
Способы реализации генетической информации вирусов
Обновлено: 18.04.2024
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ГЕНОМА ВИРУСА
Вирусы являются одним из излюбленных объектов молекулярной генетики благодаря простому строению и малой молекулярной массе их геномов, которая в 10 6 раз меньше массы генома эукариотической клетки. Организация генетического аппарата у ряда вирусов, например у SV40, настолько сходна с таковой генов эукариотической клетки, что получила название минихромосомы. Минихромосома широко используется для изучения организации и репликации ДНК.
Число генов у вирусов значительно варьирует: от 3—4 генов у просто устроенных вирусов (парвовирусы) до 150 генов и больше у сложно устроенных (вирус оспы). Геном вирусов животных является гаплоидным, за исключением ретровирусов, которые имеют диплоидный геном, представленный двумя идентичными молекулами РНК. У вирусов с фрагментарным геномом (вирусы гриппа, реовирусы) каждый фрагмент обычно представляет собой один ген.
В составе генов ДНК-содержащих вирусов есть регуляторные участки, в том числе промотор, контролирующие функцию структурных генов. Сильными промоторами являются концы многих вирусных ДНК, представляющие собой длинные концевые повторы, сильный промотор имеют гены тимидинкиназы вирусов оспы и герпеса. Эти промоторы используются в генной инженерии для усиления транскрипции изучаемого гена.
СПОСОБЫ УВЕЛИЧЕНИЯ ИНФОРМАЦИОННОЙ ЕМКОСТИ ВИРУСНОГО ГЕНОМА
У многих вирусов молекулярная масса синтезирующихся белков превышает теоретически рассчитанную. Этот феномен объясняется наличием у вирусов механизмов, позволяющих получить развернутую генетическую информацию при максимальной экономии генетического материала; подобные механизмы выработаны в процессе эволюции вирусов как генетических паразитов.
Способами увеличения генетической информации являются: 1) двукратное считывание одной и той же иРНК, но с другого инициирующего кодона; 2) сдвиг рамки трансляции; 3) сплайсинг; 4) транскрипция с перекрывающихся областей ДНК и др.

Трансляция может происходить без сдвига рамки и со сдвигом рамки. Генетический код является триплетным, это означает, что три нуклеотида, составляющих триплет, или кодон, кодируют одну аминокислоту. В том случае, если триплеты сохранены и генетический код не изменился, то при трансляции с двух разных инициирующих кодонов будут синтезироваться полипептиды, представляющие собой укороченную копию первого полипептида (трансляция без сдвига рамки).
В том случае, если произошел сдвиг на один или два нуклеотида, образуются новые триплеты (кодоны) и появляется новый генетический код. В этом случае одна молекула иРНК может транслироваться с образованием двух уникальных белков, т. е. таких белков, у которых нет идентичных аминокислотных последовательностей.
Одним из способов экономии генетического материала является нарезание полипептида-предшественника на участки разной длины, в результате чего образуются разные полипептиды с перекрывающимися аминокислотными последовательностями. Подобный механизм нарезания имеет место у аденоассоциированных вирусов и у SV40.
Таким образом, число реальных генов превосходит молекулярную массу генома. Основанный на длине генома расчет числа генов неизменно приведет к ошибочным результатам. Более точные представления о числе генов можно получить путем биохимического и генетического анализов.
ОСНОВНЫЕ ПРОЦЕССЫ, КОНТРОЛИРУЮЩИЕ НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ ВИРУСОВ
Модификации. Модификациями называются не наследуемые (фенотипические) изменения у вирусов, обусловленные клеткой-хозяином. Эти изменения лежат в основе адаптации вируса к новому хозяину и преодоления зависимого от хозяина ограничения. Модификации нуклеиновых кислот вирусов осуществляют клеточные ферменты, ответственные за ограничение (рестрикцию) репродукции вируса.
Мутации. В основе изменчивости вирусов лежат мутации, т. е. изменения состава и последовательностей нуклеотидов вирусного генома. Мутации происходят у всех вирусов, независимо от того, является ли их генетическим аппаратом ДНК или РНК. В результате мутаций отдельные вирионы могут приобретать новые свойства. Дальнейшая судьба таких вирусов зависит от естественного отбора, сохраняющего популяцию, наиболее приспособленную к условиям существования.
Мутации могут иметь разные последствия. В одних случаях они ведут к изменению фенотипических проявлений в нормальных условиях. Например, увеличивается или уменьшается размер бляшек под агаровым покрытием; увеличивается или ослабляется нейровирулентность для определенного вида животных; вирус становится более чувствительным к действию химиотерапевтического агента и т. п.
В других случаях мутация является летальной, так как вследствие ее нарушается синтез или функция жизненно важного вирусспецифического белка, например вирусной полимеразы.
В некоторых случаях мутации являются условно летальными, так как вирусспецифический белок сохраняет свои функции в определенных, оптимальных для него, условиях и теряет эту способность в неразрешающих (непермиссивных) условиях. Типичным примером таких мутаций являются температурно-чувствительные (temperature sensitive) — ts-мутации, при которых вирус теряет способность размножения при повышенных температурах (39—42° С), сохраняя эту способность при обычных температурах выращивания (36—37° С).
По своему механизму мутации могут быть тоже разными. В одних случаях происходит делеция, т. е. выпадение одного или нескольких нуклеотидов, в других случаях происходит встраивание одного или нескольких нуклеотидов, а в некоторых случаях — замена одного нуклеотида другим.
Мутации могут быть прямыми и обратными. Прямые мутации меняют фенотип, а обратные мутации — реверсии — его восстанавливают. Возможны истинные реверсии, когда обратная мутация происходит в месте первичного повреждения, и псевдореверсии, если мутация происходит в другом участке дефектного гена (интрагенная супрессия) или в другом гене (экстрагенная супрессия). Реверсия не является редким событием, так как ревертанты обычно более приспособлены к данной клеточной системе. Поэтому при получении мутантов с заданными свойствами, например вакцинных штаммов, приходится считаться с возможной их реверсией к дикому типу.
Мутации носят случайный характер и объясняются статистическими законами.
В качестве физических мутагенов наиболее часто применяется ультрафиолетовое облучение, так как его энергия сопоставима с энергией химических связей. Реже применяются более жесткие виды облучения — рентгеновское и γ-облучение, а также обработка вирусных суспензий нейтронами, протонами, электронами и ядрами гелия, так как они вызывают сильные разрушения вирусных геномов и их инактивацию.
В качестве химических мутагенов применяют аналоги оснований (бромурацил, бромдезоксиуридин, 2-аминопурин, нитрозогуанидин и пр.), алкилирующие и флуоресцирующие соединения (профлавин), интеркалирующие агенты (актиномицин, этидий бромид), азотистую кислоту, гидроксиламин и многие другие.
ГЕНЕТИЧЕСКИЕ И НЕГЕНЕТИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ МЕЖДУ ВИРУСАМИ
Как в естественных, так и в экспериментальных условиях одна клетка может быть заражена не одним, а несколькими вирусами. В процессе такой смешанной инфекции могут иметь место различные формы взаимодействия как между вирусными геномами, так и между продуктами генов. При взаимодействии геномов могут наблюдаться такие формы генетических взаимодействий, как множественная реактивация, рекомбинация, пересортировка генов, кросс-реактивация, гетерозиготность. При взаимодействии на уровне продуктов генов могут иметь место негенетические взаимодействия: комплементация, интерференция, фенотипическое смешивание и др.
Множественная реактивация. Вирусная инфекция может возникнуть при заражении клетки несколькими ви-рионами с поврежденными геномами вследствие того, что функцию поврежденного гена может выполнять вирус, у которого этот ген не поврежден. Этот феномен был вначале обнаружен на бактериофагах и получил название множественной реактивации. В основе множественной реактивации лежит кооперативный процесс, при котором вирионы с поражением разных генов дополняют друг друга путем генетической рекомбинации, в результате чего репродуцируется исходный неповрежденный вирус. Эффективность множественности реактивации зависит от многих причин: степени повреждения генома вирионов, числа проникших в клетку вирионов, концентраций их в определенных участках клетки, аутоинтерференции поврежденных вирионов. Для множественной реактивации важное значение имеет расстояние между вирионами с поврежденными геномами внутри клетки. Обработка вирионов двухвалентными ионами металлов, ведущая к их агрегации, усиливает множественную реактивацию.
Рекомбинация. Генетической рекомбинацией называют обмен генетическим материалом, происходящий между родительскими вирусами. Возможен обмен полными генами (межгенная рекомбинация), так и участками одного и того же гена (внутригенная рекомбинация). Образующийся вирус-рекомбинант обладает свойствами, унаследованными от разных родителей.
Обычно рекомбинируемые штаммы обладают характерными признаками, которые обозначаются как маркеры. Например, были получены рекомбинанты между вирусами полиомиелита, обладающие повышенной устойчивостью и повышенной чувствительностью к гуанидину, разной нейровирулентностью, разной устойчивостью к повышенной температуре, разной чувствительностью к ингибиторам сывороток лошадей и коров и т. п. Для получения рекомбинантов используют штаммы, содержащие два или большее число маркеров.
Тест рекомбинации применяют для генетических исследований вирусов. С его помощью возможно построение генетических карт вирусов, в которых определяется, в каких участках генома произошли мутации, а также в условных единицах измеряется расстояние между разными мутациями.
Пересортировка генов. Вариантом рекомбинации является феномен, получивший название пересортировки генов. Она наблюдается при генетических взаимодействиях между вирусами, имеющими сегментированный геном. Образующиеся при этом гибридные формы вирусов называют реассортантами. Реассортанты вирусов гриппа получают при совместном культивировании вирусов с разными генами гемагглютинина и нейраминидазы. В этом случае из общего потомства путем нейтрализации соответствующих антигенов можно выделить интересующие исследователя варианты.
Существуют определенные группировки (констелляции или созвездия) генов, которые в данной системе клеток более стойки и делают вирус более жизнеспособным.
Сходные процессы пересортировки генов имеют место у вирусов гриппа типов А, В и С и у других вирусов с фрагментарным геном — у буньявирусов, аренавирусов (однонитчатые РНК) и реовирусов (ротавирусов) (двунитчатая РНК). Однако эти процессы не столь интенсивны и доступны изучению, как у вирусов гриппа.
Гетерозиготность. При совместном культивировании двух штаммов вируса может происходить формирование вирионов, содержащих в своем составе два разных генома или по крайней мере один полный геном и часть второго генома. Это явление названо гетерозиготностью.
Комплементация. Комплементация (дополнение) является таким видом негенетического взаимодействия при смешанной инфекции двумя вирусами, которое стимулирует репродукцию обоих партнеров или одного из них, но не изменяет генотипы вирусов. Принцип комплементации заключается в том, что вирус снабжает партнера недостающими компонентами, обычно белками, структурными или неструктурными.
Комплементация широко распространена среди вирусов и встречается как между родственными, так и неродственными вирусами. Феномен тесно связан с проблемой дефектности вирусов.
Поскольку в вирусной популяции помимо стандартных обычно присутствуют дефектные неинфекционные вирусные особи, в частности дефектные частицы, утратившие часть генетического материала, комплементация имеет место в инфекционном цикле многих вирусов и заключается в том, что члены популяции снабжают друг друга продуктами генов, которые дефектны у партнеров. Отличие комплементации от генетической рекомбинации заключается в отсутствии обмена генетическим материалом.
Комплементация встречается и у неродственных вирусов, принадлежащих к разным семействам. Одним из семейств, вирусы которого наиболее часто участвуют в комплементации, является семейство аденовирусов. В одних системах аденовирусы могут действовать как дефектные вирусы, в других — как помощники. Например, в культуре клеток почек макак резусов аденовирусы могут репродуцироваться только в присутствии SV40, который является в данном случае вирусом-помощником. В других системах сами аденовирусы действуют как вирусы-помощники, а вирусом-сателлитом является аденоассоци-ированный вирус, относящийся к семейству парвовирусов. Репродукция этого вируса полностью зависит от комплементирующего действия аденовирусов. Вирус гепатита В является помощником для дельта-агента, который покрывается его наружным белком — HBs-антигеном. Сочетание обоих вирусов обнаружено при наиболее тяжелых формах гепатита.
Возможна не только межцистронная, но и внутрицистронная комплементация в том случае, когда один ген кодирует несколько белков.
Фенотипическое смешивание. При совместном культивировании двух вирусов может наблюдаться феномен фенотипического смешивания, когда геном одного вируса бывает заключен в капсид, состоящий частично или полностью из белков другого вируса.
Фенотипическое смешивание наблюдается при смешанной инфекции,многими вирусами, причем эти вирусы могут быть как близкими друг другу (например, вирусы гриппа А и В или разные серологические подтипы вируса гриппа А), так и весьма далекими (онковирусы и рабдовирусы).
РЕСТРИКТАЗЫ И ФИЗИЧЕСКИЕ КАРТЫ ВИРУСОВ
Подлинную революцию в физическом картировании геномов вирусов произвело применение рестриктаз и секвенирование вирусных геномов. Рестриктазы имеют исключительное значение в молекулярной генетике вообще и генетической инженерии в частности. Их открытие (1968—1970 гг.) впервые дало возможность специфически расщеплять ДНК на строго определенные фрагменты, доступные для препаративного выделения и анализа.
Рестриктазы или эндодезоксирибонуклеазы — это просто организованные белки, являющиеся ферментами, широко распространенными среди прокариотов и участвующими в генетических процессах. В отличие от экзонук-леаз, отщепляющих концевые нуклеотиды или свободные остатки фосфорной кислоты, эндонуклеазы расщепляют молекулу ДНК изнутри, обычно — в местах, где преобладают пиримидиновые основания. Рестриктазы характеризуются высоковыраженной специфичностью, распознавая строго определенные последовательности нуклеотидов в двунитчатой ДНК.
Число новых рестриктаз стремительно нарастает и со временем, по-видимому, будут обнаружены рестриктазы, узнающие любую последовательность нуклеотидов.
Использование разных рестриктаз позволяет получать фрагменты разной величины, которые затем разделяются и анализируются путем электрофореза в агарозных или полиакриламидных гелях. Сочетание рестрикционного анализа с другими методами позволяет составить физические карты геномов вирусов. Физические карты вирусных геномов обозначают взаимное расположение генов, их границы, локализацию начала репликации, промоторов, лидеров, экзонов и интронов, сигнальных последовательностей и других генетических элементов.
Генетический код для синтеза белков вируса SV-40 записан не на одной, а на обеих нитях ДНК, а транскрипция разных генов идет в разных направлениях.
В настоящее время полностью расшифрованы нуклеотидные последовательности отдельных генов и целых геномов методом секвенирования (от англ. sequence — последовательность). Если речь идет о РНК-содержащих вирусах, то предварительным условием для дальнейшего их анализа является переписка РНК на ДНК с помощью РНК-зависимой ДНК-полимеразы (обратной транскриптазы), после чего генетический материал может быть подвергнут рестрикционному анализу.
Проектное задание к модулю
В качестве проектного задания студентам предлагается написание рефератов по следующим темам:
1. Природа дефектных вирусных геномов. Вирусы-сателлиты.
2. Вирусная интерференция
3. Необычные свойства ретровирусов
4. Трансформация клетки опухолеродными ДНК-вирусами
5. Классификация и основные свойства вирусов гриппа
6. Индукция специфического иммунного ответа на вирусы
7. Особенности репродукции пикорнавирусов
8. Семейство тогавирусов. Особенность репродукции и инфекционного процесса.
9. Вирус клещевого энцефалита.
10. Вирус бешенства
11. Вирусная персистенция
12. Основные свойства парамиксовирусов
13. Вирус кори. Биология возбудителя. Особенности патогенеза
14. Аденовирусы, их репликация и связь с другими вирусами
16. Пути распространения вирусных болезней растений.
17. Прионы. Возбудители или провокаторы или….
18. Вироиды как вирусоподобные инфекционные агенты.
19. Вирусы, вызывающие респираторные инфекции. Сравнительная характеристика
20. Место вирусов в биосфере.
21. Бактериофаги. Их морфологическое многообразие и взаимодействие с бактериями.
22. Атипичная пневмония и возможность происхождения новых вирусов
23. Роль вирусов в возникновении злокачественных опухолей
24. Особенности транскрипции РНК- содержащих вирусов.
25. Морфогенез вирусов или морфологические превращения в процессе репродукции.
Тест рубежного контроля
Стратегия репликации вирусов. Методика репликации вирусов.
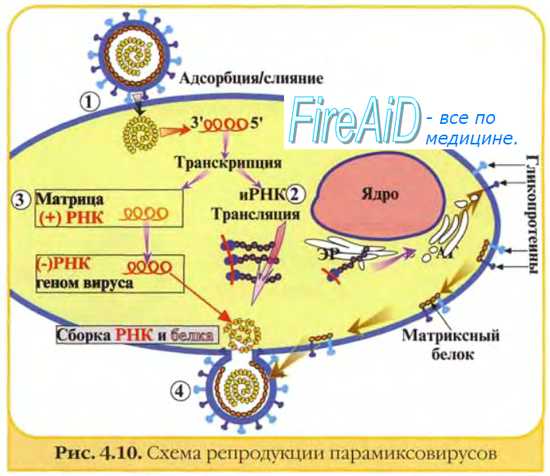
В ходе эволюции вирусов сложилось несколько стратегий, обеспечивающих: а) организацию вирусных геномов и их кодирующую функцию; б) экспрессию вирусных генов; в) репликацию вирусных геномов и г) сборку и созревание вирусного потомства. Прежде чем рассмотреть данные положения, следует напомнить, что фундаментальные биологические особенности вирусов делают реализацию их генетической информации процессом, не имеющим себе аналогов среди других форм жизни, включая микоплазмы, риккетсии и хламидии. Во-первых, это обусловлено тем, что вирусы содержат один тип нуклеиновой кислоты — РНК или ДНК. Во-вторых, у вирусов отсутствует собственный аппарат для реализации своей генетической информации. Вирусы не имеют ферментов, участвующих в производстве энергии, и белоксинтезирующих систем. Поэтому вирус должен представить информационную РНК, которую клетка должна распознавать и транслировать. Пути образования мРНК в процессе репликации РНК-вирусов представлены на рисунке. В клетке нет ферментов для транскрипции мРНК с вирусного РНК-генома, а в цитоплазме нет ферментов для транскрипции вирусной ДНК.
В связи с этим клеточную транскриптазу для синтеза вирусных мРНК могут использовать только вирусы, содержащие ДНК и способные проникать в ядро. Все другие вирусы вынуждены создавать собственные ферменты для синтеза мРНК.
Организация вирусного генома
Почти все РНК-геномы относят к линейным молекулам. Некоторые из них, например, геномы пикорнавирусов, содержат на 5'-конце РНК ковалентно связанные полипептиды. Геномы одноцепочечных РНК-вирусов могут быть либо монолитными (пикорна-, тога-, парамиксо-, рабдо-, корона-, ретровирусы), либо сегментированными (ортомиксо-, арена- и буньявирусы). Среди РНК вирусов с двухцепочечным сегментированным геномом наиболее изучены реовирусы.
Все известные ДНК-вирусы позвоночных имеют монолитный геном. Все они, за исключением парвовирусов, имеют полностью или частично двухспи-ральный геном. Отдельные представители парвовирусов содержат линейную оцДНК; у других (например, у аденоассоциированных вирусов) комплементарные цепи ДНК находятся в разных вирионах. ДНК папиллома- и полиомавиру-сов — кольцевая и сверхспиральная, в то время как ДНК герпесвирусов, аденовирусов и осповирусов — линейная.
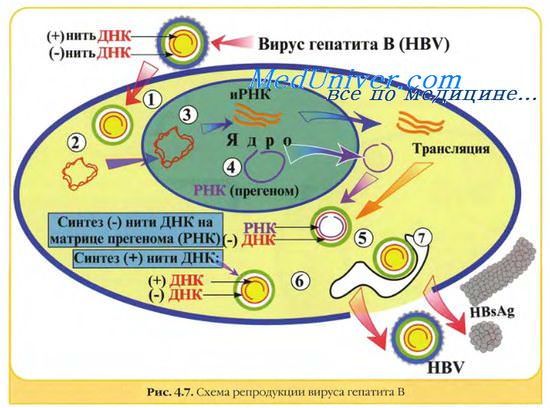
ДНК вируса гепатита В представляет собой кольцевую двуспиральную молекулу, на обеих цепях которой в разных областях обнаружены большой и малый одноцепочечные участки.
Репликация большинства ДНК-вирусов включает механизмы, которые хорошо известны в клеточной биологии: транскрипция мРНК на двуцепочечной ДНК и репликация ДНК. Для РНК-вирусов ситуация является уникальной и связана с генетической информацией, кодируемой РНК. РНК-вирусы с различным типом генома связаны с необходимостью использовать различные пути синтеза мРНК. Кроме (+) оцРНК вирусов, у которых геномная РНК функционирует как мРНК, все другие типы вирусной РНК, прежде всего, должны быть транскрибированы в мРНК. Так как клетки эукариотов не содержат РНК-зависимой РНК-полимеразы, (-) оцРНК вирусы и дцРНК вирусы должны содержать вирионную РНК-зависимую РНК-полимеразу
Эукариотические клетки не могут транслировать полицистронные мРНК частями в виде нескольких индивидуальных белков. ДНК-вирусы расщепляют свои полицисторные РНК-транскрипты на моноцисторные мРНК, используя клеточный механизм.
Большинство РНК-вирусов, размножающихся в цитоплазме, не имеют доступа к ферментам, осуществляющим РНК-процессинг и сплайсинг в ядре, и вынуждены по-разному решать эту проблему. У вирусов с сегментированным геномом каждый фрагмент является отдельным геном. Другие вирусы имеют полицистронный геном, но образуют моноцистронные РНК-транскрипты, разделяя окончание и реинициацию транскрипции.
Некоторые вирусы имеют полицистронную РНК, которая транслируется в полипротеин, который затем расщепляется протеолитически с образованием конечных продуктов.
Экспрессия и репликация вирусного генома
Анализ данных о молекулярных механизмах репликации вирусов позвоночных позволил установить сходство и различия в способах реализации генетической информации вирусов, относящихся к различным таксономическим группам. Благодаря этому оказалось возможным выделить основные типы стратегии экспресии и репликации вирусных геномов. Семейства вирусов, реплицирующиеся по сходному пути, объединены в один тип. Репликация вирусов каждого типа начинается с инициирующих единиц, которыми являются проникшие в клетку вирусные геномы: вирионные РНК или ДНК или вирионные нуклеопротеины.
Вирусы, геном которых может служить в качестве мРНК, принято называть вирусами с позитивным (+) геномом. Вирусы с негативным (-) РНК геномом неспособность функционировать в качестве мРНК компенсируют наличием вирионной транскриптазы.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Типы инфицирования клеток вирусами. Репродуктивный цикл вирусов. Основные этапы репродукции вирусов. Адсорбция вириона к клетке.
По характеру взаимодействия генома вируса с геномом клетки выделяют автономное (геном вируса не интегрирован в геном клетки) и интеграционное (геном вируса интегрирован в геном клетки) инфицирование. Особую форму составляют латентное и персистирующее инфицирование.
Персистирующее инфицирование клеток вирусам. Некоторые РНК-вирусы могут вызывать персистиру-ющие инфекции, проявляющиеся образованием дочерних популяций возбудителя после завершения острой фазы болезни. При этом происходит постепенное выделение вирусных частиц, но инфицированная клетка не лизируется. Нередко дочерние популяции вирионов дефектны (часто наблюдают у лиц с иммунодефицитами). Иногда такие хронические поражения протекают без клинических проявлений. В частности, вирус гепатита В способен вызывать персистирующее поражение гепатоцитов с развитием хронического гепатита; в дальнейшем возможна малигнизация клеток.
Репродуктивный цикл вирусов
Изображённые на рис. 2-3 этапы репродукции (от адсорбции вирионов до высвобождения дочерней популяции) происходят при продуктивном взаимодействии вируса с клеткой.
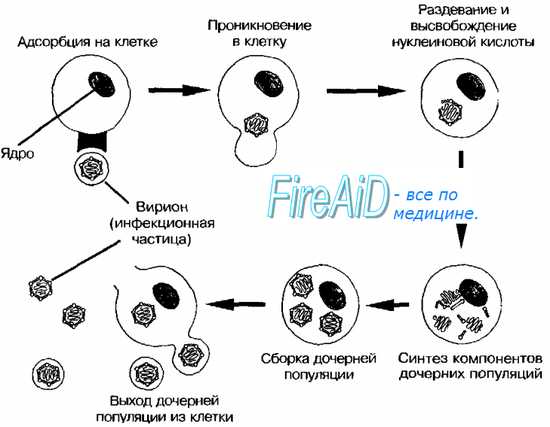
Рис. 2-3. Основные этапы репродукции вирусов.
Адсорбция вириона к клетке
• Процесс адсорбции не зависит от температуры (то есть не требует энергетических затрат) и протекает в две фазы; фаза ионного притяжения обусловлена неспецифическим взаимодействием, фаза прикрепления происходит благодаря структурной гомологии либо комплемен-тарности взаимодействующих молекул.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Транскрипция вируса в клетке. Трансляция вирусов.
Транскрипция +РНК-содержащих вирусов. Функции мРНК выполняет геном (+РНК), поэтому у таких вирусов для синтеза вирусных белков (трансляция) нет необходимости в процессе транскрипции. Другими словами, у +РНК-содержащих вирусов транскрипция отсутствует.
Транскрипция -РНК-содержащих вирусов и вирусов, имеющие две нити РНК. Функции мРНК выполняют транскрипты, комплементарные -РНК вириона. Поэтому у таких вирусов транскрипция существует как самостоятельный этап репродуктивного цикла. Для образования транскриптов в составе вирионов имеется собственная РНК-полимераза (транскриптаза).
Транскрипция ДНК-вирусов. Транскрипция — самостоятельный этап репродуктивного цикла, так как геном ДНК-вирусов должен транскрибироваться для образования мРНК. Вирусы, репродуцирующиеся в ядре (например, герпес- и аденовирусы) для этой цели используют клеточную ДНК-зависимую РНК-полимеразу (транскриптазу). Вирусы, репродуцирующиеся в цитоплазме (например, поксвирусы) лишены такой способности и содержат (как и вирусы с -РНК) собственную транскриптазу.
Рис. 2-3. Основные этапы репродукции вирусов.
Трансляция вирусов
Вирусные геномы кодируют синтезы двух классов белков: структурные белки входят в состав дочерних популяций, а неструктурные белки обслуживают процессы репродукции, но не входят в состав дочерних популяций (ингибиторы синтеза клеточных РНК и белков, протеазы и др.).
Трансляция РНК-содержащих вирусов. Поскольку вирусный геном кодирует несколько белков, то возможно два варианта трансляции:
1) каждый полипептид синтезируется отдельно от других (тога- и ретровирусы);
Некоторые вирусы используют оба этих механизма. Полипептиды, образующиеся при обоих вариантах трансляции, могут подвергаться посттрансляционной модификации (гликозилирование, фосфорилирование или сульфатирование).
Трансляция ДНК-содержащих вирусов. В трансляционных процессах доминирует трансляция отдельных мРНК, кодирующих индивидуальные полипептиды. В отдельных случаях (например, у аденовирусов) не менее трёх белков образуются путём нарезания общего полипептида-предшественника.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Транскрипция — это переписывание ДНК на РНК по законам генетического кода. Это означает, что РНК состоит из нуклеотидных последовательностей, комплементарных ДНК. Нити ДНК в участке транскрипции разделяются и функционируют как матрицы, к которым присоединяются комплементарные нуклеотиды благодаря спариванию комплементарных оснований (аденин связывается с тимином, урацил — с аденином, гуанин — с цитозином и цитозин - с гуанином). Транскрипция осуществляется с помощью специального фермента — РНК-полимеразы, который связывает нуклеотиды путем образования 3'-5'-фосфодиэфирных мостиков. Такое связывание происходит лишь в присутствии ДНК-матрицы.
Продуктами транскрипции в клетке являются иРНК. Сама клеточная ДНК, являющаяся носителем генетической информации, не может непосредственно программировать синтез белка. Передачу генетической информации от ДНК к рибосомам осуществляет РНК-посредник. На этом основана центральная догма молекулярной биологии, которая выражается следующей схемой:
ДНК - (транскрипция) –и РНК – (трансляция) - белок
где стрелки показывают направление переноса генетической информации.
Реализация генетической информации у вирусов. Стратегия вирусного генома в отношении синтеза иРНК у разных вирусов различна. У ДНК-содержащих вирусов иРНК синтезируется на матрице одной из нитей ДНК. Формула переноса генетической информации у них такая же, как и в клетке.
ДНК-содержащие вирусы, репродукция которых происходит в ядре, используют для транскрипции клеточную полимеразу. К этим вирусам относятся паповавирусы, аденовирусы, вирусы герпеса. ДНК-содержащие вирусы, репродукция которых происходит в цитоплазме, не могут использовать клеточный фермент, находящийся в ядре. Транскрипция их генома осуществляется вирусспецифическим ферментом — ДНК-полимеразой, которая проникает в клетку в составе вируса. К этим вирусам относятся вирусы оспы и иридовирусы.
К этой группе вирусов относятся пикорнавирусы, тогавирусы, коронавирусы. У них нет необходимости в акте транскрипции для синтеза вируспецифических белков. Поэтому транскрипцию как самостоятельный процесс у этих вирусов не выделяют. Иначе обстоит дело у вирусов, геном которых не может выполнять функцию иРНК. В клетке синтезируется комплементарная геному РНК, которая и является информационной. Передача генетической информации у этих вирусов осуществляется по схеме:
РНК– иPHK - белок
У этих вирусов транскрипция выделена как самостоятельный процесс в инфекционном цикле. К ним относятся две группы вирусов животных.
2. Вирусы, геном которых представлен двунитчатой РНК (диплорнавирусы). Среди вирусов животных к ним относятся реовирусы.
В клетке нет фермента, который может полимеризовать нуклеотиды на матрице РНК. Эту функцию выполняет вирусспецифический фермент — РНК-полимераза, или транскриптаза, которая находится в составе вирусов и вместе с ними проникает в клетку.
Среди РНК-содержащих вирусов животных есть семейство ретровирусов, которые имеют уникальный путь передачи генетической информации. РНК этих вирусов переписывается на ДНК, ДНК интегрирует с клеточным геномом и в его составе переписывается на РНК, которая обладает информационными функциями. Путь передачи генетической информации в-этом случае осуществляется по более сложной схеме:
РНК - ДНК - иРНК - белок
Ферменты, транскрибирующие вирусный геном. Транскрипция ряда ДНК-содержащих вирусов — паповавирусов, аденовирусов, вирусов герпеса, парвовирусов, гепадна-вирусов осуществляется в ядре клетки, и в этом процессе широко используются механизмы клеточной транскрипции — ферменты транскрипции и дальнейшей модификации транскриптов. Транскрипция этих вирусов осуществляется клеточной РНК-полимеразой II — ферментом, который осуществляет транскрипцию клеточного генома. Однако особая группа транскриптов аденовируса синтезируется с помощью другого клеточного фермента — РНК-полимеразы III. У двух других семейств ДНК-содержащих вирусов животных - вирусов оспы и иридовирусов — транскрипция происходит в цитоплазме. Поскольку в цитоплазме нет клеточных полимераз, транскрипция этих вирусов нуждается в специальном вирусном ферменте — вирусной РНК-полимеразе. Этот фермент является структурным вирусным белком.
У РНК-содержащих вирусов транскрипция осуществляется вирусспецифическими транскриптазами, т. е. ферментами, закодированными в вирусном геноме. Вирусспецифические транскриптазы могут быть как структурными белками, входящими в состав вириона (эндогенная транскриптаза), так и неструктурными белками, которые синтезируются в зараженной клетке, но не включаются в вирион.
Транскрипция в зараженной клетке. Синтез комплементарных РНК на родительских матрицах с помощью родительской транскриптазы носит название первичной транскрипции в отличие от вторичной транскрипции, происходящей на более поздних стадиях инфекционного цикла на вновь синтезированных, дочерних матрицах, с помощью вновь синтезированной транскриптазы. Большая часть иРНК в зараженной клетке является продуктом вторичной транскрипции.
Транскриптивные комплексы. У сложно устроенных РНК-содержащих вирусов животных транскрипция происходит не на матрице голой РНК, а в составе вирусных нуклеокапсидов или сердцевин (транскриптивные комплексы). Связанные с геномом капсидные белки не только не препятствуют транскрипции, но и необходимы для нее, обеспечивая правильную конформацию тяжа РНК, защиту его от клеточных протеаз, связь отдельных фрагментов генома друг с другом, а также регуляцию транскрипции.
Регуляция транскрипции. Транскрипция вирусного генома строго регулируется на протяжении инфекционного цикла. Регуляция осуществляется как клеточными, так и вирусспецифическими механизмами. У некоторых вирусов, в основном ДНК-содержащих, существует три периода транскрипций — сверхранняя, ранняя и поздняя. К этим вирусам относятся вирусы оспы, герпеса, паповавирусы, аденовирусы. В результате сверхранней и ранней транскрипции избирательно считываются сверхранние и ранние гены с образованием сверхранних или ранних иРНК. При поздней транскрипции считывается другая часть вирусного генома — поздние гены, с образованием поздних иРНК. Количество поздних генов обычно превышает количество ранних генов. Многие сверхранние гены являются генами для неструктурных белков — ферментов и регуляторов транскрипции и репликации вирусного генома. Напротив, поздние гены обычно являются генами для структурных белков. Обычно при поздней транскрипции считывается весь геном, но с преобладанием транскрипции поздних генов.
Фактором регуляции транскрипции у ядерных вирусов является транспорт транскриптов из ядра в цитоплазму, к месту функционирования иРНК — полисомам.
У РНК-содержащих вирусов синтез транскриптов также строго контролируется в отношении как количества каждого класса транскриптов, так и периода инфекции, когда определенные транскрипты синтезируются с максимальной скоростью. На ранней стадии инфекции преимущественно синтезируются транскрипты двух генов вируса гриппа — NP и NS, на поздней стадии инфекции — транскрипты генов М, НА и NA. Остальные три гена для Р-белков синтезируются примерно с одинаковой скоростью на протяжении всего периода инфекции. У реовирусов на ранней стадии инфекции преимущественно транскрибируется 4 из 10 фрагментов генома и лишь на поздней стадии транскрибируется весь геном. Однако если поместить геном вируса в бесклеточную РНК-синтезирующую систему, будет происходить равномерная транскрипция всех 10 фрагментов генома. Эти факты говорят о жестком контроле транскрипции со стороны клетки-хозяина и возможном наличии специфических клеточных регуляторов.
ТРАНСЛЯЦИЯ
Синтез белка в клетке происходит в результате трансляции иРНК. Трансляцией называется процесс перевода генетической информации, содержащейся в иРНК, на специфическую последовательность аминокислот. Иными словами, в процессе трансляции осуществляется перевод 4-буквенного языка азотистых оснований на 20-буквенный язык аминокислот.
Рибосомы. Синтез белка в клетке осуществляется на рибосоме. Рибосома состоит из двух субъединиц, большой и малой, малая субъединица примерно в два раза меньше большой. Обе субъединицы содержат по одной молекуле рибосомальной РНК и ряд белков. Рибосомальные РНК синтезируются в ядре на матрице ДНК с помощью РНК-полимеразы. В малой рибосомальной субъединице есть канал, в котором находится информационная РНК. В большой рибосомальной субъединице есть две полости, захватывающие также малую рибосомальную субъединицу. Одна из них содержит аминоацильный центр (А-центр), другая — пептидильный центр (П-центр).
Фазы трансляции. Процесс трансляции состоит из трех фаз: 1) инициации, 2) элонгации и 3) терминации.
Вначале с иРНК связывается малая рибосомальная субъединица. К комплексу иРНК с малой рибосомальной субъединицей присоединяются другие компоненты, необходимые для начала трансляции. Их по крайней мере три в прокариотической клетке и более девяти в эукариотической клетке. Инициаторные факторы определяют узнавание рибосомой специфических иРНК и, таким образом, являются определяющим фактором в дискриминации между различными иРНК, присутствующими в клетке, как правило, в избыточном количестве.
В результате формируется комплекс, необходимый для инициации трансляции, который называется инициаторным комплексом. В инициаторный комплекс входят: 1) иРНК; 2) малая рибосомальная субъединица; 3) аминоацил-тРНК, несущая инициаторную аминокислоту; 4) инициаторные факторы; 5) несколько молекул ГТФ.
В рибосоме осуществляется слияние потока информации с потоком аминокислот. Аминоацил-тРНК входит в А-центр большой рибосомальной субъединицы, и ее антикодон взаимодействует с кодоном иРНК, находящейся в малой рибосомальной субъединице. При продвижении иРНК на один кодон тРНК перебрасывается в пептидильный центр, и ее аминокислота присоединяется к инициаторной аминокислоте с образованием первой пептидной связи. Свободная от аминокислоты тРНК выходит из рибосомы и может опять функционировать в транспорте специфических аминокислот.
Терминация трансляции. Терминация трансляции происходит в тот момент, когда рибосома доходит до терминирующего кодона в составе иРНК. Трансляция прекращается, и полипептидная цепь освобождается из полирибосомы. После окончания трансляции полирибосомы распадаются на субьединицы, которые могут войти в состав новых полирибосом.
Читайте также: