
Семейство вирусов ана я
Обновлено: 17.04.2024
РЕТРОВИРУСЫ (Retroviridae) - семейство вирусов, объединяющим признаком для к-рых служит наличие в структуре вирионов ревертазы (обратной транскриптазы) — фермента, синтезирующего ДНК на матрице РНК вируса.
Впервые Р. были описаны в 1908 г. Эллерманном и Бангом (V. Ellermann, О. Bang), изучавшими возбудителей эритро- и миелобластозов птиц. Позже Ф. Раус (1911) описал вирус саркомы кур, получивший название вирус саркомы Рауса. По классификации 1970 г. эти и другие сходные вирусы были объединены в род лейковирусов (Leukovirus; греч. leukos белый, т. е. вирусы, обусловливающие злокачественное белокровие, лейкозы). В 1975 г. эти вирусы были включены в классификацию вирусов как онкорнавирусы [греч. oncos опухоль + RNA (англ. ribonucleic acid) — опухолевые РНК-содержащие вирусы]. Однако эти названия долго не просуществовали, поскольку, с одной стороны, в группу этих вирусов входили вирусы, не только обусловливающие лейкозы, но и приводящие к развитию сарком и опухолей молочной железы, а с другой — в эту группу были включены вирусы, не обладащие онкогенными свойствами, но содержащие ревертазу. Наличие ревертазы, обеспечивающей характерный для таких вирусов обратный (лат. retro назад) поток информации (от РНК к ДНК), и послужило основанием для образования названия этого семейства — Retrоviridae, включающего подсемейства онковирусов, спумавирусов и лентивирусов (табл.).
Наиболее изученными являются вирусы подсемейства онковирусов (Oncovirinae). Им присуща круглая форма, диаметр не превышает 100 нм, а плавучая плотность составляет 1,15—1,19 г/см 3 . Онковирусы содержат РНК, к-рая представлена двумя агрегированными идентичными 35 S-молекулами, состоящими примерно из 10 000 нуклеотидов, мол. вес (масса) каждой молекулы составляет ок. 3*10 6 дальтон, они соединены между собой водородными связями в области 5'-концов. Кроме этой высокомолекулярной РНК, обычно называемой 70 S-РНК, в составе вирионов выявляются также низкомолекулярные РНК (4—5 S), представляющие собой клеточные sPHK или гРНК. Все РНК структурно входят в состав нуклеоида (плавучая плотность 1,26—1,31 г/см 3 ), покрытого оболочкой. Это образование, получившее название сердцевины вириона (плавучая плотность 1,21 — 1,23 г/см 3 ), окружено внешней оболочкой, имеющей отростки и состоящей из внешней и внутренней мембран. Морфологические структуры у онковирусов имеют определенные родоспецифические различия (см. рис. к ст. Онкогенные вирусы). Так, у онковирусов С сердцевина крупная и расположена центрально, у онковирусов В и D сердцевина расположена эксцентрично; нуклеоид у онковирусов В круглый, а у онковирусов D — цилиндрический. Отростки у онковирусов В и С длинные, а у онковирусов D и подрода онковирусов С млекопитающих — короткие.
Помимо онковирусов А и С обычных размеров, описаны также их минимальные формы, размеры к-рых ок. 30—40 нм. Минимальные формы содержат все белки, присущие вирионам нормальных размеров, но значительно меньшие по размерам молекулы рибонуклеиновой кислоты.
Хим. состав онковирусов следующий: РНК — ок. 1—2%, белки (5—8 различных полипептидов) — 60—70%, липиды — 30—40% и углеводы — ок. 1—2%. Главный компонент оболочки — гликопротеид (500—1000 молекул в одном вирионе) с мол. массой 50—80 килодальтон; второй гликопротеид имеет мол. массу 22—45 килодальтон. Присутствуют также негликозилированные белки с мол. массой 15 и 12 килодальтон. В составе сердцевины выделяют главный внутренний белок (мол. масса 24—36 килодальтон) и дополнительные белки, среди к-рых ревертаза (10—20 молекул в вирио-не) и другие ферменты (ДНК-лигаза, ДНК-полимераза, экзо- и эндонуклеазы, РНК-метилаза и др.). Вирусы, относящиеся к одному виду, имеют идентичные (или близкие) главные внутренние белки, названные группоспецифическим антигеном (gs-антиген); подвиды, входящие в состав одного вида, отличаются по антигенным свойствам белков оболочки или отдельных дополнительных внутренних белков. Внутри подвида также могут существовать определенные варианты.
РНК онковирусов — носитель генетических свойств вириона. В ней различают гены, кодирующие синтез наружных белков (ген env), ревертазы (ген pol), внутренних белков (ген gag) и гены, ответственные за трансформацию инфицированных клеток (ген src). Утрата гена src не влияет на способность вирусов репродуцироваться в клетках, тогда, как утрата любого из трех других генов делает вирус дефектным, нуждающимся в вирусе-помощнике.
В лабораторных условиях онковирусы культивируют на животных, для к-рых они обладают онкогенной потенцией, и в клетках как первичных, так и перевиваемых клеточных культур. Цикл репродукции онковирусов в клетках состоит из двух фаз. Первая фаза начинается с адсорбции и проникновения вируса в клетку — процессов, не отличающихся от аналогичных процессов у любых вирусов (см.). Она завершается характерной для онкогенных вирусов (см.) интеграцией с клеточным геномом ДНК-провируса, т. е. ДНК, к-рая синтезировалась ревертазой; на матрице онковирусной РНК, В результате в геном одной зараженной клетки может встроиться от одной до пяти ДНК-провируса, Вторая фаза заключается в экспрессии интегрированной ДНК-провируса, к-рая, однако, не начинается, пока зараженные клетки не пройдут стадию митоза. В результате экспрессии ДНК-провируса образуются РНК онковирусов, затем белки вирусов и, наконец, онковирусы. Завершающим этапом в образовании онковирусов является выход их иа клетки, в процессе к-рого синтезировавшаяся сердцевина вируса связывается с внутренней поверхностью измененного участка плазматической мембраны клетки и по мере выхода из последней окружается этой мембраной.
Онковирусы не вызывают гибели клеток: инфицирование ими носит хронический характер. Репродукция онковирусов сопровождается антигенной конверсией клеток, поскольку на плазматической мембране клеток появляются неклеточные полипептиды — продукты генов env и gag. В случае наличия в геноме онковируса гена src в клетках синтезируется так наз. раковый белок, к-рый обусловливает трансформацию клетки в опухолевую (раковую).
Клетки могут получать геном онковирусов наследственным путем (вертикальный путь передачи). В этом случае все потомство особи, содержащей в своем геноме ДНК-прови-руса, также будет содержать в своей ДНК информацию для синтеза онковирусов. При этом не обязательно, чтобы в случае наследования клетками онковирусных геномов, происходила неизбежная реализация заключенной в них информации. Чтобы такая реализация имела место, необходимо воздействие на эти клетки определенных хим. соединений, известных под названием канцерогенов. Возможна, однако, и спонтанная индукция синтеза онковирусов.
Для выявления онковирусов в тканях (клетках) существует ряд взаимоподтверждающих друг друга методов. Широко применяются методы электронно-микроскопических исследований для обнаружения в клетках онковирусов, обладающих специфической структурой. Используются также методы физикохимические и биохимические (обнаружение вирусных частиц с характерной плавучей плотностью, содержащих 70 S— РНК и ревертазу), иммунологические (выявление в клетках специфических для онковирусов антигенов) и биологические (установление в опытах на животных или с клеточными культурами специфического биологического эффекта).
Онковирусы широко распространены у животных, причем для многих из них доказана роль этиологического фактора в возникновении опухолей (напр., вирус рака молочных желез мышей, вирус саркомы птиц и т. д.). Многими исследователями разделяется гипотеза Хюбнера и Тодаро (R. J. Huebner, G. J . Todaro), сформулированная в 1969 г., о том, что у человека (как и у животных) злокачественные опухоли могут возникать в результате активации унаследованных геномов ретровирусов. Однако у человека такие онковирусы не выявлены. Обнаружение в перевиваемых опухолевых клетках человека онковирусов, в частности подобных тем, что вызывали рак молочной железы у обезьян, большинство вирусологов считают лишь лабораторными вирусами-кон-таминантами. Одним из широко распространенных аргументов против этиологической роли онковирусов в патологии человека принято считать тот общеизвестный факт, что злокачественные опухоли человека не заразны.

Рис. 1. Электронограмма ультратонкого среза спумавирусов: 1 — вирусы внутри клетки; 2 — вирусы вне клетки; х 100 000.
Спумавирусы (Spumavirinae; англ. spume пена), или пенящие вирусы, синцитиальные, симпластообразующие вирусы,— подсемейство, объединяющее группу вирусов, к-рые при инфицировании эпителиоидных и фибробластных клеток обусловливают слияние этих клеток с образованием синцитиев, содержащих до 100 и более ядер, и появление в цитоплазме многочисленных вакуолей, что напоминает пену. Под электронным микроскопом спумавирусы (рис. 1) сходны с вирионами онковирусов А. Спумавирусы выделены от обезьян, кошек, хомяков, крупного рогатого скота и человека; вызывают персистентные бессимптомные инфекции у естественных хозяев и экспериментальных животных. Распространяются спумавирусы как респираторным путем (горизонтальный путь передачи), так и наследственным путем. Способность к опухолеобра-зованию или трансформации клеток, равно как и патогенность у этих вирусов, не установлены. Для инфекционного процесса, обусловленного спумавпрусами, свойственно медленное развитие: от момента инфицирования до выявления характерных специфических изменений в цитоплазме клеток проходит несколько недель. Внеклеточный вирус не обнаруживается; его распространение происходит непосредственно из клетки в клетку.

Лентивирусы (Lentivirinae; лат. lentus медленный) — подсемейство Р., объединяющее группу вирусов, характерной чертой к-рых является медленное развитие обусловленного ими инфекционного процесса (см. Медленные вирусные инфекции). Лентивирусы в естественных условиях выделяются только от овец. К ним относятся вирусы висна (рис. 2), мэди и прогрессирующей пневмонии овец. Морфологически они напоминают онковирусы С, размножаются в культурах клеток овец, вызывая симпластообразование. Имеются указания на способность вируса висна вызывать трансформацию клеток, однако опухоли эти вирусы не индуцируют. В антигенном отношении лентивирусы близки, что дает основание нек-рым исследователям рассматривать их как варианты одного и того же вируса.
Вирус мэди вызывает после длительного (2—3 года) инкубационного периода медленно прогрессирующую хроническую интерстициальную пневмонию, заканчивающуюся гибелью животных спустя год после появления признаков заболевания. При вскрытии павших овец у них обнаруживается лимфоцитарная диффузная периваскуляр-ная и перибронхиальная инфильтрация, диффузная пролиферация мезенхимы, увеличение перибронхи-альных, трахеобронхиальных и ме-диастинальных лимфатических узлов, отмечаются изменения ив ц. н. с. В эксперименте вирусом мэди удается инфицировать кроликов, морских свинок и мышей; помимо клеток овец, вирус мэди можно культивировать в клетках почек эмбрионов коров и надпочечников хомячков.
Вирус висна вызывает демиели-низирующее заболевание определенных пород овец, разводимых в Исландии. После инкубационного периода продолжительностью до 4 лет отмечаются симптомы поражения ц. н. с.: скованность, хромота, боковое отклонение головы, затем наступают парезы и параличи. Болезнь длится около месяца, все заболевшие овцы погибают. Патологоанатомические изменения характеризуются признаками диффузного энцефалита с демиелинизацией. При внутрилегочном инфицировании овец вирусом висна отмечаются те же изменения, что и при болезни мэди.
Прогрессирующая пневмония овец впервые была описана в Исландии, но затем наблюдалась во Франции, Голландии, Южной Африке. Инкубационный период может продолжаться до 3 лет. Симптомы заболевания сходны с наблюдавшимися при болезни мэди, однако патологоанатомическая картина несколько отлична от таковой при мэди. Через 3—12 мес. после появления первых симптомов заболевания наступает гибель животных в результате нарушения дыхания.
Таблица. КЛАССИФИКАЦИЯ ВИРУСОВ СЕМЕЙСТВА РЕТРОВИРУСОВ (RETROVIRIDAE) ЖИВОТНЫХ
Строение вирусов. Классификация вирусов
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
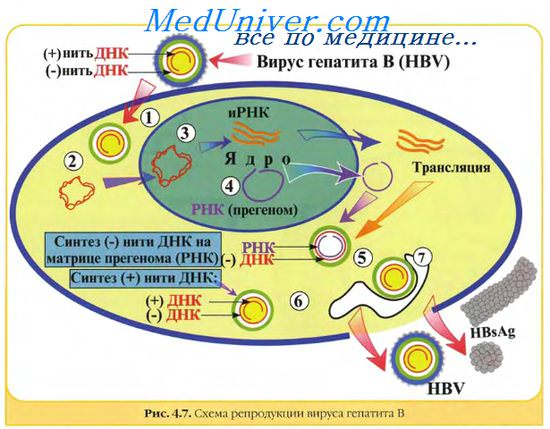
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
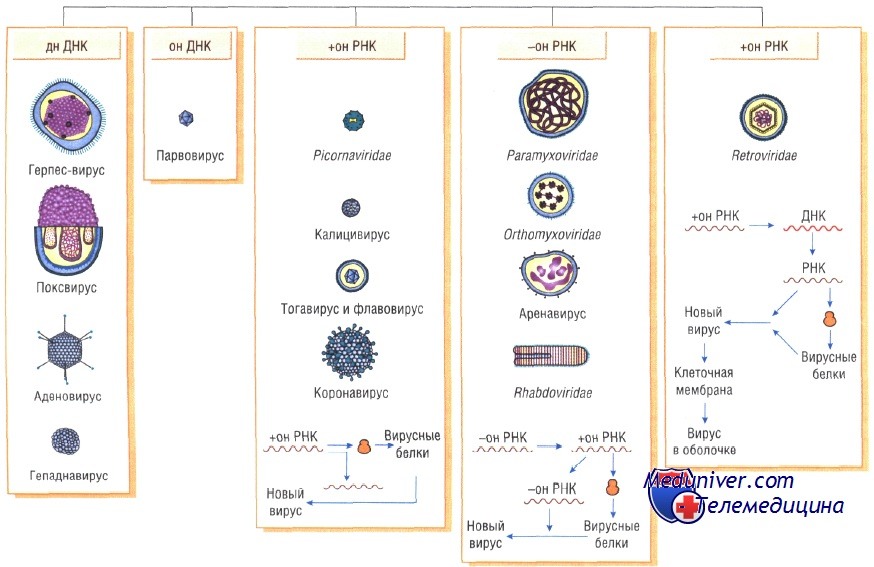
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Классификация вирусов. Современная номенклатура в вирусологии.
Установлено, что все изученные организмы поражаются вирусами. Множество различных вирусов вызывают заболевания или латентно инфицируют позвоночных и беспозвоночных животных, а также простейших, растения, грибы и бактерии. Известно более 4000 разных вирусов, из которых несколько сотен инфицируют людей и животных.
Международным комитетом по таксономии вирусов (ICTV, 2000) разработана единая система классификации и номенклатуры вирусов, которая основана на изучении различных свойств вирусов, таких как размер, морфология и структура вириона, его устойчивость к воздействию различных факторов и антигенные свойства.
Классификация вирусов
На сегодня главным критерием в таксономии вирусов являются: 1) тип и структура вирусного генома 2) стратегия репликации вируса 3) структура вириона. Полное или частичное секвенирование вирусного генома увеличивает таксономическую информацию и очень часто используется с целью идентификации вируса.
Сведения о нуклеотидной последовательности для всех таксономических групп вирусов имеются в опубликованной базе данных (Генбанк, Национального Центра биотехнологической информации, Национальной медицинской лаборатории, Национального Института здоровья, Бетезда, Мериленд, США). Это достижение в большинстве случаев укорачивает таксономическую идентификацию, хотя традиционные методы все еще часто используются по экономическим соображениям. Универсальная система таксономии вирусов включает несколько уровней: порядок, семейство, подсемейство, род и вид. Патогенные вирусы позвоночных (человека и животных) в соответствии с современной системой классификации вирусов объединены в 2 (Mononegavirales и Nidovirales) порядка и 28 семейств, из которых 10 являются ДНК-вирусами и 18 РНК-вирусами.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вирусы оспы. Виды вирусов оспы. Строение вирусов оспы.
Вирусы оспы (ВО) — самые крупные вирусы, содержащие ДНК, молекулярная масса которой больше, чем у любого другого вируса животных. Они широко распространены в природе, вызывают заболевания позвоночных и беспозвоночных с летальным исходом или в виде легко протекающей продолжительной инфекции с образованием доброкачественных опухолей. ВО передаются членистоногими при прямом контакте, аэрозольно или механически. ВО обычно имеют узкий круг хозяев, хотя некоторые из них представляют исключения, в том числе вирусы вакцины и оспы птиц. Вирусы оспы позвоночных включают восемь родов (орто-, пара-, ави-, капри-, лепори-, суи-, моллюсци- и ятапоксвирусы).
Разделение вирусов оспы позвоночных на роды проведено с учетом массы и структуры генома, размера и формы вириона, способности к генетической рекомбинации, антигенной связи и спектра патогенности.
Ортопоксвирусы. Типичный представитель - вирус осповакцины (ВОВ). Спектр естественных хозяев узкий, обычно ограничен одним видом животных: вирусы агглютинируют эритроциты цыплят; варьируют по вирулентности для естественных хозяев и лабораторных животных. Другие виды: вирусы оспы буйволов, верблюдов, крупного рогатого скота, мышей, обезьян и вирус натуральной оспы.
Парапоксвирусы. Типичный представитель — вирус контагиозного пустулёзного дерматита (эктимы) овец и коз (вирус орф). Представители рода серологически связаны между собой, но отличаются от представителей других родов; гемагглютинин не образуют. Другие виды: вирусы пустулёзного стоматита крупного рогатого скота, контагиозной эктимы серн и сыпи доярок.
Авипоксвирусы. Естественные хозяева — птицы; типичный представитель — вирус оспы кур. Члены рода серологически связаны между собой. Другие виды: вирусы оспы канареек, голубей, перепелов, воробьев, скворцов, индеек и оспы Юнко. Передаются членистоногими.
Каприпоксвирусы. Естественные хозяева — парнокопытные; типичный представитель — ВО мелких жвачных (овец и коз). В данный род входит также вирус кожной бугорчатки крупного рогатого скота.
Лепорипоксвирусы. Типичный представитель — вирус миксомы кроликов. Передается механически членистоногими. Другие виды: вирусы фибромы зайцев, кроликов (вирус Шоупа) и белок. Вирус злокачественной фибромы кроликов — летальный туморогенный поксвирус, возникший, по-видимому, вследствие рекомбинации между вирусами фибромы и миксомы кроликов.
Суипоксвирусы. Типичный представитель — вирус оспы свиней. В инфицированных клетках образуются несколько типов цитоплазматических включений и наблюдается вакуолизация ядра. Другие члены рода не определены.
Моллюсципоксвирусы. Типичный представитель — вирус контагиозного моллюска. Возможные члены рода — вирусы однокопытных и обезьян.
Ятапоксвирусы включают вирус опухолей обезьян Яба и вирус оспы Тана.
Вирусы оспы — наиболее крупные из всех вирусов животных. Под электронным микроскопом они выглядят как большие овальные (кирпичеобразные) частицы размером около 250-350x200-270 нм.
Орто-, ави-, лепорипоксвирусы более вытянуты, а вирус оспы свиней шире, чем другие ВО. Парапоксвирусы имеют овальную (коконообразную) форму и размер 260x160 нм.
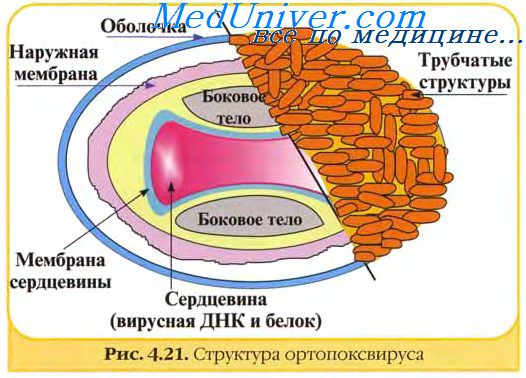
В структуре вирусов оспы различают три основных компонента: двояковогнутую сердцевину, овальные боковые тела и оболочку вириона. Сердцевину вириона составляют ДНК и связанные с нею белки. Сердцевина окружена гладкой мембраной (толщиной около 5 нм), снаружи покрытой слоем вертикально уложенных и плотно прилегающих друг к другу цилиндрических субъединиц (5x10 нм). Вогнутость сердцевины с обеих сторон занята овальными образованиями (неизвестной природы), называемыми боковыми телами. Они как бы сдавливают сердцевину, придавая ей форму двояковогнутого диска, имеющего на разрезе вид гантели.
Вирионы заключены в липопротеиновую супероболочку, которая имеет толщину 20—30 нм и содержит липиды клетки и вирусспецифические белки.
Вирионы большинства вирусов оспы окружены слоем беспорядочно расположенных трубчатых структур, придающих им характерный вид. Эти структуры состоят из сферических субъединиц диаметром около 5 нм. Субъединицы построены из молекул протеина или гликопротеина. В состав ворсинок длиной 20 нм покрывающих поверхность вируса осповакцины (ВОВ), входит белок с молекулярной массой 58 кД, относящийся к главным полипептидам вириона. Поверхность парапоксвирусов покрыта длинными нитеподобными трубочками, уложенными крест-накрест, напоминающими клубок пряжи.
Вирус осповакцины содержит белки, липиды и ДНК, которые соответственно составляют 90, 5 и 3,2% массы вириона (5х10~15 г). В вирусе оспы птиц около 1/3 массы составляют липиды.
Вирион образуется включением ДНК внутрь незрелых вирусоподобных частиц, которые затем созревают, покрываясь дополнительно наружными оболочками. Репликация и сборка вирионов происходят в разных местах цитоплазмы (в виропластах или вирусных фабриках), и вирионы освобождаются почкованием или при лизисе клеток.
Вирионы, освободившиеся из клетки почкованием до ее разрушения, покрыты оболочкой, которая содержит клеточные липиды и несколько вирусспецифических белков.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Но и Дженнер не имел представления о том, что является причиной заболевания оспой. В XIX веке все болезнетворные организмы и вещества без разбора называли вирусами. Лишь благодаря опытам отечественного биолога Дмитрия Иосифовича Ивановского прекратилась эта путаница! Он пропускал экстракт заражённых табачной мозаикой 1 растений через бактериальные фильтры, сквозь которые не проходят даже самые мелкие бактерии. Выяснилось, что экстракт оставался по-прежнему заразным для других растений. Значит, возбудителями табачной мозаики были организмы, меньшие по размеру, чем бактерии; их назвали фильтрующимися вирусами. Вскоре бактерии перестали называть вирусами, а сами вирусы выделили в отдельное царство живых организмов. Дмитрий Ивановский же во всём мире по праву считается основателем вирусологии — науки о вирусах.

Рис. 2. Дженнер прививает Джеймса Фиппса от оспы
Но что мы пока поняли про вирусы? Только то, что они меньше бактерий. Чем же вирусы так не похожи на другие организмы? И почему понадобилось вдруг их выделять в отдельное царство? А вот почему. В отличие от других живых организмов, вирусы не имеют клеточного строения, а значит, и всех характерных для клетки структур. А ещё они единственные, кто не умеет самостоятельно производить белок, главный строительный материал всего живого. Поэтому их размножение невозможно вне заражённой клетки. Из-за этого многие учёные не без оснований считают вирусы внутриклеточными паразитами.
Жертвами различных вирусов становятся представители всех без исключения существующих царств живых организмов! Так, есть вирусы растений — вирус табачной мозаики (рис. 3, слева), вирус мозаики костра (это растение изображено на рисунке 3, справа), вирус желтухи свёклы, вызывающий иногда даже эпидемии. Кстати, в растение вирус просто так не проникнет. Заражение происходит при травмах растительных тканей. Типичный пример: тля пьёт сок из стебля и для этого протыкает покровные ткани — а вирус тут как тут.

Рис 3. Слева: листья табака, поражённые вирусом табачной мозаики. Справа: костёр (лат. Brómus) — род многолетних травянистых растений семейства Злаки. Если посмотреть на заросли костра в ветреную погоду, его крупные метёлки, склоняясь под ветром то в одну, то в другую сторону, отсвечивают красноватым светом в солнечных лучах, очень напоминая языки пламени. Отсюда, вероятно, и произошло русское название этого растения
Грибы тоже поражаются вирусами, вызывающими, например, побурение плодовых тел у шампиньонов или изменение окраски у зимнего опёнка. Причиной многих опасных заболеваний животных и человека тоже служат вирусы: вирус гриппа, ВИЧ (вирус иммунодефицита человека), вирус Эбола, вирус бешенства, герпеса, клещевого энцефалита и т. д.
Есть даже вирусы, поражающие бактерии, их называют бактериофагами 2 . Так, в конце XIX века исследователи из Института Пастера заметили, что вода некоторых рек Индии обладает бактерицидным действием, то есть способствует снижению роста бактерий. И достигалось это благодаря присутствию в речной воде бактериофагов.

Рис. 4. Слева: вирус табачной мозаики. В центре: вирус мозаики костра похож на футбольный мяч (справа)

Рис. 5. Слева направо: вирус герпеса, аденовирус А человека, бактериофаг

Рис. 6. Маленькие вирусы-спутники внутри гигантского мимивируса
Но не стоит думать, что вирусы причиняют исключительно вред другим организмам! Так, исследователи из Пенсильванского университета показали, что безвредный для человека вирус AAV2, встречающийся почти у всех людей, убивает самые разные виды раковых клеток. При этом здоровые клетки организма вирус не заражает.
А совсем недавно стало известно, что вирусы тоже болеют. Мимивирус, поражающий амёбу Acanthamoeba polyphaga, сам страдает от другого вируса-спутника (рис. 6). Он, кстати, так и называется — Спутник. Этот вирус-спутник использует механизмы воспроизводства мимивируса для собственного размножения, мешая ему нормально развиваться в клетке амёбы. По аналогии с бактериофагами, он был назван вирофагом, то есть пожирающим вирусы. Можно сказать, что присутствие вируса-спутника в амёбе обеспечивает ей больше шансов на выживание в борьбе с мимивирусом.
Читайте также: