). Сиаловая кислота всегда является последним сахаром в цепи, которая прикрепляется к белку. Справа показана химическая структура сиаловой кислоты; следующий сахар (справа) – галактоза. Вирионы гриппа прикрепляются к клеткам, когда HA захватывает очень маленькую сиаловую кислоту.
Сахар в сравнении с HA в действительности очень мал – он подходит к небольшому кармашку на острие шипа. На рис. 11 изображена молекулярная модель, демонстрирующая, как HA прикрепляется к аналогу сиаловой кислоты. Шаровидный конец HA показан в верхней части рис. 11. Маленькие красные и белые сферы показывают места будущего прикрепления сиаловой кислоты – кармашки на верхнем конце HA.
Рисунок 13.
Процесс вхождения вируса гриппа в клетки – наиболее понятный из всех известных механизмов проникновения вирусов. После прикрепления вириона к сиаловой кислоте, имеющей рецепторы на поверхности клетки, вирус-рецепторный комплекс входит в клетки посредством эндоцитоза – процесса, в ходе которого клетки обычно берут молекулы из внеклеточной жидкости. Пока эндосомальные везикулы, содержащие частицы вируса, движутся к ядру клетки, их pH понижается. Это изменение в pH завершает клеточный канал, закачивающий протоны (H+) в везикулу. Когда эндосомальная pH достигает 5,0, вирусный белок HA подвергается конформационной перестройке. Это изменение подвергает пептид слияния на HA короткой гидрофобной последовательности, вставляющей его в эндосомальную мембрану, которая от этого сливается с вирусной оболочкой. Когда это происходит, вирусные РНК проникают в цитоплазму. Затем они транспортируются в ядро клетки, где начинается размножение.
В вирионе гриппа вирусные РНК не оголены, поскольку к ним прикреплены вирусные белки, включая белок M1. Этот белок образует оболочку, лежащую в основе липидной мембраны вириона. К сожалению, если вирусные РНК прикрепляются к белку M1, когда выходят из вириона, они не могут достичь ядра. Для решения этой проблемы вирион гриппа имеет в своей мембране несколько копий белка, называемого M2. Этот вирусный белок образует канал в мембране, который активно закачивает протоны из эндосомы во внутреннюю часть вириона. Данные протоны понижают pH внутри вириона, освобождая вирусные РНК от M1. В этом случае РНК могут войти в ядро.
Ионный канал M2, являющийся мишенью для противовирусных адамантанов, показан на рис. 14. Эти образования забивают канал и не позволяют протонам закачиваться в вирион. В присутствии адамантанов вирусным РНК нужно прикрепляться к M1, отчего они не могут достичь ядра. Поэтому вирусная репликация подавляется. Устойчивость к адамантанам возможна при изменении в аминокислотах, прокладывающих канал M2. Такие изменения не дают лекарству забить канал.
Рисунок 15.
На рис. 15 сферический конец белка HA, который прикрепляется к рецепторам клетки, изображен сверху, а вирусная мембрана – снизу. Для наглядности помечен только сайт расщепления HA. Нерасщепленная форма белка называется HA0; после расщепления клеточным ферментом образуются два белка, называемые HA1 (синий) и HA2 (красный). Две субъединицы остаются вместе на поверхности вирусной частицы. Новая амино(N)-концевая область HA2, образовавшегося при расщеплении, содержит последовательность гидрофобных аминокислот, называемых пептидом слияния. Во время вхождения вируса гриппа в клетки пептид слияния вставляется в эндосомальную мембрану и вызывает слияние вирусной и клеточной оболочек. Следовательно, вирусные РНК гриппа могут войти в цитоплазму. Процесс слияния описан в предыдущем параграфе.
Если белок HA не расщеплен, чтобы образовать HA1 и HA2, слияния не происходит. Поэтому вирусы гриппа с нерасщепленным HA не заразны. Расщепление вирусного HA происходит после того, как вновь синтезированные вирионы выходят из клетки. Вирусы гриппа эффективно размножаются в яйцах из-за присутствия протеазы в аллантоисной жидкости, способной расщеплять HA. Однако репликация многих штаммов вируса гриппа в культурах клеток требует добавления в среду соответствующей протеазы (как привило, трипсина).
У людей репликация вируса гриппа ограничена дыхательными путями, потому что это единственный участок, где производится протеаза, расщепляющая HA. Однако белок HA высокопатогенных штаммов H5 и H7 вируса птичьего гриппа может расщепляться протеазами, производимыми во многих различных тканях. В результате эти вирусы способны размножаться во многих органах птиц, включая селезенку, печень, легкие, почки и мозг. Это свойство объясняет способность штаммов H5N1 вируса птичьего гриппа размножаться вне дыхательных путей человека.
Подобно белкам HA высокопатогенных вирусов H5 и H7, HA штамма вируса гриппа 1918 г. может также расщепляться повсеместно присутствующими клеточными протеазами. Следовательно, вирус может размножаться в культурах клеток без добавления трипсина.
Белки HA H5 и H7 имеют множество основных аминокислотных остатков на сайте расщепления HA1-HA2, которые делают возможным расщепление посредством широко представленных протеаз. Но HA вируса гриппа H1 1918 г. не обладал этим свойством. Также N1 1918 г. не был в состоянии привлечь протеазы, которые расщепляли бы HA, т. е. не было механизма, позволяющего штамму A/WSN/33 вируса гриппа размножаться в клетках без трипсина. Понимание того, как белок HA вируса H1 1918 г. мог быть расщеплен протеазами – основной в понимании высокой патогенности данного штамма.
Рекомендуемая литература.
Chaipan, C., Kobasa, D., Bertram, S., Glowacka, I., Steffen, I., Solomon Tsegaye, T., Takeda, M., Bugge, T., Kim, S., Park, Y., Marzi, A., &Pohlmann, S. (2009). Proteolytic Activation of the 1918 Influenza Virus Hemagglutinin Journal of Virology, 83 (7), 3200-3211 DOI:10.1128/JVI.02205-08.
Рецепторы клеток в условиях вирусной инфекции.
Проблема адренергического контроля, да и вообще проблема передачи регуляторного сигнала внутрь клетки в условиях вирусной инфекции настолько сложна и интересна, насколько не изучена. Ведь даже выяснение роли липидов во взаимодействии вируса гриппа с клеткой-хозяином может пролить свет на понимание характера и причин изменений липидного матрикса мембран, опосредующего трансдукцию разнообразных регуляторных влияний в условиях вирусной инфекции.
Липиды вируса образуют бислой, присущий всем биологическим мембранам. Генез вирусной мембраны, представляющей дифференцированный сегмент плазматической мембраны клетки-хозяина, обусловливает высокую степень сходства этих мембран по липидному составу. Результаты изучения вируса гриппа и других оболочечных вирусов позволяют считать, что их липидный состав отражает состав доменов плазматической мембраны в момент сборки на них вирионов. Как известно, оболочки вирусов содержат значительно больше холестерина, чем мембраны клетки-хозяина.
Естественно, что в процессе инфекции, вызванной вирусом гриппа, липидный метаболизм клетки-мишени может модифицироваться. Если учесть, что эритроциты (наряду с лейкоцитами и макрофагами) рассматриваются как реальные вирусоносители не только при вирусемии в острый период, но и на этапе персистенции, а возможность прикрепления к эритроцитам и даже визуализация этого процесса убедительно продемонстрирована, то можно ожидать, что вирус способен индуцировать изменение микровязкости в эритроцитарных мембранах.

Эта модификация может быть обусловлена, в частности, обогащением вирусной мембраны холестерином. Кстати, только адсорбция на поверхности клеток оболочечных вирусов (в том числе вируса гриппа) сопровождается быстрым и значительным повышением текучести мембранных липидов инфицированных клеток.
Изменения микровязкости липидного матрикса при кратковременном и длительном воздействии на клетки-мишени, несомненно, могут модифицировать и рецепторно-мембранные события при опосредовании связывания самых различных лигандов (токсинов, медиаторов, гормонов, лектинов, антител). При этом для адренергической активации могут быть характерны так называемые парадоксальные ответы вследствие инверсии проведения сигнала.
Весьма любопытны данные о том, что неполный вирус гриппа по сравнению со стандартным вирусом характеризуется более низким содержанием холестерина и повышенной текучестью вирусной мембраны, что определяет низкую инфекционность неполного вируса, опосредующего развитие персистентной инфекции.
Подытоживая раздел об особенностях мембрано-рецепторных изменений при остром бактериальном и вирусном воспалении в бронхолегочной системе, необходимо еще раз подчеркнуть фрагментарность знаний по данной проблеме. Судя же по имеющимся данным, можно сделать вывод о том, что острая бактериальная и вирусная инфекция могут вызывать изменения конечного эффекторного механизма в адренореактивной системе, хотя конкретные звенья этой рецепторной системы (количество и аффинитет адренорецепторов, свойства сопрягающих N-белков, аденилатциклазы) не изучены.
По-видимому, существует обратимость изменений в мембрано-рецепторном комплексе, отражающая фазу воспаления.
И, наконец, - при вирусной инфекции наблюдаются наиболее выраженные изменения МРК, приводящие к формированию так называемых парадоксальных или извращенных конечных эффектов, а также к нарушениям негативного контроля в рецепторных системах.
- Вернуться в оглавление раздела "Пульмонология."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Типы инфицирования клеток вирусами. Репродуктивный цикл вирусов. Основные этапы репродукции вирусов. Адсорбция вириона к клетке.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Репродуктивный цикл вирусов
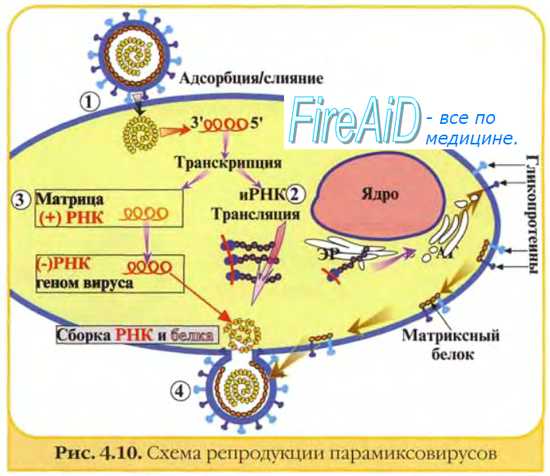
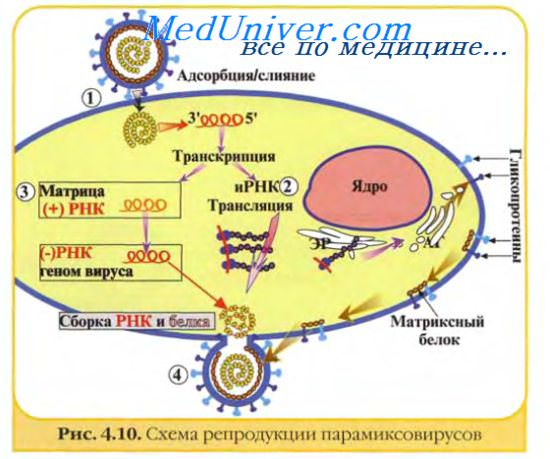
Изображённые на рис. 2-3 этапы репродукции (от адсорбции вирионов до высвобождения дочерней популяции) происходят при продуктивном взаимодействии вируса с клеткой.
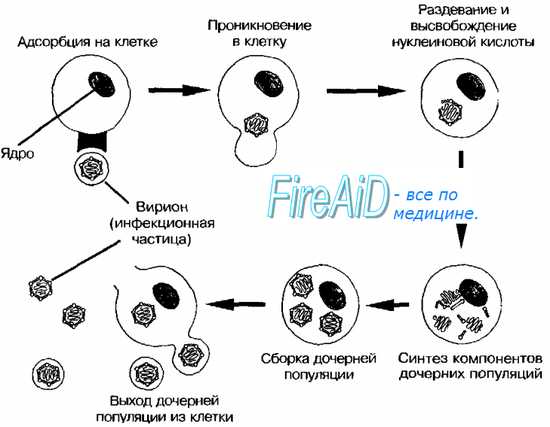
Рис. 2-3. Основные этапы репродукции вирусов.
Адсорбция вириона к клетке
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Цикл размножения вирусов. Прикрепление или адсорбция вируса.
Цикл репродукции вирусов при продуктивной инфекции представляет собой специфическую последовательность событий, приводящих к образованию нового потомства вирионов. Его можно разделить на несколько стадий: адсорбция вирионов на поверхности клетки, проникновение вируса или вирусного нуклеопротеида внутрь клетки, депротеинизация генома, синтез вирусных компонентов, формирование и выход из клетки зрелых вирионов. Иногда не все стадии протекают последовательно и полностью даже при продуктивной инфекции. При заражении вирусной нуклеиновой кислотой отсутствует стадия депротеинизации. Цикл репродукции вируса может прерываться на какой-либо промежуточной стадии, и в этом случае инфекционные вирионы не образуются (абортивная инфекция).
Классическое изучение кривой роста вируса в одном цикле репродукции предполагает одновременное инфицирование клеток культуры с использованием высокой множественности заражения (М=10—100) и определение накопления вируса через определенные промежутки времени.
Прикрепление (адсорбция) вирионов к поверхности клетки - первая стадия вирусной инфекции. Для того, чтобы началась инфекция, вирионы должны быть способны прикрепляться к клетке. Безусловно, этот процесс в организме является более сложным, чем в культуре клеток.
Детальное изучение процесса адсорбции вирусов показало, что он состоит из двух быстро следующих друг за другом периодов — обратимого и необратимого. В период обратимой адсорбции вирус можно удалить с поверхности клетки при обработке версеном, хемотрипсином и другими химическими веществами. При необратимой адсорбции вирус удалить с поверхности клетки не удается.
Первичный контакт с клеткой происходит в результате случайных столкновений вирионов с поверхностью клетки. Количество стабильных прикреплений вириона к клетке во много раз меньше количества случайных столкновений.
В основе прикрепления вируса к клетке лежат два механизма — неспецифический и специфический. Первый из них определяется силами электростатического взаимодействия, возникающими между разноименно заряженными группами, расположенными на поверхности клетки и вируса. В этом процессе, прежде всего, могут участвовать положительно заряженные аминные группы вирусного белка и отрицательно заряженные группы клеточной поверхности. В пользу электростатического взаимодействия между клеткой и вирусом свидетельствуют данные о зависимости присоединения вирионов к клеткам от ионной силы раствора и его рН, наличия в среде одно- и двухвалентных катионов и присутствия отрицательно заряженных групп на поверхности клетки.
Наиболее важным механизмом прикрепления вируса является специфическое взаимодействие рецепторов вируса с комплементарными рецепторами клетки.
Прикрепление происходит за счет связи между вирионными прикрепительными белками на поверхности вирионов и рецепторами на плазматической мембране клеток.
На поверхности вирионов многих семейств вирусов (все РНК-содержащие оболочечные вирусы и аденовирусы) имеются пепломеры (выступы и шипы) длиной от 10 до 30 нм, которые принимают непосредственное участие в прикреплении вируса к клетке. Отщепление пепломеров с помощью протеолитических ферментов приводит к потере вирионами способности адсорбироваться на клетках. На поверхности вирионов, не имеющих пепломеров, находятся участки связывания с клеточной поверхностью, которые отличаются сложным строением и, вероятно, состоят из нескольких полипептидов.
Спектр чувствительности клеток к вирусам в значительной мере определяется наличием соответствующих рецепторов. Рецепторный барьер может быть преодолен при заражении инфекционной нуклеиновой кислотой.
На поверхности клетки может находиться большое количество различных видов рецепторов. Полагают, что каждая клетка может адсорбировать от десятков до нескольких тысяч вирионов. На поверхности одной клетки находится около 10000 рецепторов для полиовируса. Каждый рецептор специфичен для одного или нескольких вирусов, как родственных между собой, так и относящихся к различным таксономическим группам. Например, аденовирус типа 2 и вирус Коксаки ВЗ связываются с одними и теми же клеточными рецепторами.
У герпесвирусов прикрепительными белками могут служить гликопротеины оболочки, которые соединяются с некоторыми рецепторами клетки. Клеточным рецептором для многих ортомиксовирусов являются концевые сиаловые кислоты олигосахаридов, входящих в структуру гликопротеринов или гликолипидов, экспонированных на поверхности клетки. Рецепторами для многих риновирусов являются иммуноглобулины. У вируса ВИЧ в прикреплении первоначально участвуют молекулы СД4, расположенные на поверхности клеток, особенно макрофагов и Т-хелперных лимфоцитов. Подобные взаимодействия рецептор-лиганд свойственны другим лентивирусным инфекциям.
Рецепторы для одних вирусов имеются на клетках нескольких или многих тканей, тогда как для других вирусов они обнаружены на клетках одного вида тканей или в один из периодов онтогенеза (новорожденные или взрослые животные).
Рецепторы клеток способны к регенерации после удаления их протеолитическими ферментами.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Обзор
Автор
Редакторы
Обратите внимание!
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science.
Эволюция и происхождение вирусов
В 2007 году сотрудники биологического факультета МГУ Л. Нефедова и А. Ким описали, как мог появиться один из видов вирусов — ретровирусы. Они провели сравнительный анализ геномов дрозофилы D. melanogaster и ее эндосимбионта (микроорганизма, живущего внутри дрозофилы) — бактерии Wolbachia pipientis. Полученные данные показали, что эндогенные ретровирусы группы gypsy могли произойти от мобильных элементов генома — ретротранспозонов. Причиной этому стало появление у ретротранспозонов одного нового гена — env, — который и превратил их в вирусы. Этот ген позволяет вирусам передаваться горизонтально, от клетки к клетке и от носителя к носителю, чего ретротранспозоны делать не могли. Именно так, как показал анализ, ретровирус gypsy передался из генома дрозофилы ее симбионту — вольбахии [7]. Это открытие упомянуто здесь не случайно. Оно нам понадобится для того, чтобы понять, чем вызваны трудности борьбы с вирусами.
Из давних письменных источников, оставленных историком Фукидидом и знахарем Галеном, нам известно о первых вирусных эпидемиях, возникших в Древней Греции в 430 году до н.э. и в Риме в 166 году. Часть вирусологов предполагает, что в Риме могла произойти первая зафиксированная в источниках эпидемия оспы. Тогда от неизвестного смертоносного вируса по всей Римской империи погибло несколько миллионов человек [8]. И с того времени европейский континент уже регулярно подвергался опустошающим нашествиям всевозможных эпидемий — в первую очередь, чумы, холеры и натуральной оспы. Эпидемии внезапно приходили одна за другой вместе с перемещавшимися на дальние расстояния людьми и опустошали целые города. И так же внезапно прекращались, ничем не проявляя себя сотни лет.
Вирус натуральной оспы стал первым инфекционным носителем, который представлял действительную угрозу для человечества и от которого погибало большое количество людей. Свирепствовавшая в средние века оспа буквально выкашивала целые города, оставляя после себя огромные кладбища погибших. В 2007 году в журнале Национальной академии наук США (PNAS) вышла работа группы американских ученых — И. Дэймона и его коллег, — которым на основе геномного анализа удалось установить предположительное время возникновения вируса натуральной оспы: более 16 тысяч лет назад. Интересно, что в этой же статье ученые недоумевают по поводу своего открытия: как так случилось, что, несмотря на древний возраст вируса, эпидемии оспы не упоминаются в Библии, а также в книгах древних римлян и греков [9]?
Строение вирусов и иммунный ответ организма

Рисунок 1. Первооткрыватель вирусов Д.И. Ивановский (1864–1920) (слева) и английский врач Эдвард Дженнер (справа).

Почти все известные науке вирусы имеют свою специфическую мишень в живом организме — определенный рецептор на поверхности клетки, к которому и прикрепляется вирус. Этот вирусный механизм и предопределяет, какие именно клетки пострадают от инфекции. К примеру, вирус полиомиелита может прикрепляться лишь к нейронам и потому поражает именно их, в то время как вирусы гепатита поражают только клетки печени. Некоторые вирусы — например, вирус гриппа А-типа и риновирус — прикрепляются к рецепторам гликофорин А и ICAM-1, которые характерны для нескольких видов клеток. Вирус иммунодефицита избирает в качестве мишеней целый ряд клеток: в первую очередь, клетки иммунной системы (Т-хелперы, макрофаги), а также эозинофилы, тимоциты, дендритные клетки, астроциты и другие, несущие на своей мембране специфический рецептор СD-4 и CXCR4-корецептор [13–15].

Одновременно с этим в организме реализуется еще один, молекулярный, защитный механизм: пораженные вирусом клетки начинают производить специальные белки — интерфероны, — о которых многие слышали в связи с гриппозной инфекцией. Существует три основных вида интерферонов. Синтез интерферона-альфа (ИФ-α) стимулируют лейкоциты. Он участвует в борьбе с вирусами и обладает противоопухолевым действием. Интерферон-бета (ИФ-β) производят клетки соединительной ткани, фибробласты. Он обладает таким же действием, как и ИФ-α, только с уклоном в противоопухолевый эффект. Интерферон-гамма (ИФ-γ) синтезируют Т-клетки (Т-хелперы и (СD8+) Т-лимфоциты), что придает ему свойства иммуномодулятора, усиливающего или ослабляющего иммунитет. Как именно интерфероны борются с вирусами? Они могут, в частности, блокировать работу чужеродных нуклеиновых кислот, не давая вирусу возможности реплицироваться (размножаться).

Причины поражений в борьбе с ВИЧ
Тем не менее нельзя сказать, что ничего не делается в борьбе с ВИЧ и нет никаких подвижек в этом вопросе. Сегодня уже определены перспективные направления в исследованиях, главные из которых: использование антисмысловых молекул (антисмысловых РНК), РНК-интерференция, аптамерная и химерная технологии [12]. Но пока эти антивирусные методы — дело научных институтов, а не широкой клинической практики*. И потому более миллиона человек, по официальным данным ВОЗ, погибают ежегодно от причин, связанных с ВИЧ и СПИДом.

Подобный вирусный механизм характерен не только для ВИЧ. Он описан и при инфицировании некоторыми другими опасными вирусами: такими, как вирусы Денге и Эбола. Но при ВИЧ антителозависимое усиление инфекции сопровождается еще несколькими факторами, делая его опасным и почти неуязвимым. Так, в 1991 году американские клеточные биологи из Мэриленда (Дж. Гудсмит с коллегами), изучая иммунный ответ на ВИЧ-вакцину, обнаружили так называемый феномен антигенного импринтинга [23]. Он был описан еще в далеком 1953 году при изучении вируса гриппа. Оказалось, что иммунная система запоминает самый первый вариант вируса ВИЧ и вырабатывает к нему специфические антитела. Когда вирус видоизменяется в результате точечных мутаций, а это происходит часто и быстро, иммунная система почему-то не реагирует на эти изменения, продолжая производить антитела к самому первому варианту вируса. Именно этот феномен, как считает ряд ученых, стоит препятствием перед созданием эффективной вакцины против ВИЧ.

Открытие биологов из МГУ — Нефёдовой и Кима, — о котором упоминалось в самом начале, также говорит в пользу этой, эволюционной, версии.

Сегодня не только ВИЧ представляет опасность для человечества, хотя он, конечно, самый главный наш вирусный враг. Так сложилось, что СМИ уделяют внимание, в основном, молниеносным инфекциям, вроде атипичной пневмонии или МЕRS, которыми быстро заражается сравнительно большое количество людей (и немало гибнет). Из-за этого в тени остаются медленно текущие инфекции, которые сегодня гораздо опаснее и коварнее коронавирусов* и даже вируса Эбола. К примеру, мало кто знает о мировой эпидемии гепатита С, вирус которого был открыт в 1989 году**. А ведь по всему миру сейчас насчитывается 150 млн человек — носителей вируса гепатита С! И, по данным ВОЗ, каждый год от этой инфекции умирает 350-500 тысяч человек [33]. Для сравнения — от лихорадки Эбола в 2014-2015 гг. (на состояние по июнь 2015 г.) погибли 11 184 человека [34].
* — Коронавирусы — РНК-содержащие вирусы, поверхность которых покрыта булавовидными отростками, придающими им форму короны. Коронавирусы поражают альвеолярный эпителий (выстилку легочных альвеол), повышая проницаемость клеток, что приводит к нарушению водно-электролитного баланса и развитию пневмонии.

Рисунок 8. Электронная микрофотография воссозданного вируса H1N1, вызвавшего эпидемию в 1918 г. Рисунок с сайта phil.cdc.gov.
Почему же вдруг сложилась такая ситуация, что буквально каждый год появляются новые, всё более опасные формы вирусов? По мнению ученых, главные причины — это сомкнутость популяции, когда происходит тесный контакт людей при их большом количестве, и снижение иммунитета вследствие загрязнения среды обитания и стрессов. Научный и технический прогресс создал такие возможности и средства передвижения, что носитель опасной инфекции уже через несколько суток может добраться с одного континента на другой, преодолев тысячи километров.
Читайте также: