
Семейства роды и виды вирусов
Обновлено: 25.04.2024
ПОКСВИРУСЫ (Poxviridae; англ. pox пустула + вирусы) — семейство крупных ДНК-содержащих вирусов.
Ранее П. называли вирусами оспы. П. самые крупные вирусы. Они способны инфицировать млекопитающих, включая человека (см. Оспа животных, у человека), птиц, рыб, амфибий и даже насекомых. Это семейство включает девять родов (табл.).
Вирион П. кирпичеобразной или овоидной формы, имеет размеры 300—450 нм X 150—260 нм. Сердцевина вириона представляет собой ДНК, ассоциированную с белками. Эта структура заключена в белковую оболочку, построенную из хорошо различимых субъединиц, и имеет сложную симметрию. Наружная оболочка вириона содержит липиды и покрыта трубчатыми или нитеобразными белковыми структурами, организованными в характерные витые спирали. Под наружной оболочкой располагаются латеральные тела, функция к-рых не выяснена. В вирионах П. выявлено ок. 100 вирусоспецифических белков, часть из к-рых является гликопротеинами. По крайней мере 10 из них обладают ферментативной активностью. Среди них ДНК-зависимая РНК-полимераза, поли(А)-полимераза, мРНК-гуанилтрансфераза, мРНК-(гуанин-7) метилтрансфераза, мРНК-(рибоза-2') метилтрансфераза, НК-зависимые нуклеозид-трифосфат фос-фогидролазы, дезоксирибонуклеаза (одна из трех, обнаруживаемых в вирионе), протеинкиназа, фермент, восстанавливающий разрывы ДНК, и др. Наличие ферментов позволяет П. осуществлять транскрипцию ДНК на ранних этапах без участия ферментов клетки-хозяина.
Геном П. представляет собой единую двухцепочечную молекулу ДНК с мол. весом (массой) 130—240 X 106 дальтон; содержание гуанина + цитозина у поксвирусов позвоночных составляет 35—40%, а у поксвирусов насекомых — ок. 26%. ДНК поксвирусов не обладает инфекционностью. При обработке ДНК, изолированной из вирионов П., бактериальными рестриктазами ДНК расщепляется на фрагменты, количество и размеры к-рых различны у отдельных П. Этот метод анализа используют при изучении происхождения нек-рых П. и их принадлежности к определенному роду.
У П. позвоночных имеется по крайней мере один антиген, являющийся внутренним компонентом вириона, который дает перекрестную серол. реакцию с представителями всех П. Внутри каждого рода П. позвоночных наблюдаются четкие перекрестные серол, реакции.
Для П. характерна высокая частота рекомбинации (см.), а также типичный для вирусов только этого семейства феномен, который получил название негенетической реактивации (см. Негенетические взаимодействия вирусов); этот феномен наблюдается при взаимодействии П. позвоночных как в пределах рода, так и между представителями различных родов. У П. получены и описаны мутанты, различающиеся по ряду свойств: размеру и характеру поражений хорион-аллантоисной оболочки куриного эмбриона (напр., так наз. белые мутанты вирусов оспы кроликов, коров, обезьян, образующие белые оспины на хорион-аллантоисной оболочке в отличие от оспин красного цвета, вызываемых родительскими штаммами); по размеру и характеру бляшек, образуемых на культуре ткани; способности к репродукции при повышенной температуре; устойчивости к физ.-хим. воздействиям и т. п. Получены условно-летальные мутанты П. (температурно-чувствительные и зависимые от клетки-хозяина), которые с успехом используются при изучении механизмов репродукции этих вирусов.
При заражении животных П. чаще всего возникают кожные поражения в виде сыпи папулезно-пустулезного характера. Вирус оспы обезьян вызывает у них при естественном заболевании клин, проявления двух типов: отек лицевой части головы и шеи, который может привести к гибели от асфиксии, и кожные негене-рализованные поражения. При оспе мышей, вызываемой вирусом эктромелии, поражаются либо кожа, либо внутренние органы. Наиболее характерны при этом заболевании поражения кожи конечностей, заканчивающиеся некрозом и отторжением части конечности. Вирус оспы кроликов обладает высокой патогенностью для этих животных, и заболевание часто заканчивается летальным исходом. Парапоксвирусы копытных могут поражать и человека, вызывая, напр., заболевание, сопровождающееся появлением узелков на коже рук (узелки доильщиц). Вирусы оспы птиц вызывают заболевания, при к-рых кожные поражения носят пролиферативный характер, отчасти сходные с течением опухолей. Типичные опухоли развиваются при поражении лепорипоксвирусами — вирусами миксомы и фибромы кроликов, а также вирусом обезьян Яба, который может обусловливать образование опухолей и у человека. Вирус контагиозного моллюска также способен вызывать у человека мелкие опухоли кожи. Наибольшее значение для патологии человека имеет вирус натуральной оспы (см. Оспа натуральная). Это заболевание полностью ликвидировано во всем мире благодаря высокоактивной живой противооспенной вакцине (см. Оспопрививание), а также ряду организационных мероприятий по выявлению очагов заболевания, вакцинации и пр., которые проводились под эгидой ВОЗ с активным участием специалистов из Советского Союза и при использовании миллионов доз оспенной вакцины, изготовленной в нашей стране.
Строение вирусов. Классификация вирусов
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
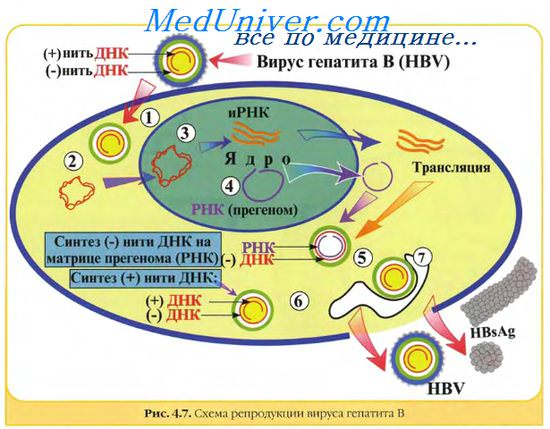
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
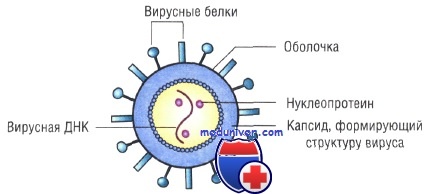
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Классификация вирусов. Современная номенклатура в вирусологии.
Установлено, что все изученные организмы поражаются вирусами. Множество различных вирусов вызывают заболевания или латентно инфицируют позвоночных и беспозвоночных животных, а также простейших, растения, грибы и бактерии. Известно более 4000 разных вирусов, из которых несколько сотен инфицируют людей и животных.
Международным комитетом по таксономии вирусов (ICTV, 2000) разработана единая система классификации и номенклатуры вирусов, которая основана на изучении различных свойств вирусов, таких как размер, морфология и структура вириона, его устойчивость к воздействию различных факторов и антигенные свойства.
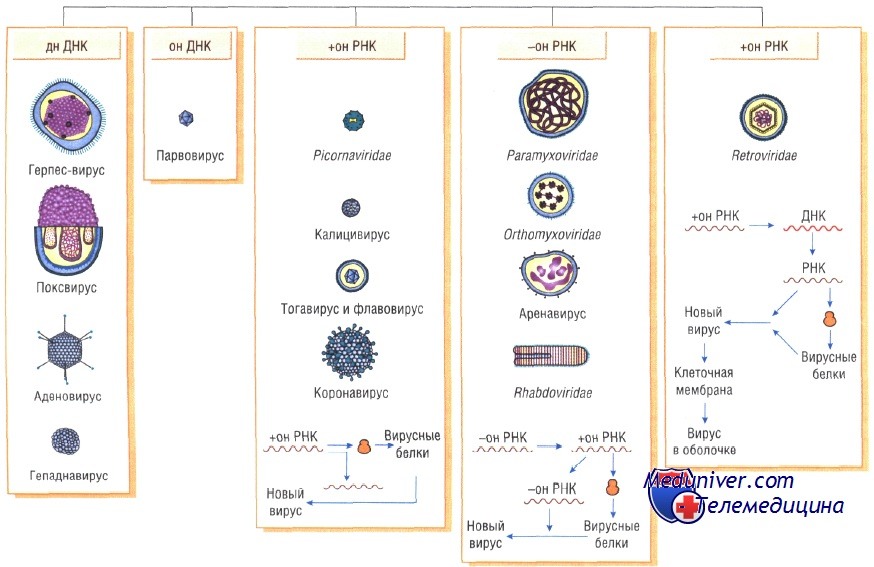
Классификация вирусов
На сегодня главным критерием в таксономии вирусов являются: 1) тип и структура вирусного генома 2) стратегия репликации вируса 3) структура вириона. Полное или частичное секвенирование вирусного генома увеличивает таксономическую информацию и очень часто используется с целью идентификации вируса.
Сведения о нуклеотидной последовательности для всех таксономических групп вирусов имеются в опубликованной базе данных (Генбанк, Национального Центра биотехнологической информации, Национальной медицинской лаборатории, Национального Института здоровья, Бетезда, Мериленд, США). Это достижение в большинстве случаев укорачивает таксономическую идентификацию, хотя традиционные методы все еще часто используются по экономическим соображениям. Универсальная система таксономии вирусов включает несколько уровней: порядок, семейство, подсемейство, род и вид. Патогенные вирусы позвоночных (человека и животных) в соответствии с современной системой классификации вирусов объединены в 2 (Mononegavirales и Nidovirales) порядка и 28 семейств, из которых 10 являются ДНК-вирусами и 18 РНК-вирусами.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Систематика вирусов. Особенности классификации вирусов. Основные критерии таксономической классификации вирусов.
Вирусы отнесены к царству Vira. В основу их классификации положен тип нуклеиново кислоты, образующей геном. Соответственно выделяют рибовирусы (РНК-вирусы) и дезоксирибовирусы (ДНК-вирусы).
Для вирусов предложены следующие таксономические категории (по восходящей): Вид (Species) —> Род (Genus) —> Подсемейство (Subfamilia) —> Семейство (Familia). Но категории подсемейств и родов разработаны не для всех вирусов. Видовые названия вирусов обычно связывают с вызываемыми ими заболеваниями (например, вирус бешенства) либо по названию места, где они были впервые выделены (например, вирусы Коксаки, вирус Эбола). Если семейство включает большое количество видов, то видовые названия дают в соответствии с антигенной структурой и разделяют их на типы (например, аденовирус 32 типа или вирус герпеса 1 типа). Реже используют фамилии учёных, впервые их выделивших (например, вирус Эпстайна-Барр или вирус саркомы Рауса). Иногда используют устаревшие названия групп вирусов, отражающих их уникальные эпидемиологические характеристики (например, арбовирусы). Классификационные признаки патогенных для человека вирусов приведены в табл. 3-1.
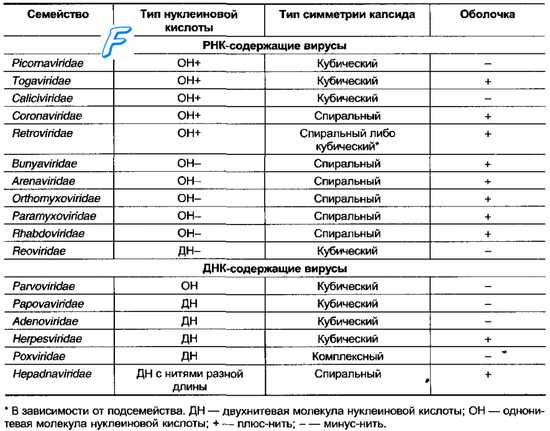
Таблица 3-1. Семейства вирусов, включающие возбудителей инфекций человека
К вирусам отнесены вироиды [от virus и греч. eidos, сходство] — мелкие кольцевые однонитевые суперспирализованные молекулы РНК (аналогичную организацию имеет геном вируса гепатита D). Поскольку у вироидов нет белковой оболочки, они не проявляют выраженных иммуногенных свойств, и поэтому их нельзя идентифицировать серологическими методами. Вироиды вызывают заболевания у растений. В качестве безымянного таксона в царство Vira также включены и прионы.
Основные критерии таксономической классификации вирусов
При систематизировании вирусов выделяют следующие основные критерии: сходство нуклеиновых кислот, размеры, наличие или отсутствие суперкапсида, тип симметрии нуклеокапсида, характеристика нуклеиновых кислот (молекулярная масса, тип кислоты (ДНК или РНК), полярность [плюс или минус], количество нитей в молекуле либо наличие сегментов, наличие ферментов), чувствительность к химическим агентам (особенно к эфиру), антигенная структура и иммуногенность, тропизм к тканям и клеткам, способность образовывать тельца включений.
Дополнительный критерий — симптоматология поражений: способность вызывать генерализованные поражения либо инфекции с первичным поражением определённых органов (нейроинфекции, респираторные инфекции и др.).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
РАБДОВИРУСЫ (Rhabdoviridae; греч. rhabdos палка, стержень + вирусы) — семейство РНК-содержащих вирусов, включающее роды вирусов Lyssavirus (вирусы бешенства) и Vesiculovirus (вирусы везикулярного стоматита), а также несгруппированные вирусы позвоночных, насекомых и растений. К Р. условно отнесены близкие к ним по свойствам вирусы Марбург и Эбола.

Рис. 1. Электронограммы вирионов вирусов везикулярного стоматита (а) и бешенства (б); х 100 000; по Мейдли (C.R. Madeley), 1972.

Рис. 2. Электронограммы атипичных форм вирионов вирусов: а — бешенства (укороченная форма); б — везикулярного стоматита (бациллярная форма); х 200 000; по Хаммелеру и Томассини, 1973.
Средние размеры Р.: длина 130— 300 нм, диам. 60—80 нм. Вирионы Р. имеют пулевидную форму, т. е. представляют цилиндр, один конец к-рого полукруглый, а другой — плоский (рис. 1). Вирионы Р. имеют, кроме пулевидной, бацилловидную форму. Пулевидная форма присуща преимущественно Р., инфицирующим позвоночных, бацилловидная — вирусам, инфицирующим растения. У ряда Р. позвоночных выявлены нетипичные формы вирионов (рис. 2). Бацилловидные формы вирионов с закругленными с обеих сторон концами описаны у вирусов везикулярного стоматита и у Р. рыб. Длинные филаментозные формы выявлены у Р. рыб; они также типичны для вирионов Марбург. Короткие частицы (1/3 длины типичного вириона) соответствуют дефектным интерферирующим частицам, выявленным у вирусов бешенства и везикулярного стоматита.
Сердцевина вириона (core) диам. 45—50 нм также пулевидной формы и покрыта липидсодержащей двухслойной оболочкой с отростками длиной 5—10 нм и диам. 3 нм. Нуклеокапсиды, выделенные из спонтанно или химически деградированного вируса везикулярного стоматита, представляют собой волнистые ленты шириной 8 — 10 нм и толщиной 3—4 нм. Средняя длина нуклеокапсида нормальных вирионов составляет ок. 3,5 мкм. Нуклеокапсид вируса бешенства также представляет собой одноцепочечную спираль диаметром 16 нм, периодичностью 7,5 нм и длиной, превышающей 1 мкм. В негативно окрашенных тонких срезах вирионов Р. видны пересекающиеся борозды глубиной 4,5—5 нм и центральный осевой канал. На поверхности частиц некоторых Р. обнаруживается ячеистость.
Мол. вес (масса) вирионов Р. 300—1000×10 6 , коэффициент седиментации 550—1000S, плавучая плотность в CsCl 1,19—1,20 г/см 3 , в сахарозе 1,17—1,19 г/см 3 . У вирусов бешенства и везикулярного стоматита мол. вес соответственно 475×10 6 и 385×10 6 , константа седиментации этих вирусов 600—625S. Вирионы Р. содержат 3—4% РНК, 64—67% белка, 20—26% липидов и 3—10% углеводов. РНК одноцепочечная, нефрагментированная с мол. весом 3,5—4 × 10 6 . Константа седиментации вирионной РНК 40— 45S, плавучая плотность 1,59—1,66 г/см 3 . Дефектные интерферирующие частицы (Т-частицы) содержат ок. 1/3 генома. В Т-частицах обнаружены двухцепочечные РНК, формирующие ковалентно связанную шпилечную структуру. В состав вириона входит РНК-зависимая РНК-полимераза, к-рая транскрибирует вирионную РНК в комплементарную мРНК и катализирует синтез полиаденилатной РНК как в клетках, так и в бесклеточных системах. При помощи электрофореза в полиакриламидном геле (см. Электрофорез) у Р. выявлено 5 структур белков, для которых предложены следующие наименования: L — крупный нуклеокапсидный белок с мол. весом 190 000; GP — мембранный гликопротеин с мол. весом 69 000; N — второй нуклеокапсидный белок с мол. весом 50 000; NS — третий нуклеокапсидный белок с мол. весом 40 000—50 000, ранее считавшийся неструктурным протеином; М — негликозилированный мембранный белок с мол. весом 29 000. У всех изученных Р. найден по меньшей мере один (белок N) и обычно два или три фосфопротеина.
Углеводы, выявленные у Р., включают глюкозу, галактозу, маннозу, фукозу, глюкозамин, галактозамин и нейраминовую к-ту. В вирусе везикулярного стоматита выявлены холестерин и фосфолипиды. Р. чувствительны к жирорастворителям, умеренно чувствительны к теплу и устойчивы при низких температурах. Вирусы бешенства, везикулярного стоматита (типы Нью-Джерси, Индиана, Кокал) и Керн-Каньон агглютинируют гусиные эритроциты. Гемагглютинация (см.) связана с цельным вирионом или с его укороченными формами. Реакция лучше протекает на холоду при pH 6,0 и весьма чувствительна к нормальным сывороточным ингибиторам. Рецепторы клеток, с к-рыми взаимодействуют Р., не разрушаются ими и перийодатом. Нейраминидазная активность у Р. не выявлена.

Рис. 3. Электронограмма клетки при почковании вирионов вируса везикулярного стоматита с ее поверхности: 1 — продольный срез вириона; 2 — поперечный срез вирионов; х 20 000; по Хаммелеру и Томассини (Hummaler, Tomassini), 1973.
Вирионы Р. синтезируются в цитоплазме. Вирусная РНК транскрибируется вирионной транскриптазой в несколько видов позитивных цепей РНК, которые действуют как матричная РНК (мРНК) в полирибосомных комплексах. Место образования зрелых частиц Р. варьирует в зависимости от вида вируса и клеток хозяина. Так, нуклеокапсид вируса везикулярного стоматита синтезируется в цитоплазме и затем почкуется из цитоплазматических мембран клеток (рис. 3). Сборка вирионов вируса бешенства осуществляется в ассоциации с гранулярными цитоплазматическими матрицами, которые соответствуют тельцам Бабеша—Негри инфицированных клеток. Оболочка вириона вируса бешенства может образовываться внутри или на краю цитоплазматической матрицы, однако описано также почкование вирионов на поверхности клетки.
Распространение Р. может осуществляться как биол, хозяевами, так и механическим путем с помощью членистоногих. Механическое распространение может осуществляться путем прямого контакта, аэрозольно, а также за счет укуса.
Лиссавирусы (Lyssavirus) — самостоятельный род в семействе Р., включающий 6 видов вирусов. Название рода происходит от греч, слова lyssa — бешенство. Типичным представителем лиссавирусов является вирус бешенства (см. Бешенство). К ним же относятся вирусы летучих мышей Лагоса, а также вирусы Мокола, Котонкан, Дувенхадж, Ободьянг.
Вирус летучих мышей Лагоса выделен из мозга летучей мыши, вирус Мокола — от землеройки и от ребенка с тяжелым поражением ц. н. с. (в Нигерии). При экспериментальном заражении вирусом Лагоса и Мокола у землероек развивается инф. процесс с летальным исходом. Вирусы Мокола и Лагоса у экспериментально инфицированных животных подобно вирусу бешенства распространяются по нервам в различные участки ц. н. с. Оба вируса вызывают у обезьян и собак инфекционное заболевание с летальным исходом.
По антигенной структуре близкими к вирусу Мокола являются вирусы Ободьянг и Котонкан, но по морфологии они сходны с вирусом эфемерной (кратковременной) лихорадки крупного рогатого скота, имеющим вирионы преимущественно конусовидной формы. Возможно, что вирус Котонкан способен инфицировать людей и домашних животных. Вирусы Котонкан и Ободьянг выделены от комаров соответственно в Нигерии и Эфиопии. Предполагается также, что вирус Котонкан является возбудителем заболевания коров, сходного с эфемерной лихорадкой.
Вирус Дувенхадж выделен от больного с клиникой бешенства. Хотя в мозгу больного и экспериментально зараженных мышей были выявлены тельца Бабеша — Негри, однако антисыворотка к вирусу бешенства в реакции иммунофлюоресценции дала отрицательный результат.
Везикуловирусы (Vesiculovirus) выделены от позвоночных и беспозвоночных животных. Название рода происходит от лат. слова vesicula — пузырек, поскольку при заболевании у животных наблюдается везикулярная сыпь. Типовым представителем рода является вирус везикулярного стоматита (см. Стоматит). Имеется по меньшей мере 2 антигенных типа — (Нью-Джерси и Индиана), которые могут быть дифференцированы в РСК и в реакции нейтрализации. Тип Индиана делится на 3 подтипа: Кокал и аргентинские штаммы, бразильские штаммы и Индиана С-штаммы. Вирусы Пири и Чандипура являются антигенно сходными друг с другом и с вирусами Индиана и Кокал.
Вирус везикулярного стоматита умеренно патогенен для лошадей и крупного рогатого скота — вызывает сравнительно невысокую заболеваемость животных. Заболевание у животных напоминает ящур, но протекает значительно легче; у людей оно протекает по типу гриппа и наблюдается редко, преимущественно у ветеринаров и работников вирусе л. лабораторий.
Р. растений включают большую группу вирусов, имеющих преимущественно бацилловидную форму и большую длину (до 380 нм), чем у других представителей Р. Их белковый состав сходен с белками вируса везикулярного стоматита, включая транскриптазу. Некоторые Р. растений имеют два белка М (М1 и М2). Р. быстро инактивируются при комнатной температуре, размножаются в растениях и членистоногих.
Библиография: Атабеков И. Г. Реализация генетической информации вирусных РНК, М., 1972; Бочаров А. Ф. и Бочаров Е. Ф. Персистенция вирусов, Новосибирск, 1979, библиогр.; Ген-дон Ю. 3. Молекулярная генетика вирусов человека и животных, М., 1975, библиогр.; Knudson D. L. Rhabdo-viruses, J. gen. Virol., v. 20,suppl.,p. 105, 1973.
Читайте также: