
Рнк зависимая днк полимераза вируса
Обновлено: 25.04.2024
ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
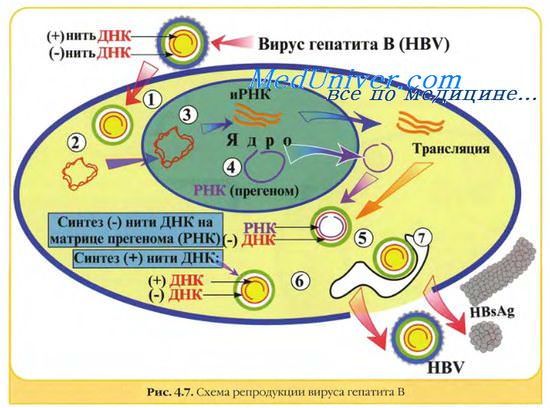
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Новость
Репарация ДНК: процесс, необходимый для жизни клетки
Автор
Редакторы
РНК выступает в роли матрицы для синтеза ДНК при обратной транскрипции ретровирусов (например, ВИЧ), ретротранспозонов и при поддержании длины теломер (повторяющихся последовательностей ДНК на концах хромосом, необходимых для их стабильности). Никаких указаний на прямое участие РНК как матрицы в репарации хромосом, включая залечивание двухцепочечных разрывов, получено не было, несмотря на то, что РНК присутствует в ядре в большом количестве. В большинстве организмов ДЦР устраняются за счёт гомологичной рекомбинации, или же за счёт негомологичного соединения концов разрыва.
Было обнаружено, что РНК косвенно может участвовать в репарации (негомологичном соединении концов), являясь матрицей для синтеза одноцепочечной ДНК-копии (кДНК), с последующим образованием комплиментарной цепи ДНК.
РНК также участвует в гомологичной рекомбинации в клетках почкующихся дрожжей (Saccharomyces cerevisiae), в которых рекомбинация проходит очень легко. И там РНК действует не напрямую, а посредством своей кДНК, синтезируемой в цитоплазме обратной транскриптазой, ферментом, поставляемым ретротранспозоном Ty.
Образование духцепочечных молекул (так называемых гетеродуплексов) одноцепочечными фрагментами ДНК и РНК возможно как in vitro, так и in vivo, однако прямой гомологичный обмен генетической информации между молекулами РНК и ДНК известен не был.
Авторы статьи, досрочно опубликованной на сайте журнала Nature, утверждают, что РНК может служить матрицей для синтеза ДНК во время репараци ДЦР хромосомной ДНК дрожжей [1]. Репарация осуществляется рибоолигонуклеотидами (одноцепочечными короткими фрагментами РНК), комплиментарными разорванным концам ДНК. Принимая во внимание этот факт и то, что ДНК-полимеразы α и δ, участвующие в репликации дрожжевой ДНК, могут копировать короткие РНК-матрицы in vitro, можно говорить о возможности переноса генетической информации молекулами РНК in vivo за счёт прямого взаимодействия с гомологичными последовательностями хромосомной ДНК.
Открытие синтеза ДНК in vitro и in vivo полимеразами α и δ на матрице РНК-содержащих олигониклеотидов существенно в ситуациях, когда фрагменты РНК оказывается в составе цепи ДНК in vivo, как, например, в случае с митохондриальным геномом млекопитающих. Включение рибонуклеотидных остатков в состав ДНК может происходить в процессе нормального метаболизма ДНК: некоторые ДНК полимеразы могут использовать в качестве субстрата рибонуклеотиды in vitro, a ДНК-лигаза I (фермент, сшивающий фрагменты ДНК) может пришивать к ДНК также и остатки рибонуклеотидов в процессе созревания фрагментов Оказаки in vitro.
Способность РНК переносить генетическую информацию на гомологичные молекулы хромосомных ДНК открывает новые возможности для направленного изменения генов (gene targeting), учитывая легкость амплификации РНК внутри клетки. Более того, РНК-матрицы, участвующие в репарации ДНК, могут вносить свой вклад в процессы эволюции генома и поддержания его стабильности.

Обзор
здесь и далее рисунки Андрея Занкевича
Автор
Редактор

Генеральный партнер конкурса — ежегодная биотехнологическая конференция BiotechClub, организованная международной инновационной биотехнологической компанией BIOCAD.

Спонсор конкурса — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Напомним, что РНК (рибонуклеиновая кислота) — это такая молекула, представляющая собой одну цепочку нуклеотидов. В составе каждого нуклеотида присутствует остаток моносахарида рибозы. На сегодняшний день известно множество разных типов РНК, которые выполняют совершенно разные функции: от кодирования клеточных белков (мРНК) до противовирусной защиты (некоторые микроРНК) [1–5]. РНК, входящие в состав многих вирусов, могут иметь ряд оригинальных функций, таких как регуляция времени экспрессии различных вирусных генов путем изменения пространственной организации цепи РНК или привлечение клеточных белковых комплексов.
Но бывает ли такое, что в пределах одной молекулы РНК одна ее часть, кодирующая некоторый белковый продукт, имеет положительную полярность, в то время как другая часть цепи представлена участком отрицательной полярности, кодирующим другой белок? Могла ли такая молекула возникнуть в процессе эволюции живых форм?
Ответ — да! И для того, чтобы разобраться, как функционируют такие молекулы, нам предстоит погрузиться в таинственный мир вирусов.
Давным-давно, в далекой-далекой галактике.
Как известно, все формы жизни обладают определенной наследственностью, которая определяет степень генетической идентичности живых объектов. В качестве молекул, ответственных за поддержание такой наследственной идентичности, выступают нуклеиновые кислоты (ДНК и РНК). Клеточные формы жизни для хранения и передачи информации используют только один тип нуклеиновых кислот — дезоксирибонуклеиновую кислоту (ДНК), в то время как вирусы в качестве основной молекулы наследственности могут использовать либо ДНК, либо РНК.
В случае ДНК-содержащих вирусов реализуется, как правило, Центральная догма молекулярной биологии в классическом виде: попав в клетку, ДНК удваивается в процессе репликации вирусного генома, на матрице ДНК в ходе транскрипции синтезируются мРНК, которые затем прочитываются рибосомой, синтезирующей по ним вирусные белки, то есть осуществляется трансляция. Далее вирусные белки ассоциируются с ДНК-геномом вируса в вирусную частицу (вирион), которая способна заражать новые клетки.
Очевидно, что РНК-содержащие вирусы используют иные стратегии размножения и, следовательно, реализации своего генома. Непривычные для большинства биологов молекулярные механизмы, которые используются такими вирусами, вероятно, унаследованы от далеких предков из того самого РНК-мира.
Размножение РНК-вирусов подразумевает использование разных типов РНК:
- геномная РНК находится внутри вириона, в зависимости от конкретного вируса, она может быть представлена (+)РНК, (–)РНК, (±)РНК, либо двухцепочечной РНК;
- комплементарная геномной РНК антигеномная РНК образуется в процессе репликации вирусов с одноцепочечным РНК-геномом и обладает полярностью, противоположной геномной РНК;
- субгеномная РНК (вирусная мРНК) имеет (+)полярность и является продуктом транскрипции геномной или антигеномной РНК. Как и подобает мРНК, субгеномная РНК участвует в процессе трансляционного синтеза белка.
Немного истории
Первым найденным РНК-вирусом стал бактериофаг f2, инфицирующий бактерию кишечную палочку (Escherichia coli) [7]. Выделенная геномная РНК фага f2 имела свойства мРНК, то есть она распознавалась рибосомой и могла транслироваться. На родственном РНК-бактериофаге Qβ была изучена РНК-зависимая РНК-полимераза (RdRp), которая, как оказалось, может синтезировать цепь РНК на матрице другой цепи РНК, то есть осуществлять репликацию вирусного РНК-генома! В ходе работы с РНК-бактериофагами f2 и его родственником Qβ были получены общие представления о биологии таких РНК-вирусов [8].
После РНК-бактериофагов были найдены (+)РНК-вирусы животных, такие, как вирус полиомиелита [9], [10], представитель группы пикорнавирусов. Подобные вирусы не содержат репликативных белков в составе вирусной частицы (вириона).
Встречаются вирусы, геном которых представлен двухцепочечной РНК. Как и в случае (–)РНК-вирусов, у дцРНК-вирусов во время репликации в клетке первым делом происходит синтез (+)цепи. Вирусные частицы этой группы также включают в свой состав RdRp.
Вирусы с двусмысленным РНК-геномом из семейства Bunyaviridae

Рисунок 1. Схематичное изображение структуры вириона флебовирусов
В семействе Bunyaviridae роды Phlebovirus, Tospovirus и Tenuivirus являются вирусами с двусмысленным РНК-геномом и, в отличие от остальных представителей семейства, имеют чуть более длинный S-сегмент генома (РНК S) (±)полярности. Род Tospovirus имеет вдобавок (+)участок на РНК M, который делает и эту РНК амбисенсной.
Флебовирусы
Вирусы рода Phlebovirus выделяют практически по всему миру и относят к нетаксономической группе арбовирусов, распространяющихся в членистоногих переносчиках и в позвоночных, на которых питаются переносчики. Члены этого рода переносятся кровососущими членистоногими. Инфекции не обходят стороной человека: вирусы сицилийской и неаполитанской москитных лихорадок широко распространены по территории Средиземноморья [15]. Среди симптомов таких инфекций — продолжительная сильная лихорадка, тошнота, рвота, диарея и головные боли. Вирус Тосканы, также переносимый москитами, обладает способностью проникать в нервную ткань и, вдобавок к вышеперечисленным симптомам, вызывает асептический менингит и менингоэнцефалит. Флебовирусы, переносимые клещами, например, вирус Бханджа, вирус тяжелой лихорадки с синдромом тромбоцитопении, или вирус Хартленд, вызывают серьезные вспышки инфекций среди людей [16].
Эти вирусы получили свое имя от латинского названия москитов (Phlebotominae), которые являются их основными переносчиками. Вирионы флебовирусов имеют диаметр 100-125 нанометров. Внутри вириона находятся три вирусных рибонуклеопротеина (вРНП), содержащих геномные сегменты, однако для вируса лихорадки долины Рифт (RVFV) было показано [17], что вирионы также могут содержать ещё три дополнительных вРНП, образованных цепочками антигеномных РНК, комплементарных геномным вирусным РНК. Рецептор-распознающий аппарат вирусов представлен гетеродимерами гликопротеинов Gn и Gc, которые организованным способом распределены по мембране вириона.
Структура генома флебовирусов
Геном флебовирусов как и других представителей семейства Bunyaviridae, включает три молекулы РНК: PHК L, РНК M, РНК S, имеющие на 5′- и 3′- концах уникальные для каждого геномного сегмента комплементарные последовательности. РНК L (–)полярности кодирует белок репликазы L. (–)РНК M кодирует предшественник гликопротеинов G1 и G2. (±)РНК S кодирует белок нуклеокапсида N на (–)полярном участке (ближе к 3′) и неструктурный белок NSs на (+)полярном участке (ближе к 5′) (рис. 2).

Рисунок 2. Схема структуры генома флебовирусов. Отмечены участки РНК, обладающие (–)- и (+)полярностью. Пунктирной линией обозначен сайт протеолиза белкового продукта.
NSs выполняет ряд функций, среди которых подавление индукции интерферона, усиление репликации и транскрипции вирусной РНК и определение круга хозяев [18]. NSs через цепочку белковых факторов способен приводить к инактивации противовирусной протеинкиназы R организма-хозяина [19].
Механизмы транскрипции и репликации РНК флебовирусов
Остановка транскрипции (–)участка РНК S определяется межгенным сигналом терминации. Похожие сигналы терминации находятся в 5′-концевой области РНК M и РНК L. В результате синтезируются кэпированные, но неполиаденилированные (и, следовательно, не такие стабильные, как клеточные мРНК) субгеномные РНК [18]. Также было показано [17], что в инфицированных клетках наблюдается ранняя экспрессия белка NSs, к тому же при детальном анализе состава вирионов обнаружили, что в вирусную частицу может упаковываться как три геномных цепи, так и еще три антигеномных цепи. Считается, что антигеномная РНК S присутствует в вирионе для осуществления ранней транскрипции мРНК, кодирующей NSs, поскольку этот неструктурный белок способен регулировать клеточные процессы, и чем раньше он начнёт работать в зараженной клетке, тем интенсивнее будет протекать вирусная инфекция.
Жизненный цикл флебовирусов
Жизненный цикл состоит из следующих стадий (рис. 3):

Рисунок 3. Схема, демонстрирующая основные этапы цикла флебовируса
Тосповирусы и тенуивирусы
Название рода Tospovirus происходит от сокращения названия вируса пятнистого увядания томатов (tomato spotted wilt virus, ТоSWV), впервые выделенного в 1930 году из зараженных растений томата. Этот вирус имеет очень широкий спектр хозяев и важное хозяйственное значение, борьба с ним ведется, в основном, за счет контроля численности трипсов.
Структура генома тосповирусов и тенуивирусов
Представители родов Тospovirus и Tenuivirus (тенуивирусы близки к тосповирусам, но не имеют липидной оболочки) являются единственными известными РНКвирусами растений с двусмысленным геномом [23]. Геном тосповирусов представлен тремя РНК-сегментами: большим, средним и малым (L, M, S). РНК L кодирует репликазу L. РНК S, подобно таковой у флебовирусов, кодирует белок нуклеокапсида N в (–)области и неструктурный белок NSs в (+)области. Эти области не пересекаются, они разделены межгенным некодирующим участком, содержащим сигналы терминации транскрипции. М-сегмент генома имеет принципиально отличную от РНК М флебовирусов структуру, являясь амбисенсной РНК. РНК М тосповирусов имеет область (–)полярности, в которой находится последовательность, кодирующая мРНК GnGc — предшественника поверхностных гликопротеинов, а также участок (+)полярности в 5′-области, кодирующий белок межклеточного транспорта NSm. Эти последовательности также разделены межгенным участком (рис. 4). Механизмы транскрипции и репликации РНК этих вирусов сходны с таковыми у флебовирусов [18].

Рисунок 4. Схема структуры генома тосповирусов. Отмечены участки РНК, обладающие (–)- и (+)полярностью. Пунктирной линией обозначен сайт протеолиза белкового продукта.
Отдельного внимания заслуживает неструктурный белок, закодированный в S-сегменте генома тосповирусов — NSs. Основной его функцией является супрессия противовирусного сайленсинга РНК, системы малых интерферирующих РНК [5], [24], распознающих вирусные РНК, что приводит к деградации последних [25]. Логично предположить, что синтез такого белка должен происходить как можно раньше, поэтому, возможно, по аналогии с белком NSs флебовирусов, ранняя транскрипция такой последовательности происходит в результате наличия в вирионе, помимо геномной цепи РНК S, еще и соответствующей ей антигеномной.
Вирусы с двусмысленным РНК-геномом из семейства Arenaviridae
Помимо семейства Bunyaviridae, амбисенсные РНК имеют представители семейства Arenaviridae. Аренавирусы являются таксономической группой вирусов позвоночных с сегментированным двусмысленным РНК-геномом. Вирусы, инфицирующие млекопитающих, определены в род Mammarenavirus, а заражающие рептилий — в роды Reptarenavirus и Hartmanivirus [26].
Вирионы аренавирусов, как и рассмотренных выше буньявирусов, плеоморфны, а их диаметр может варьировать от 40 до 200 нанометров в зависимости от вида, однако и частицы одного вида могут заметно различаться по размерам [27]. Границы вириона представлены липопротеидной оболочкой — производной клеточной мембраны, модифицированной равномерно распределёнными гликопротеиновыми комплексами (гетеродимерный гликопротеин GP1/GP2). Гликопротеины синтезируются в виде предшественника, который разрезается примерно пополам клеточной протеиназой на рецептор-распознающую субъединицу GP1 и трансмембранную субъединицу GP2. Последняя ответственна за слияние мембран при проникновении в цитоплазму [28]. В вирионе гликопротеины ассоциированны с лежащими на внутренней стороне мембраны молекулами матриксного белка Z, выстилающего внутреннюю поверхность мембраны, и белка нуклеокапсида N. Белок N способен связываться с РНК, распознавать кэп и ингибировать интерфероновый ответ. Структурный белок Z в клетке выполняет ряд функций (в том числе ингибирование трансляции клеточных мРНК и подавление апоптоза), являясь фактором созревания вирусных частиц (отвечает за инициацию сборки вирионов и за их отпочковывание).
Во время сборки вирусных частиц при формировании внешней оболочки иногда происходит захват субъединиц клеточных рибосом, по всей видимости, не играющих роли в вирусной инфекции (рис. 5).

Рисунок 5. Схематичное изображение структуры вириона аренавирусов
Значительная часть представителей семейства вызывает хронические и, как правило, бессимптомные инфекции у грызунов. При контакте человека с такими вирусами может развиваться острая и тяжелая инфекция, часто — геморрагическая лихорадка (например, в случае инфекции вирусом лихорадки Ласса, LasV). Вирус лимфоцитарного хориоменингита (LCMV), первый выделенный аренавирус, является нейроинвазивным. Попав в организм человека (например, через выделения грызунов), вирионы с током крови преодолевают гематоэнцефалический барьер центральной нервной системы и вызывают воспаления мозговых оболочек [29].
Структура генома аренавирусов

Рисунок 6. Схема структуры генома аренавирусов. Отмечены участки РНК, обладающие (–)- и (+)полярностью. Пунктирными линиями обозначены сайты протеолиза белковых продуктов.
РНК L на (–)участке несет последовательность, комплементарную гену репликазы L, и рамку матриксного и регуляторного белка Z на (+)участке в 5′-концевой части геномной РНК.
РНК S кодирует белок нуклеокапсида N в области (–)полярности и содержит рамку считывания GP1GP2 — предшественника поверхностных гликопротеинов GP1/GP2 (также в 5′-концевой части геномного сегмента).
Транскрипция и репликация генома аренавирусов
Переключение на репликацию связано с наличием белка N: когда его накапливается такое количество, что он начинает покрывать строящиеся цепи РНК, это, вероятно, влияет на конформацию репликазы и приводит к проскоку сигналов терминации транскрипции в виде межгенных шпилек. РНК S при репликации накапливается в больших количествах, так как нужно много копий белка нуклеокапсида, а также гликопротеинов (для экспрессии последних необходим предварительный синтез антигеномной РНК).
Заключение
Такая необычная организация двусмысленных геномных сегментов является интересным способом представления двух кодирующих последовательностей в одном геномном сегменте. На примере вирусов с двусмысленными РНК-геномами заметно, насколько изобретательной может быть эволюция вирусных РНК. Поскольку вирусы с двусмысленными РНК-геномами до сих пор удерживают определенную нишу, можно утверждать, что такой способ кодирования обладает некоторыми преимуществами по сравнению с более привычным для родственных вирусов способом, использующим только (–)РНК-сегменты.
Как возникли амбисенсные РНК и почему поддержались отбором, до сих пор остается одной из загадок современной вирусологии.
Благодарности от автора
Я благодарю доктора биологических наук, профессора кафедры вирусологии биологического факультета МГУ им. М.В. Ломоносова Аграновского Алексея Анатольевича за интересные и содержательные лекции по молекулярным процессам РНК-вирусов и вдохновение на написание данной статьи. Также выражаю благодарность художнику Андрею Занкевичу, чьи наглядные и яркие иллюстрации украшают данную статью.
Обратная транскриптаза (также известная как ревертаза или РНК-зависимая ДНК-полимераза) — фермент (КФ 2.7.7.49), катализирующий синтез ДНК на матрице РНК в процессе, называемом обратной транскрипцией.
Называется так потому, что большинство процессов транскрипции в живых организмах происходит в другом направлении, а именно, с молекулы ДНК синтезируется РНК-транскрипт.
Содержание
Значение для вирусов
Обратная транскрипция необходима, в частности, для осуществления жизненного цикла ретровирусов, например, вирусов иммунодефицита человека и T-клеточной лимфомы человека типов 1 и 2. После попадания вирусной РНК в клетку обратная транскриптаза, содержащаяся в вирусных частицах, синтезирует комплементарную ей ДНК, а затем на этой цепи ДНК, как на матрице, достраивает вторую цепь.
Значение для эукариот
Ретротранспозоны эукариот кодируют обратную транскриптазу, которая используется ими для встраивания в геном хозяина подобно тому, как это происходит у вирусов.
Роль в генетической инженерии
В генетической инженерии обратную транскриптазу используют для получения кДНК — копии эукариотического гена, не содержащей интронов. Для этого из организма выделяют зрелую мРНК, кодирующую соответствующий генный продукт (белок, РНК) и проводят с ней в качестве матрицы обратную транскрипцию. Полученную кДНК можно трансформировать в клетки бактерий для получения трансгенного продукта.
См. также
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "РНК-зависимая ДНК-полимераза" в других словарях:
РНК-зависимая ДНК-полимераза — * РНКзалежная ДНК полімераза * RNA dependent DNA polymerase … Генетика. Энциклопедический словарь
Обратная транскриптаза РНК-зависимая ДНК-полимераза ревертаза — Обратная транскриптаза, РНК зависимая ДНК полимераза, ревертаза * зваротная транскрыптаза, РНК залежная ДНК полімераза, рэвертаза * reverse transcriptase or PTase or RNA dependent DNA polymerase ретровирусный многофункциональный фермент класса… … Генетика. Энциклопедический словарь
Транскриптаза обратная РНК-зависимая ДНК-полимераза ревертаза — Транскриптаза обратная, РНК зависимая ДНК полимераза, ревертаза * транскрыптаза зваротная, РНК залежная ДНКполімераза, рэвертаза * reverse transcriptase … Генетика. Энциклопедический словарь
ДНК-зависимая ДНК-полимераза — Трехмерная структура ДНК связывающих спирально шпилечных участков в человеческой бета ДНК полимеразе ДНК полимераза фермент, участвующий в репликации ДНК. Ферменты этого класса катализируют полимеризацию дезоксирибонуклеотидов вдоль цепочки… … Википедия
Полимераза — * полімераза * polymerase фермент, катализирующий синтез полимера (см.). Существуют ДНК зависимая РНКполимераза, катализирующая синтез РНК через транскрипцию ДНК; РНК зависимая ДНК полимераза, катализирующая синтез цепи ДНК на информационной РНК… … Генетика. Энциклопедический словарь
ДНК-праймаза — * ДНК праймаза * DNA primase фермент, осуществляющий синтез РНК праймера (), необходимого для синтеза фрагментов Оказаки, а также РНК праймеров в процессе синтеза репликативной формы ДНК бактериофагов. У E. coli ДНК п. кодируется геном dna C (М.… … Генетика. Энциклопедический словарь
ДНК — Двойная спираль ДНК Дезоксирибонуклеиновая кислота (ДНК) один из двух типов нуклеиновых кислот, обеспечивающих хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. Основная… … Википедия
РНК-полимераза РНК-синтетаза — РНК полимераза, РНК синтетаза * РНКполімераза, РНК сінтэтаза * RNA polymerase or RNA synthetase 1. Фермент, ответственный за транскрипцию перевод информации с молкул ДНК на молекулу РНК. Он копирует информацию, содержащуюся на фрагменте ДНК путем … Генетика. Энциклопедический словарь

Выделяют ДНК-зависимую ДНК-полимеразу (КФ 2.7.7.7), использующую в качестве матрицы одну из цепей ДНК, и РНК-зависимую ДНК-полимеразу (другое название обратная транскриптаза, КФ 2.7.7.49), способную также к считыванию информации с РНК (обратная транскрипция).
ДНК-полимеразу считают холоферментом, поскольку для нормального функционирования она требует присутствия ионов магния в качестве кофактора. В отсутствии ионов магния о ней можно говорить как об апоферментe.
ДНК-полимераза начинает репликацию ДНК, связываясь с отрезком цепи нуклеотидов. Среднее количество нуклеотидов, присоединяемое ферментов ДНК-полимеразой за один акт связывания/диссоциации с матрицей, называют процессивностью.
Содержание
Действие ДНК-полимеразы
Некоторые ДНК-полимеразы обладают также способностью исправлять ошибки во вновь собираемой цепочке ДНК. Если происходит обнаружение неправильной пары нуклеотидов, ДНК-полимераза откатывается на один шаг назад. Благодаря своему экзонуклеазному действию ДНК-полимераза может исключить неправильный нуклеотид из цепочки и затем вставить на его место правильный, после чего репликация продолжается в нормальном режиме.
Многообразие ДНК-полимераз
Структура ДНК-полимераз достаточно жестко фиксирована. Их каталитические субъединицы очень мало различаются в различных видах живых клеток. Такая фиксация структуры обычно появляется там, где отсутствие разнообразия обусловлено огромной важностью или даже незаменимостью для функционирования клетки.
Генами некоторых вирусов тоже кодируются особые ДНК-полимеразы, которые могут избирательно реплицировать вирусные ДНК. Ретровирусы обладают геном необычной ДНК-полимеразы, называемой еще обратной транскриптазой, являющейся РНК-зависимой ДНК-полимеразой и осуществляющей сборку ДНК на основе шаблонной РНК.
ДНК-полимераза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных разными цветами), лигирующая повреждённую цепь ДНК
Семейства ДНК-полимераз
На основании своей структуры ДНК-полимеразы могут быть разбиты на семь различных семейств: A, B, C, D, X, Y, и RT.
Семейство A
Семейство A включает в себя репликативные и восстановительные ДНК-полимеразы. Репликативные члены этого семейства представлены, например, хорошо исследованной ДНК-полимеразой вируса Т7 или эукариотической митохондриальной ДНК-полимеразой γ. Среди восстановительных полимераз мы находим такие примеры как ДНК-полимераза I E. coli, полимераза I из Thermus aquaticus или полимераза I Bacillus stearothermophilus. Восстановительные полимеразы участвуют в процессе устранения ошибок в собираемой ДНК, а также в обработке фрагментов Оказаки.
Семейство B
В семейство B в основном входят восстановительные полимеразы, в том числе основные эукариотические ДНК-полимеразы α, δ, и ε, а также ДНК-полимераза ζ. К этому семейству также относят ДНК-полимеразы некоторых бактерий и бактериофагов, например бактериофагов T4, Phi29 and RB69. Эти ферменты используются в синтезе и 3’-5’, и 5’-3’ моноцепей ДНК. Отличительной особенностью полимераз этого семейства является замечательная точность репликации. Многие также обладает сильным 3’-5’-экзонуклеазным действием (за исключением ДНК-полимераз α и ζ, у которых способности корректировать ошибки не наблюдается).
Семейство C
Полимеразы этого семейства — в основном бактериальные хромосомные репликативные ферменты, обладающие, кроме того, 3’-5’-экзонуклеазным действием.
Семейство D
Полимеразы этого семейства недостаточно изучены. Все известные образцы считаются репликативными полимеразами и обнаружены у архей субдомена Euryarchaeota.
Семейство X
Семейство Y
Полимеразы этого семейства отличаются от прочих низкой производительностью на целостных шаблонах, а также способностью осуществлять репликацию на шаблонах поврежденных ДНК. Вследствие этого члены семейства называются полимеразами транслезионного синтеза. В зависимости от характера повреждения (лезии) ТЛС-полимеразы могут восстановить исходную цепочку. Ошибка может и не быть восстановлена, что приводит к мутациям. Страдающие Xeroderma pigmentosum, например, обладают мутантным геном ДНК-полимеразы η (eta), который толерантен к повреждениям, однако другие полимеразы, например ζ (относящаяся к семейству B), страдают от мутациям, что, как считается, приводит к предрасположенности к онкологическим заболеваниям.
Другие члены этого семейства — человеческие полимеразы ι, κ, а также концевая дезоксинуклеотидил-трансфераза Rev1. У E.coli имеются две ТЛС-полимеразы: IV (DINB) и V (UMUC).
Семейство RT
Семейство обратных транскриптаз (название семейства происходит от англ. reverse transcriptase) содержит полимеразы, обнаруженные как у ретровирусов, так и у эукариот. Они являются РНК-зависимыми ДНК-полимеразами, то есть, в отличие от описанных выше ферментов, используют в качестве матрицы для синтеза РНК, а не ДНК. Эукариотические полимеразы в основном представлены теломеразами. Эти полимеразы используют шаблонную РНК для синтеза цепочки ДНК.
Прокариотические ДНК-полимеразы
У бактерий обнаружено пять ДНК-полимераз:
- ДНК-полимераза I задействована в восстановлении ДНК, обладает и 5'-3', и 3'-5'-экзонуклеазным действием;
- ДНК-полимераза II участвует в репликации поврежденной ДНК. Обладает способностью 5'-3'-удлинения цепочки и 3'-5'-экзонуклеазным действием;
- ДНК-полимераза III — основная полимераза бактерий, обладающая также 3'-5'-экзонуклеазным действием;
- ДНК-полимераза IV, ДНК-полимераза семейства Y;
- ДНК-полимераза V, ДНК-полимераза семейства Y, принимающая участие в пропуске поврежденных участков ДНК.
Эукариотические ДНК-полимеразы
Эукариоты содержат по меньшей мере пятнадцать видов ДНК-полимераз [1] :
- ДНК-полимераза α выступает сначала в роли праймазы, синтезируя праймер ДНК, а затем как нормальная полимераза, присоединяя к этому праймеру нкулеотиды. После того, как длина цепочки достигнет около 20 нуклеотидов [2] , к транскрипции приступают полимеразы δ и ε;
- ДНК-полимераза β задействована в восстановлении ДНК;
- Pol γ, осуществляющая репликацию митохондриальной ДНК;
- ДНК-полимераза δ — основная полимераза эукариот. Она высокопроизводительна, а также обладает 3'-5'-экзонуклеазным действием;
- ДНК-полимераза ε, иногда замещающая ДНК-полимеразу δ во время синтеза 3’-5’-моноспирали. Основное назначение этой полимеразы неясно;
- ДНК-полимеразы η, ι, κ, и Rev1 из семейства Y, а также ζ из семейства B. Эти полимеразы задействованы в пропуске поврежденных участков ДНК [3] .
- Существуют также другие эукариотические ДНК-полимеразы, которые пока недостаточно изучены: θ, λ, φ, σ и μ.
Обнаружены и другие эукариотические полимеразы.
Ни одна эукариотическая полимераза не может отщеплять праймеры, то есть не обладает 5’-3’-экзонуклеазным действием. Эту функцию выполняют другие ферменты. Только полимеразы, осуществляющие элонгацию (γ, δ и ε) обладают 3'-5'-экзонуклеазными свойствами.
См. также
Ссылки
Внешние ссылки
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "ДНК-зависимая ДНК-полимераза" в других словарях:
ДНК-зависимая РНК-полимераза — * ДНКзалежная РНК полімераза * DNA dependent RNA polymerase фермент, с участием которого происходит ДНК зависимый синтез РНК (см.). У прокариот существует два типа ДНК з. РНК п.: ДНК праймаза (см.) катализирует синтез РНК праймера () для… … Генетика. Энциклопедический словарь
ДНК-зависимая РНК-полимераза — Фермент, осуществляющий ДНК зависимый синтез РН ; у прокариот существует 2 типа ДНК з.РНК п.: ДНК праймаза катализирует синтез РНК затравки для фрагментов Оказаки при репликации ДНК, в то время как РНК полимераза синтезирует все остальные… … Справочник технического переводчика
ДНК-зависимая РНК-полимераза — DNA dependent RNA polymerase ДНК зависимая РНК полимераза. Фермент, осуществляющий ДНК зависимый синтез РНК ; у прокариот существует 2 типа ДНК з.РНК п.: ДНК праймаза катализирует синтез РНК… … Молекулярная биология и генетика. Толковый словарь.
ДНК-зависимая ДНК-полимераза ДНКполимераза — ДНК зависимая ДНК полимераза, ДНКполимераза * ДНК залежная ДНК полімераза, ДНК полімераза * DNA dependent DNA polymerase or DNA polymerase фермент, катализирующий полимеризацию (см.) дезоксирибонуклеозидных трифосфатов в полимерную… … Генетика. Энциклопедический словарь
РНК-зависимая ДНК-полимераза — Обратная транскриптаза (также известная как ревертаза или РНК зависимая ДНК полимераза) фермент (КФ 2.7.7.49), катализирующий синтез ДНК на матрице РНК в процессе, называемом обратной транскрипцией. Называется так потому, что большинство… … Википедия
РНК-зависимая ДНК-полимераза — * РНКзалежная ДНК полімераза * RNA dependent DNA polymerase … Генетика. Энциклопедический словарь
Обратная транскриптаза РНК-зависимая ДНК-полимераза ревертаза — Обратная транскриптаза, РНК зависимая ДНК полимераза, ревертаза * зваротная транскрыптаза, РНК залежная ДНК полімераза, рэвертаза * reverse transcriptase or PTase or RNA dependent DNA polymerase ретровирусный многофункциональный фермент класса… … Генетика. Энциклопедический словарь
Читайте также: