
Тип симметрии вируса гриппа
Обновлено: 18.04.2024
Внутри вириона гриппа А находятся восемь сегментов вирусной РНК. Эти молекулы несут всю необходимую информацию для образования новых частиц вируса гриппа. Эти восемь РНК схематично показаны желто-зелеными линиями в верхней части рис. 2. РНК представляют собой цепи из четырех различных нуклеотидов – A, C, G и U. В случае с вирусом гриппа восемь РНК имеют длину порядка 14000 нуклеотидов. Нуклеотиды составляют генетический код, который считывается преобразующим механизмом клетки в триплеты, определяющие аминокислоту.
Необходимо рассмотреть два важных аспекта этих вирусных РНК. Во-первых, вы можете видеть, что концы вирусных РНК помечены как 3′ и 5′. Нуклеиновые кислоты обладают полярностью, так что один конец цепи имеет отличные химические свойства от другого. Эта полярность представлена как 3′ и 5′. Во-вторых, во время копирования, или удвоения, нуклеиновой кислоты ферментами, которые носят название полимераз, появляется нить с комплементарной полярностью. Вирусные РНК гриппа называются (-), или РНК с отрицательной нитью, поскольку они представляют собой отрицательную полярность РНК с трансляцией в белок. Молекулы РНК, являющиеся образцами для синтеза белков, называются (+), или положительной полярностью. После вхождения в клетку отрицательная нить (-) вирусной РНК гриппа должна скопироваться в комплементарные (+) нити, так что они могут служить образцом для белков. Вирусные РНК копируются ферментом (РНК-полимеразой), который привносится в клетку вместе с вирусом.
На рис. 2 желто-зелеными линиями обозначены обнаруженные в вирионе гриппа РНК с отрицательными нитями. Как только вирион входит в клетку, эти восемь РНК копируются в положительную нить мРНК. Наконец, мРНК может служить образцом для синтеза белков. Специфические вирусные белки, производимые каждой вирусной мРНК, показаны в нижней части рис. 2. Видно, что, например, сегмент 4 РНК несет информацию о вирусном белке HA, а сегмент 6 – о вирусном белке NA. Заметьте, что некоторые сегменты РНК несут информацию более чем об одном белке. Вирусы гриппа субтипов А и B имеют восемь сегментов РНК, тогда как субтипа C – только семь.
Вирусы гриппа называются РНК-вирусами с отрицательной нитью из-за полярности РНК, привносимой в вирион. Другие РНК-вирусы, такие как полиовирус, являются РНК-вирусами c положительной нитью, так как их геномная РНК может преобразовываться в белок сразу после вхождения в клетку.
1.3. Субтипы A, B и C вируса гриппа
рисунок). Оболочечные вирионы с шестиугольной структурой на поверхности формируют длинную (500 микрон) напоминающую шнур структуру при отпочковании от клетки (рис. 4). Как и в случае с вирусами гриппа A и B, ядро вируса гриппа C состоит из рибонуклеопротеина, созданного из вирусной РНК и 4 белков. Белок M1 расположен под мембраной, как и в вирионах гриппа A и B. Второстепенный вирусный оболочечный белок CM2 функционирует как ионный канал. Основной оболочечный гликопротеин вируса гриппа C называется HEF (слияние гемагглютинина и эстеразы – hemagglutinin-esterase-fusion), поскольку он обладает функциями и HA, и NA. Поэтому вирион гриппа содержит 7 сегментов РНК, а не 8, как вирусы гриппа субтипов A и B.
Практически все взрослые когда-либо заражались вирусом гриппа C, вызывающим мягкое течение болезни в верхних дыхательных путях. Осложнения с переходом на нижние дыхательные пути – редкость. Против вируса гриппа C не существует вакцины.
Мне достаточно хорошо знакомы вирусы гриппа B и C – я получил степень доктора наук за их изучение. В моей работе говорится, что геном вируса гриппа C состоит из 7 сегментов РНК, и показана рекомбинация среди различных штаммов вируса гриппа C.
Рисунок 4.
Рекомендуемая литература.
- Hatta, M., & Kawaoka, Y. (2003). The NB Protein of Influenza B Virus Is Not Necessary for Virus Replication In Vitro Journal of Virology, 77 (10), 6050-6054 DOI:10.1128/JVI.77.10.6050-6054.2003.
- RacanielloVR, & Palese P (1979). Isolation of influenza C virus recombinants. Journal of Virology, 32 (3), 1006-14 PMID: 513198.
1.4. РНК вируса гриппа: трансляция в белок
Рисунок 6.
Рисунок 7.
Последовательности, взятые нами для образца, принадлежат штамму 1918 H1N1 вируса гриппа. Обратите внимание на аминокислоту PB1-F2, выделенную синим цветом. Эта аминокислота играет важную роль в биологическом функционировании белка, которую мы еще рассмотрим впоследствии.
Эпидемиология гриппа. Распространенность гриппа. Морфология вирусов гриппа.
Грипп [от фр. gripper, хватать], или инфлюэнца [от итал. influenza di freddo, влияние холода], — острая инфекция, проявляющаяся поражениями дыхательного тракта, непродолжительной лихорадкой, упадком сил, головной болью, миалгиями и др.
Резервуар гриппа — инфицированный человек (больные и бессимптомные носители). Больной становится заразным за 24 ч до появления основных симптомов и представляет эпидемическую опасность в течение 48 ч после их исчезновения.
Грипп регистрируют повсеместно, рост заболеваемости наблюдают в холодные месяцы. Эпидемии гриппа развиваются с интервалом 2-3 года.
Передача возбудителя гриппа происходит воздушно-капельным путём. Наиболее восприимчивы дети и лица преклонного возраста. Вирусы гриппа чувствительны к действию высоких температур, высушиванию, инсоляции и УФ-облучению. Также они лабильны к действию эфира, фенола, формальдегида и других веществ, денатурирующих белки.

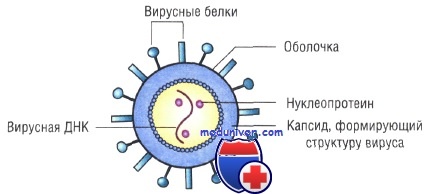
Морфология вирусов гриппа
Суперкапсид вируса гриппа образован липидным бислоем, который пронизывают гликопротеиновые шипы (спикулы), определяющие гемагглютинирующую (Н) либо нейраминидазную (N) активность. Репликация ортомиксовирусов первично реализуется в цитоплазме инфицированной клетки; синтез вирусной РНК происходит в ядре.
• Гемагглютинин обусловливает проникновение вирусов гриппа в клетки в результате слияния с мембраной клетки и мембранами лизосом. AT к нему обеспечивают защитный эффект. Нейраминидаза распознаёт и взаимодействует с рецепторами, содержащими N-ацетилнейраминовую кислоту, то есть приводит к проникновению вируса, а также, отщепляя нейраминовую кислоту от дочерних вирионов и клеточной мембраны, к выходу вирусов из клеток.
• Семь сегментов вирусного генома кодируют структурные белки, восьмой — неструктурные белки NS1 и NS2 вируса гриппа, существующие только в инфицированных клетках. Основные из них — матриксный (М) и нуклеопротеидный (NP) белки. В меньших количествах присутствуют внутренние белки (P1, P2, Р3), участвующие в этапах транскрипции и репликации вируса гриппа.
• М-белок вирусов гриппа играет важную роль в морфогенезе вирусов и защищает геном, окружая нуклеокапсид. Белок NP выполняет регуляторные и структурные функции. Внутренние белки являются ферментами: Р1 — транскриптаза, Р2 — эндонуклеаза, Р3 — репликаза.
Штаммы вируса гриппа. Антигены вируса гриппа. Строение вируса гриппа.
Вирулентные штаммы вируса гриппа, в отличие от невирулентных, обязательно активизируются за счет расщепления протеазами. Гемагглютинины вируса гриппа расщепляются внутриклеточно и потому способны инфицировать клетки различных тканей и вызывать системную инфекцию. Ортомиксовирусы с нерасщепленным НА имеют низкую инфекционную активность. Инфекционная форма вируса с расщепленным НА в организме млекопитающих образуется только в клетках, выстилающих дыхательный тракт. При отсутствии трипсина в культуре клеток расщепляется НА только вирулентных штаммов вируса. Для вируса гриппа птиц установлена строгая корреляция между расщепляемостью НА, способностью размножаться в культуре клеток и патогенностью для кур. Расщепление НА на субъединицы не является обязательным для сборки и выхода вирионов из клетки и проявления гемагглютинирующей активности. Полагают, что фактором, детерминирующим патогенность вируса гриппа птиц, является чувствительность НА к протеолитическому расщеплению.
В сайте расщепления НА у вирулентных штаммов вируса гриппа содержится несколько основных аминокислот, а у невирулентных — только один остаток аргинина. Таким образом, только участок из нескольких основных аминокислот у С-конца НА образует сайт узнавания для ферментов, ответственных за расщепление.

Основные функции НА: гемагглютинирующая активность вируса; прикрепление вирионов к клеткам-мишеням путем связывания с сиалосодержащими рецепторами; слияние вирусной и клеточной мембран. НА является основным специфическим антигеном вируса, определяющим (наряду с NA) подтип и вызывающим образование антител, нейтрализующих инфекционность вируса и его ГА-активность. НА играет главную роль в индукции протективного иммунитета при гриппе. В молекуле НА имеются 3-4 антигенных домена, изменения в которых определяют антигенный дрейф. NA является ферментом, катализирующим отщепление сиаловой кислоты от субстрата. Удаление сиаловой кислоты с НА облегчает его расщепление клеточными протеазами. Антитела к белку NA имеют вспомогательное значение в защите и нейтрализуют вирус лишь при высоком титре. Матриксный белок М - самый низкомолекулярный структурный белок. Он принимает участие в морфогенезе вириона и стабилизации его структуры. Нуклеопротеид (NP) - основной внутренний белок, формирующий субъединицы капсида. Белки NP и М являются типоспецифическими антигенами, общими для всех вирусов гриппа одного типа, и в этом отношении резко отличаются от высокодивергентных поверхностных белков. Они не вызывают образования протективных антител. Однако нуклеопротеин вируса гриппа — основной антиген, узнаваемый цитотоксическими Т-лимфоцитами. Аминокислотные последовательности 260—283 нуклеопротеина вируса гриппа А являются индукторами Т-клеточного ответа. Изменение антигенных свойств вируса гриппа — результат двух генетических процессов: антигенных дрейфа и шифта. Антигенный дрейф происходит в основном через накопление аминокислотных замен (точечных мутаций) в НА1. Замена одного аминокислотного остатка в эпитопе нарушает его связывание с соответствующими МАТ. Основные механизмы антигенного шифта — реассортация отдельных генов, возвращение в популяцию старых генов, прямые мутации, изменяющие специфичность к хозяину. NA может измениться независимо от НА. Вариабельность НА вируса гриппа А значительно выше вариабельности НА вируса гриппа В.
В процессе адаптации вируса гриппа к различным системам наблюдают изменение антигенной структуры НА, в основе которой лежит селекция мутантов с измененной рецепторсвязывающей специфичностью. Вирусы гриппа А и В человека, размноженные в КЭ, могут изменяться антигенно и претерпевать мутации в ГА (замена трех аминокислот в ГА). Вирус, размноженный в КЭ или в культуре клеток, защищает против культурального вируса более эффективно, чем против полевого вируса, выделенного от человека.
Один и тот же клинический изолят вируса гриппа при размножении в культуре клеток животных (МДСК) или в КЭ имеет разные антигенные свойства, что связано с заменой одной-двух аминокислот в разных участках молекулы НА. Особенно заметные изменения антигенности в молекуле НА наблюдали при замене аминокислотных остатков 187 и 189 в процессе адаптации вируса к КЭ. Изменение сайта гликозилирования может сопровождаться изменением антигенности и вирулентности вируса. Состав углеводов гемагглютинина вирусов и гриппа птиц может изменяться в различных хозяйских клетках даже в течение одного пассажа. У вирусов гриппа птиц типа А идентифицировано девять нейраминидазных N-антигенов, обозначенных 1-9, и 13 гемагглютинирующих антигенов Н, обозначенных 1-13. Состав Н- и N-антигенов полевых изолятов зависит от вида птиц, места и времени вспышек гриппа. Свиньи восприимчивы к различным подтипам вируса гриппа А и, возможно, вместе с водоплавающей птицей являются главным резервуаром вируса гриппа в природе.
В пермиссивных клетках вирусный НА активируется путем расщепления на две части НА1 и НА2, которые остаются связанными дисульфидными связями. Вирионы прикрепляются к клеткам, активированным НА, соединяются с рецепторами сиаловой кислоты плазматической мембраны и входят в клетку эндоцитозом. После сплавления оболочки вируса и эндоплазматической мембраны транскрипционный комплекс освобождается и транспортируется в ядро, где происходит транскрипция и репликация РНК.
Так же, как у всех других вирусов с негативно-полярным РНК-геномом, геном ортомиксовирусов выполняет две функции: 1) матричную для синтеза мРНК и 2) матричную для синтеза позитивно-полярной промежуточной формы РНК, которая служит матрицей для синтеза потомства геномной РНК. Из 8 первичных транскриптов, синтезированных на 8 генных сегментах вирусов гриппа А и В, 6 являются моноцисторными и транслируются прямо в белки. Два других — подвергаются сплайсингу с образованием двух мРНК, которые транслируются с различных рамок считывания с образованием двух белков. Вирусные белки синтезируются, используя клеточный механизм трансляции. Ортомиксовирусы используют несколько механизмов для увеличения кодирующей способности: сплайсинг мРНК, спаренную стоп-старт-трансляцию тандемных генов и сдвиг рамки считывания.
Репликация геномных РНК сегментов требует синтеза полноразмерных, положительной полярности РНК посредников, которые, в отличие от соответствующих мРНК транскриптов, не имеют КЭП-структуры на 5'-конце и поли (А) последовательности на З'-конце. Вновь синтезированный нуклеопротеин присоединяется к этим РНК, облегчая их использование в качестве матрицы для синтеза геномной РНК. Вирионы формируются почкованием, включая М белок и нуклеокапсид, которые встроены на плазматической мембране, в которую включены НА и NA. Механизмы копирования каждого РНК сегмента и включение их в каждый вирион неизвестны.
Хотя в результате реассортации генов теоретически могут возникать вирусы с любой комбинацией Н и N генов, однако только ограниченное число вирусов с определенной комбинацией Н и N генов оказались важными патогенами, имеющими право на существование. В том числе вызывающие респираторную патологию у людей: H1N1, H2N2, H3N2, H5N1 и, возможно, H3N8; у лошадей H7N7 и H3N8; у норок H10N4; у котиков H7N7 и H4N5; вирусы, часто выделяемые от свиней - H1N1 и H3N2. Главными возбудителями гриппа птиц являются вирусы с H5N2 и H7N1, хотя встречаются и другие варианты вируса гриппа А. Так как установлена резистентность диких видов птиц к вирусу гриппа А, им отводили роль резервуара вируса, прежде всего для домашней птицы. При экспериментальном заражении вирусом H1N5 дикие утки, как правило, не проявляли клинических признаков болезни, хотя вирус в их организме размножался и выделялся во внешнюю среду. Латентное инфицирование вирусом гриппа может иметь место у других домашних и диких животных.
Строение вирусов. Классификация вирусов
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
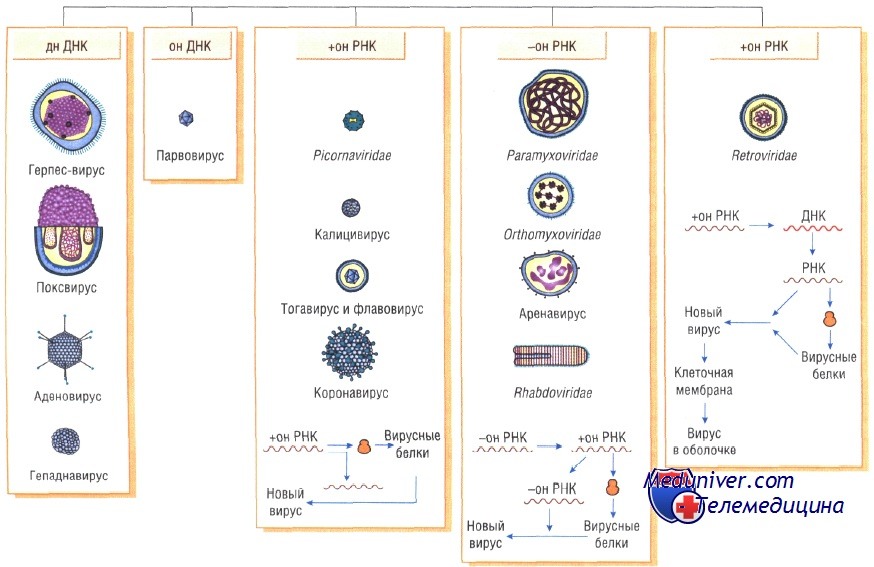
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Обзор
Автор
Редактор
Вирус гриппа

Рисунок 1. Вирус гриппа. Слева — электронная микрофотография вирионов гриппа, увеличение ≈100 000 раз. Справа — молекулярное строение вируса гриппа; из 11 вирусных белков, закодированных в геноме, состоящем из нескольких фрагментов РНК, подписаны только гликопротеины оболочки гемагглютинин ( HA ) и нейраминидаза ( NA ).
Впервые появившись в XVI веке, грипп вызывает регулярные эпидемии и пандемии, причём с серотипом А связывают эпидемии с промежутками 2–3 года (наиболее крупные эпидемии и пандемии сведены в таблицу), а с серотипом B — каждые 4–6 лет. Серотип С эпидемий не вызывает; исключение составляют единичные вспышки у детей и ослабленных людей.
| Год | Серотип | Распространение |
|---|---|---|
| 1889–1890 | H2N8 | Тяжёлая эпидемия |
| 1900–1903 | H3N8 | Умеренная эпидемия |
| 1918–1919 | H1N1 | Тяжёлая пандемия [1] |
| 1933–1935 | H1N1 | Средняя эпидемия |
| 1946–1947 | H1N1 | Средняя эпидемия |
| 1957–1958 | H2N2 | Тяжёлая пандемия |
| 1968–1969 | H3N2 | Умеренная пандемия |
| 1977–1978 | H1N1 | Средняя пандемия |
| 1995–1996 | H1N1 и H3N2 | Тяжёлая пандемия |
Читайте также: