
Способ размножения вирусов дизъюнктивный
Обновлено: 25.04.2024
При данном типе размножения вирион, попав в клетку хозяина перестает обнаруживаться, нуклеиновая кислота освобождается от оболочки (капсида суперкапсида). Начинается синтез компонентов вириона, т.е его репродукция. Она носит ДИЗЪЮНКТИВНЫЙ характер поскольку компоненты вириона синтезируются в разных частях клетки: белки на рибосомах, нуклеиновые кислоты в ядре или цитоплазме. Вирус использует для этого генетический аппарат клетки, подавляя необходимые ей самой синтетические реакции. Затем после синтеза необходимых компонентов происходит сборка вириона (а точнее нескольких вирионов). Из разных структур клетки происходит транспортировка частей вириона в одно место сборки. При этом вирусные белки и нуклеиновые кислоты имеют свойство узнавать, самопроизвольно соединяться друг с другом. Многие простые вирионы имеют собираются на репликативных комплексах – мембранах ЭПР. У сложных вирионов сборка нуклеокапсида начинается на репликативных комплексах и продолжается на плазматической мембране клетки хозяина, с наружной стороны которой располагаются суперкапсидные гликопротеиды. Затем гликопротеидные и примыкающие к ним с другой стороны нуклеокапсидные участки образуют выпячивание на поверхности клетки виде почки. После отделения почки содержащий нуклеокапсид и суперкапсидные белки образуются свободные вирионы.
13 Взаимодействие вируса с клеткой. Продуктивный и непродуктивный типы инфекций.
13)Взаимодействие вируса с клеткой хозяина
Взаимодействие вируса с клеткой хозяина - это сложный «ногоступенчатый процесс, который начинается с адсорбции вирусных частиц на рецепторах клетки хозяина и продолжается после их проникновения внутрь клетки. В результате такого взаимодействия мовивается либо продуктивная, либо абортивная, либо интегративная форма клеточной инфекции. При продуктивной форме происходит размножение, точнее, репродукция (лат. reproduce - воспроизводить) вируса, при абортивной - ее нарушение на одном из этапов, при интегративной - интеграция вирусной нуклеиновой кислоты в клеточный геном.
14 Характеристика продуктивного типа инфекции
Продуктивный тип: Вирус проникает в клетку и даёт в клетке потомство. Это может быть острая вирусная инфекция (Образуется сразу тысячи новых вирионов. Клетка погибает. Происходит аутоинтаксикация) и хроническая вирусная инфекция( В клетке образуются единицы вирусных вибрионов. Клетка разрушается постепенно в течении времени)
Непродуктивный тип: Нет вирусного потомства(явного) Варианты непродуктивной инфекции:
- Вирус заходит в клетку и остаётся в цитоплазме, но не разрушается, иногда он может активироваться и приводить к хронической инфекции
-Вирус сразу внедряется в геном клетки или ч/з некоторое время. Продукции вируса нет. Многие вирусы обладают способностью к трансформации клетки за счёт того что внедряются в определённые участки генома( опухолевые участки)
-Персистирующие вирусы. Эти вирусы попав в организм не вызывают ни каких симптомов, сохраняясь в определённых клетках(пр: вирус герпеса)
Для вирусов характерен дизъюнктивный (от disjuncus — разобщенный) способ репродукции-размножения. Потомство вируса возникает в результате сборки нуклеиновых кислот и белковых субъединиц, которые синтезируются раздельно клеткой хозяина.
Проникновение вируса в клетку и воспроизведение себе подобных проходит в несколько фаз:
1.проникновение в клетку хозяина,
2.синтез ферментов, необходимых для репликации вирусных нуклеиновых кислот,
3.синтез вирусных частей,
4.сборка и композиция зрелых вирионов,
5.выход зрелых вирионов из клетки.
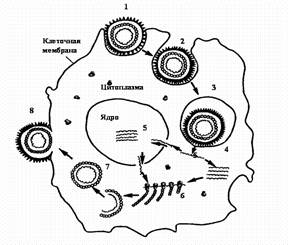
Стадии репродукции вирусов.
1 - адсорбция вириона на клетке; 2 - проникновение вириона в клетку путем виропексиса;
3 - вирус внутри вакуоли клетки; 4 - `раздевание вириона вируса; 5 - репликация вирусной нуклеиновой кислоты; 6 - синтез вирусных белков на рибосомах клетки; 7 - формирование вириона; 8 - выход вириона из клетки путем почкования.
Фаза I — адсорбция вириона на поверхности клетки.
Протекает в две стадии: первая — неспецифическая, когда вирус удерживается на поверхности клетки при помощи электростатических сил, т. е. благодаря возникновению противоположных зарядов между отдельными участками мембраны клеток и вируса. Эта фаза взаимодействия вируса с клеткой обратима, на нее оказывают влияние такие факторы, как рН и солевой состав среды.
Вторая стадия — специфическая, когда взаимодействуют специфические рецепторы вируса и рецепторы клетки, комплементарные друг другу. По химической природе рецепторы клетки могут быть мукопротеидами (или мукополисахаридами) и липопротеидами. Разные вирусы фиксируются на разных рецепторах: вирусы гриппа, парагриппа, аденовирусы — на мукопротеидах, а вирусы клещевого энцефалита, полиомиелита — на липопротеидах.
Фаза II — проникновение вируса в клетку. Электроноскопические наблюдения за процессом проникновения вирусов в чувствительные к ним клетки показали, что оно осуществляется посредством механизма, напоминающего пиноцитоз, или, как чаще называют, виропексис. В месте адсорбции вируса клеточная стенка втягивается внутрь клетки, образуется вакуоль, в которой оказывается вирион. Параллельно клеточные ферменты (липазы и протеазы) вызывают депротеинизацию вириона — растворение белковой оболочки и освобождение нуклеиновой кислоты.
Фаза III — скрытый период (период эклипса — исчезновения). В этот период в клетке невозможно определить наличие инфекционного вируса ни химическими, ни электронно-микроскопическими, ни серологическими методами. О сущности этого явления и его механизмов пока известно мало. Предполагается, что в скрытой фазе нуклеиновая кислота вируса проникает в хромосомы клетки и вступает с ними в сложные генетические взаимоотношения.
Фаза IV — синтез компонентов вириона. В этой фазе вирус и клетка представляют единое целое, вирусная нуклеиновая кислота выполняет генетическую функцию, индуцирует образование ранних белков и изменяет функцию рибосом. Ранние белки подразделяются на:
а) белки-ингибиторы (репрессоры), подавляющие метаболизм клеток
б) белки-ферменты (полимеразы), обеспечивающие синтез вирусных нуклеиновых кислот.
Синтез нуклеиновых кислот и белков протекает неодновременно и в разных структурных частях клетки. У вирусов, содержащих ДНК или РНК, эти процессы имеют некоторые различия и особенности.
Процесс формирования вирионов начинается спустя определенное время после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность этого периода довольно вариабельна и предопределяется природой вируса — для РНК-содержащих обычно короче, чем для ДНК-вирусов. Например, продукция полных вирусных частиц осповакцины начинается приблизительно спустя 5—6 ч после инфицирования клеток и продолжается в течение последующих 7—8 ч, т. е. после того как синтез вирусной ДНК уже завершен.

Между нуклеиновой кислотой и соответствующим белковыми субъединицами образуются очень прочные связи, о чем свидетельствуют трудности отделения белка от вирусной нуклеиновой кислоты. Большую прочность вирусной частице придают входящие в ее состав углеводы и особенно липиды.
Формирование вирионов, так же как и синтез компонентов вируса, происходит в разных местах клетки, при участии различных клеточных структур. После завершения процесса формирования образуется зрелая дочерняя вирусная частица, обладающая всеми свойствами родительского вириона. Но иногда наблюдается образование так называемых неполных вирусов, которые состоят или только из нуклеиновой кислоты, или из белка, или из вирусных частиц, формирование которых остановилось в какой-то промежуточной стадии.
Фаза VI — выход зрелых вирионов из клетки. Существуют два основных механизма выхода зрелых вирионов из клетки:
1) выход вириона с помощью почкования. В этом случае наружная оболочка вириона происходит из клеточной мембраны, она содержит как материал клетки хозяина, так и вирусный материал;
2) выход зрелых вирионов из клетки через бреши в мембране. Эти вирусы не имеют наружной оболочки. При таком механизме выхода вирусов клетка, как правило, погибает и в среде появляется большое количество вирусных частиц.
Причиной гибели зараженной клетки могут быть три механизма:
2.защитная реакция клетки, запускающая генетическую программу ее гибели (апоптоз);
3. иммунная система организма, уничтожающая зараженную клетку.
Кроме продуктивного типа взаимодействия вируса и клетки возможно интегративное сосуществование или вирогения. Вирогения характеризуется интеграцией (встраиванием) нуклеиновой кислоты вируса в геном клетки, а также репликацией и функционированием вирусного генома как составной части генома клетки. Для интеграции с клеточным геномом необходимо возникновение кольцевой формы двунитевой ДНК вируса. Встроенная в состав хромосомы клетки вирусная ДНК называется провирусом. Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. Под влиянием некоторых физических или химических факторов провирус может переходить в автономное состояние с развитием продуктивного типа взаимодействия с клеткой. Дополнительная генетическая информация провируса при вирогении сообщает клетке новые свойства, что может быть причиной развития опухолей, аутоиммунных и хронических заболеваний. На способности вирусов к интеграции с геномом клетки основаны персистенция (от лат. persisto - постоянно пребывать, оставаться) вирусов в организме и развитие персистентных вирусных инфекций. Например, вирус гепатита В способен вызывать персистирующие поражения с развитием хронического гепатита и часто опухолей печени.
Специфические рецепторы клеток имеют различную природу, являясь белками, липидами, углеводными компонентами белков, липидов и др. Так, рецепторами для вируса гриппа является сиаловая кислота в составе гли-копротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Вирусы бешенства адсорбируются на ацетилхолиновых рецепторах нервной ткани, а вирусы иммунодефицита человека — на СО4-рецепторах Т-хелперов, моноцитов и дендритных клеток. На одной клетке находится от десяти до ста тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов.
Наличие специфических рецепторов лежит в основе избирательности поражения вирусами определенных клеток, тканей и органов. Это так называемый тропизм(греч. tropos — поворот, направление). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках — нейротропными, в иммунокомпетентных клетках — иммунотропными и т. д.
Проникновение вирусов в клетку.Вирусы проникают в клетку путем рецептор-зависимого эндоцитоза (виропексиса), или слияния оболочки вируса с клеточной мембраной, или же в результате сочетания этих механизмов.
2. Слияние обточки вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (парамиксовирусов, ретровиру-сов, герпесвирусов), в составе которых имеются белки слияния. Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, в результате чего вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль.
В) Синтез вирусных компонентов. Синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве. Синтез осуществляется в разных частях клетки, поэтому такой способ размножения вирусов называется дизъюнктивным (от лат. disjunctus — разобщенный).
С)Синтез вирусных белков. В зараженной клетке вирусный геном кодирует синтез двух групп белков:
1. неструктурных белков, обслуживающих внутриклеточную репродукцию вируса на разных его этапах;
2. структурных белков, которые входят в состав вириона (геномные, связанные с геномом вируса, капсидные и су-перкапсидные белки).
К неструктурным белкамотносятся: 1) ферменты синтеза РНК или ДНК (РНК- или ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; 2) белки-регуляторы; 3) предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; 4) ферменты, модифицирующие вирусные белки, например, протеиназы и протеинкиназы.
I. ДНК-содержашие вирусы реализуют генетическую информацию так же, как и клеточный геном, по схеме:
Причем ДНК-содержашие вирусы используют для этого процесса клеточную полимеразу (вирусы, геномы которых транскрибируются в ядре клетки — аденовирусы, па-повавирусы, герпесвирусы) или собственную РНК-полимеразу (вирусы, геномы которых транскрибируются в цитоплазме, например поксвирусы).
II. Плюс-нитевые РНК-содержашие вирусы(например, пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняющий функцию иРНК; он распознается и транслируется рибосомами. Синтез белков у этих вирусов осуществляется без акта транскрипции по схеме:
геномная РНК вируса-> трансляция белка вируса.
III. Геном минус-однонитевых РНК-содержаших вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов) и двунитевых (реовирусов) служит матрицей, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме:
IV. Ретровирусы(вирусы иммунодефицита человека, онкогенные ретровирусы) имеют уникальный путь передачи генетической информации. Геном ретровирусов состоит из двух идентичных молекул РНК, т. е. является диплоидным. В составе ретровирусов есть особый вирусоспецифический фермент — обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т. е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК (кДНК). Комплементарная нить ДНК копируется с образованием двунитевой комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков для этих вирусов осуществляется по схеме:
Репликация вирусных геномов,т. е. синтез вирусных нуклеиновых кислот, приводит к накоплению в клетке копий исходных вирусных геномов, которые используются при сборке вирионов. Способ репликации генома зависит от типа нуклеиновой кислоты вируса, наличия вирусоспецифических или клеточных полимераз, а также от способности вирусов индуцировать образование полимераз в клетке.
Механизм репликации отличается у вирусов, имеющих:
1) двунитевую ДНК;
2) однонитевую ДНК;
3) плюс-однонитевую РНК;
4) минус-одноните-вую РНК;
5) двунитевую РНК;
6) идентичные плюс-нитевые РНК (ретровирусы).
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас-
плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая
группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
2.Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы
используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед
него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
3.Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК
полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты:
РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным
репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны
для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
4.Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус-
нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
5.Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей
РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Формирование вирусов.Вирионы формируются путем самосборки: составные части вириона транспортируются в места сборки вируса — участки ядра или цитоплазмы клетки. Соединение компонентов вириона обусловлено наличием гидрофобных, ионных, водородных связей и стерического соответствия.
Существуют следующие общие принципы сборки вирусов:
Формирование вирусов— многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по
составу полипептидов.
□ Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов.
□ У сложноустроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса).
Причем сборка вирусов, реплицирующихся в ядре клетки, происходит с участием мембраны ядра, а сборка вирусов, репликация которых идет в цитоплазме, осуществляется с
участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса.
□ У ряда сложноустроенных вирусов минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается так называемый матриксный белок (М-белок), который расположен под модифицированной клеточной ембраной. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и вирусной липопротеиновой оболочкой.
□ Сложноустроенные вирусы в процессе формирования включают в свой состав некоторые компоненты клетки хозяина, например липиды и углеводы.
Выход вирусов из клетки.Полный цикл репродукции вирусов завершается через 5—6 ч (вирус гриппа и др.) или через несколько суток (гепатовирусы, вирус кори и др.). Процесс репродукции вирусов заканчивается выходом их из клетки, который происходит взрывным путем или почкованием, экзоцитозом.
□ Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки.
□ Почкование, экзоцшпт присущи вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида или сердцевины вириона с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. При этом клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (например, буньявирусы).
Вирусы, формирующиеся в ядре клетки (например, герпесвирусы), почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазма-тических везикул на поверхность клетки.
По строению различают 2 типа вирусных частиц: простые и сложные.
Внутренняя структура простых и сложных вирусов сходна.
Сердцевина вируса - вирусная нуклеиновая кислота вирусный геном. Вирусный геном может быть представлен одной из 4 молекул РНК или ДНК: однонитчатыми и двунитчатыми РНК и ДНК. Большинство вирусов имеют один цельный или фрагментированный геном, имеюший линейную или замкнутую форму. Однонитчатые геномы могут иметь 2 полярности: 1) позитивную, когда вирионная нуклеиновая кислота одновременно служит и матрицей для синтеза новых геномов и выполняет роль и-РНК; 2) негативную, выполняющую только функцию матрицы. Геном вирусов содержит от 3 до 100 и более генов, которые делятся на структурные, кодирующие синтез белков, входящих в состав вириона, и регуляторные, которые изменяют метаболизм клетки хозяина и регулируют скорость размножения вирусов.
Ферменты вирусов также закодированы в геноме. К ним относятся: РНК-зависимая РНК-полимераза (транскриптаза), которая обнаружена у всех РНК-содержащих вирусов с негативной полярностью. Поксвирусы содержат ДНК-зависимую РНК-полимеразу. Ретровирусы имеют уникальный фермент - РНК-зависимую ДНК-полимеразу, называемую обратной транскриптазой. В геноме некоторых вирусов имеются гены, кодирующие РНК-азы, эндонуклеазы, проте-инкииазы.
Снаружи нуклеиновая кислота покрыта белковым чехлом - капсидом, образуя комплекс - нуклеокапсид (в химическом смысле - нуклеопротеид). Кап-сид состоит из отдельных белковых субъединиц - капсомеров, которые представляют уложенную определённым образом полипептидную цепь, создающую симметричную конструкцию. Если капсомеры укладываются по спирали, такой тип укладки капсида носит название спиральной симметрии. Если капсомеры укладываются по граням многогранника (12-20-гранника), такой тип укладки капсида носит название икосаэдрической симметрии
Капсид представлен a-спиральными белками, способными к полимеризации, которые выполняют защиту генома от различных воздействий, выполняют рецепторную функцию у этой группы вирусов, обладают антигенными свойствами.
Сложные вирусы имеют внешнюю оболочку - суперкапсид, расположенную поверх капсида. В состав суперкапсида входят внутренний белковый слой - М-белок, затем более объёмный слой липидов и углеводов, извлечённых из клеточных мембран клетки-хозяина. Вирусспецифические гликопротеиды проникают внутрь суперкапсида, образуя снаружи фигурные выпячивания, которые выполняют рецепторную функцию. Вирусы существуют в трёх формах:
1) вирион (вирусная частица) - внеклеточная форма;
2) внутриклеточный (вегетативный) вирус;
3) интегрированный с ДНК хозяина вирус (провирус).
Взаимодействие вируса с клеткой. Репродукция (размножение) вирусов
Вирусы - облигатные внутриклеточные паразиты, способные размножаться только в живой клетке. В отличие от прокариотических и эукариотических микроорганизмов вирусы не размножаются бинарным делением. Размножение вирусов происходит путём репродукции (англ, "reproduce" - воспроизво-аить, делать копию), то есть воспроизведение их нуклеиновых кислот и белков z последующей сборкой вирионов. Синтез нуклеиновых кислот и белков вируса происходит в разных частях клетки (ядре и цитоплазме). Такой способ репродукции получил название дизъюнктивного (разобщённого).
Процесс репродукции вирусов условно можно разделить на 2 фазы. Первая фаза включает 3 стадии: 1) адсорбцию вируса на чувствительных клетках; 2) проникновение вируса в клетку; 3) депротеинизацию вируса. Вторая фаза включает стадии реализации вирусного генома: 1) транскрипцию, 2) трансляцию, 3) репликацию, 4) сборку, созревание вирусных частиц и 5) выход вируса из клетки.
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. с прикрепления вируса к поверхности клетки.
Адсорбция представляет собой специфическое связывание вирионного белка (антирецептора) с комплементарной структурой клеточной поверхности - клеточным рецептором. По химической природе рецепторы, на которых фиксируются вирусы, относятся к двум группам: мукопротеидным и липопротеидным. Вирусы гриппа, парагриппа, аденовирусы фиксируются на мукопротеидных рецепторах. Энтеровирусы, вирусы герпеса, арбовирусы адсорбируются на липопротеидных рецепторах клетки. Адсорбция происходит лишь при наличии определённых электролитов, в частности ионов Са2+, которые нейтрализуют избыточные анионные заряды вируса и клеточной поверхности и уменьшают электростатическое отталкивание Адсорбция вирусов мало зависит от температуры Начальные процессы адсорбции носят неспецифический характер, являются результатом электростатического взаимодействия положительно и отрицательно заряженных структур на поверхности вируса и клетки, а затем наступает специфическое взаимодействие прикрепительного белка вириона со специфическими группировками на плазматической мембране клетки. Простые вирусы человека и животных содержат прикрепительные белки в составе капсида. У сложно организованных вирусов прикрепительные белки входят в состав супер-капсида. Они могут иметь форму нитей (фибры у аденовирусов), либо шипов, грибоподобных структур у миксо-, ретро-, рабдо- и других вирусов. Вначале происходит единичная связь вириона с рецептором - такое прикрепление непрочное - адсорбция носит обратимый характер. Чтобы наступила необратимая адсорбция, должы появиться множественные связи между рецептором вируса и рецептором клетки, т. е. стабильное мультивалентное прикрепление. Количество специфических рецепторов на поверхности одной клетки составляет 10 4 -10 5 . Рецепторы для некоторых вирусов, например, для арбовирусов. содержатся на клетках как позвоночных, так и беспозвоночных, для других вирусов только на клетках одного или нескольких видов.
Проникновение вирусов человека и животных в клетку происходит двумя путями: 1) виропексисом (пиноцитозом); 2) слиянием вирусной суперкапсидной оболочки е клеточной мембраной. Бактериофаги имеют свой механизм проникновения, так называемый шприцевои, когда в результате сокращения белкового отростка фага нуклеиновая кислота как бы впрыскивается в клетку.
Депротеинизация вируса освобождение геиома вируса от вирусных защитных оболочек происходит либо с помощью вирусных ферментов, либо с помощью клеточных ферментов. Конечными продуктами депротеинизации являются нуклеиновые кислоты или нуклеиновые кислоты, связанные с внутренним вирусным белком. Затем имеет место вторая фаза вирусной репродукции, ведущая к синтезу вирусных компонентов.
Транскрипция - переписывание информации с ДНК или РНК вируса на и-РНК по законам генетического кода.
Трансляция - процесс перевода генетической информации, содержащейся в и-РНК, на специфическую последовательность аминокислот.
Репликация - процесс синтеза молекул нуклеиновых кислот, гомологичных вирусному геному.
Реализация генетической информации у ДНК-содержащих вирусов идёт так же, как и в клетках:
ДНК транскрипция и-РНК трансляция белок
У РНК-содержащих вирусов с негативным геномом (вирусы гриппа, пара-гриппа и др.) формула реализации генома следующая:
-РНК транскрипция и-РНК трансляция белок
У вирусов с позитивным РНК-геномом (тогавирусы, пикорнавирусы) транскрипция отсутствует:

+РНК трансляция белок
У ретровирусов - уникальный путь передачи генетической информации:
РНК обратная транскрипция ДНК транскрипция и-РНК трансляция белок
ДНК интегрируется с геномом клетки-хозяина (провирус).

Репродукция вируса – это процесс размножения вирусных частиц в чувствительных к ним клетках. Репродуцируются только вирулентные вирусы, обладающие высокой степенью патогенности [3] .
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций [3] .
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина [2] .
- Все вирусы, содержащие молекулу ДНК, кроме вирусаоспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирусоспы размножается только в цитоплазме клетки [2] .
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами [2] .
- Белки вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению) [2] .
- Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование (присоединении к полипептиду углеводных остатков) [2] .

Этапы репродукции (жизненного цикла) вируса

1. Адсорбция вируса на мембране клетки.; 2. Проникновение вируса в клетку.; 3. Депротеинизация.; 4. Синтез компонентов вирусов.; 5. Формирование дочерних вирионов.; 6. Выход вирионов [2] .
Этапы репродукции вируса
Репродукцию или жизненный цикл вируса делят на шесть последовательных этапов:
- Адсорбция на мембране клетки [2] .
- Проникновение в клетку [2] .
- Депротеинизация [2] .
- Синтез компонентов вирусов[2] .
- Формирование дочерних вирионов[2] .
- Выход вирионов[2] .

Процесс адсорбции вириона на поверхности клетки

1. Вирион.; 2. Клеточный рецептор.; 3. Прикрепительный белок; 4. Клеточная мембрана [2] .
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона [2] .
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур [2] .
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно [2] .
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 [2] .

Проникновение вируса в клетку (Путь I)

Слияние вирусной оболочки с клеточной мембраной
1. Вирион.; 2. Инфицируемая клетка.; 3. Ядро [2] .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона [2] . Существует два пути:
- Путь I – слиянии вирусной оболочки с клеточной мембраной [2] .
- Путь II – рецептор-опосредованный эндоцитоз [2] .
Путем слияния суперкапсида с клеточной мембраной (путь I) в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки [2] .
Путем рецептор-опосредованного эндоцитоза (путь II) в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении, вирусный геном в цитоплазме клетки освобождается из эндосомы [2] .

Проникновение вируса в клетку (Путь II)

1. Вирион.; 2. Клетка.; 3. Эндосома.; 4. Ядро [2] .
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы) [2] .
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона [2] .
Синтез компонентов вируса
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [1] .
Место синтеза компонентов дочерних вирионов зависит от типа генома:
- реализация генетической информации у ДНК-содержащих вирусов идет по пути: ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом идет по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок;
- РНК-содержащие ретровирусы идут по следующему пути передачи информации: РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок [2] .
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона [2] .
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках [2] .
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК [2] .
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается [2] .
РНК-содержащие ретровирусы отличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза [2] [4] .
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков, и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры [2] .
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более [2] .
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования[2] .
Путь лизиса клетки тесно связан ее деструкцией. Он характерен для безоболочечных вирусов, не имеющих суперкапсидной оболочки (суперкапсида) [2] .
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы, высвобождаются в течении 2–6 часов. В начале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида [2] .
Читайте также: