
Трансдукция у бактерий и вирусов
Обновлено: 24.04.2024
Механизм ее заключается в том, что в процессе внутриклеточного размножения фага в его головку может быть случайно включен вместо фаговой ДНК фрагмент бактериальной ДНК, равный по длине фаговой. Это вполне возможно, так как в инфицированной клетке биосинтез ее ДНК блокирован, а сама ДНК подвергается распаду. Таким образом в процессе репродукции фага возникают дефектные вирионы, у которых в головках вместо собственной геномной ДНК содержится фрагмент ДНК бактерии. Такие фаги сохраняют инфекционные свойства. Они адсорбируются на бактериальной клетке, вводят в нее ДНК, содержащуюся в головке, но при этом размножения фага не происходит. Введенная в клетку реципиента донорная ДНК (фрагмент хромосомы донора), если она содержит гены, отсутствующие у реципиента, наделяет его новым признаком. Этот признак будет зависеть от того, какой ген (гены) попал в головку трансдуцирующего фага. В случае рекомбинации привнесенного фагом фрагмента ДНК донора с хромосомой клетки-реципиента этот признак наследственно закрепляется.
Похожие книги из библиотеки

Йога. 7 духовных законов. Как исцелить свое тело, разум и дух
В этой книге знаменитый духовный лидер современности и автор десятков бестселлеров Дипак Чопра и его единомышленник, врач-невропатолог Дэвид Саймон, рассказывают о семи духовных законах йоги – глубоком смысле древней практики, способном привести вас к счастливой, гармоничной и полноценной жизни. Все мы знаем о том, как полезна йога: она дарит нам гибкость, мышечный тонус, выносливость, гармонию с самим собой и жизненную силу. Но все это – лишь часть удивительной силы, которой обладает эта духовная практика. Медитация, мантры, дыхательные упражнения и последовательности поз способны не только укрепить ваше здоровье, но и полностью преобразить вашу жизнь. Эта книга позволит вам заниматься йогой вдумчиво и осознанно, чтобы раскрыть свой творческий потенциал, научиться любить, сострадать и добиваться успеха во всем, чем бы вы ни занимались.

Я не могу уснуть. Уникальная система избавления от бессонницы за 5 недель
Замечали ли вы, что чем больше вы стараетесь уснуть, тем сложнее вам это дается? Эффективная методика, описанная в книге Гая Мэдоуса, покажет вам, как перестать бороться и начать спать. Используя сочетание техник осознанности и Терапии Принятия и Ответственности (ТПО) доктора Мэдоуса, а также применяя удобно расписанный по неделям курс избавления от бессоницы, вы сможете освободиться от проблем со сном, вне зависимости от того, страдаете ли вы от редких бессонных ночей или же продолжительной бессонницы. Инновационные методы Гая Мэдоуса имели беспрецедентный успех в США и были рекомендованы абсолютно всеми клиентами семинаров и курсов доктора. По многочисленным просьбам его высокоэффективный и естественный способ лечения бессонницы представлен в виде этой книги и теперь доступен в России. Внимание! Информация, содержащаяся в книге, не может служить заменой консультации врача. Необходимо проконсультироваться со специалистом перед применением любых рекомендуемых действий.

Лечение и чистка суставов. Классические и нетрадиционные методы

Новейший справочник медицинской сестры
Новейший справочник медицинской сестры является наиболее полным изданием, подготовленным в соответствии с требованиями национального стандарта РФ ГОСТ Р 52623.3-2015, вступившем в силу 1 марта 2016 г. В книге понятно и доступно излагается стандарт технологии выполнения простых медицинских процедур терапевтического, хирургического и травматологического профилей. Издание предназначено для среднего медицинского персонала отраслевых учреждений и организаций, также будет полезно и широкому кругу читателей.

Здоровая спина за 10 минут в день
Боль в спине является причиной большого числа снижений и потери трудоспособности. Общие советы, направленные на то, чтобы спина человека функционировала как можно лучше и дольше, кратко изложены в начальных главах книги; затем подробно описаны рекомендуемые конкретные упражнения, которые объединены в 7 групп в зависимости от целевого назначения или методики их выполнения.

М**ак не ходит в одиночку
Мы работаем с ними бок о бок. Сидим рядом в вагоне метро. Живем под одной крышей. И, о боже, порой видим их в зеркале! Клавдия Хохбрун, психиатр с многолетним стажем, называет этих людей мудаками, но, по сути, речь идет о 9 типах трудных характеров. В той или иной степени их черты присутствуют в каждом человеке. Именно поэтому эта книга так полезна. Она учит понимать, принимать и блокировать неприятные аспекты личности окружающих. Несмотря на легкий стиль, сдобренный немецким юмором, все рекомендации автора основаны на исследованиях в области психиатрии. Недаром Клавдия Хохбрун более 20 лет проработала в судебно-психиатрической системе, имея дело с особо опасными преступниками. Эта книга научит вас: • подмечать деструктивные черты характера у окружающих и блокировать их; • понимать настоящие причины разногласий с коллегами, руководителем или близкими людьми; • эффективным стратегиям общения с трудными людьми; • держать под контролем собственных тараканов; • радоваться жизни.

Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная трансдукция. Феномен лизогении.
Трансдукция — перенос бактериофагом в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг. Трансдуцирующий бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент).
Образовавшийся фаг и есть частица, вызывающая неспецифическую (общую) трансдукцию. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены.
При неспецифической трансдукции фагом может быть перенесён любой фрагмент ДНК хозяина, а при специфической лишь строго определённые фрагменты ДНК. Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при транедукции. При абортивной трансдукции внесённый фрагмент ДНК донора не встраивается в генофор реципиента, а остаётся в цитоплазме, где его ДНК транскрибируется, но не реплицируется. Это приводит к тому, что при клеточном делении он передаётся только одной из дочерних клеток (то есть наследуется однолинейно) и затем теряется в потомстве.
Свойства трансдуцирующих фаговых частиц следующие:
• Частицы несут лишь часть ДНК фага, то есть не являются функциональными вирусами, а скорее ёмкостями, переносящими фрагменты бактериальной ДНК.
• Подобно прочим дефектным вирусам, частицы не способны к репликации.
• Трансдуцирующие фаги могут содержать какую-либо часть хромосомы хозяина с генами, дающими реципиентной бактерии некоторые преимущества (например, гены устойчивости к антибиотикам или гены, кодирующие способность к синтезу различных веществ). Подобное приобретение бактериями новых свойств получило название феномен лизогении.
• Феномен трансдукции может быть использован для картирования бактериальной хромосомы, если следовать тем же принципам, что и при картировании с использованием феномена трансформации.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Перенос бактериальной ДНК. Конъюгация бактерий. F-фактор бактерии.
Длительное время считали, что бактерии — изолированные генетические системы, и каждая особь имеет одного (и только одного) родителя, то есть их изменчивость вызвана лишь мутациями. Никто не мог себе представить, что, подобно гаметам высших организмов, бактерии способны обмениваться генетическим материалом и, по аналогии с половым размножением, давать начало потомству с новыми свойствами. Однако Лёдерберг и Тёйтем (1946), высевая на минеральную среду, предварительно смешав два типа мутантов Escherichia coli (один нуждался в биотине и метионине, другой — в треонине и лейцине), показали, что у бактерий генетические рекомбинанты со способностью к синтезу всех четырёх факторов роста возникают с частотой 10"6, хотя теоретическая вероятность реверсии по двум генам составляет 10-14-10-16 на генерацию. Это противоречие было снято установлением факта прямой передачи генетического материала от донорной клетки к реципиентной (конъюгация). Помимо конъюгации, передача генетического материала у бактерий может осуществляться также с помощью трансформации и трансдукции (рис. 4-16).
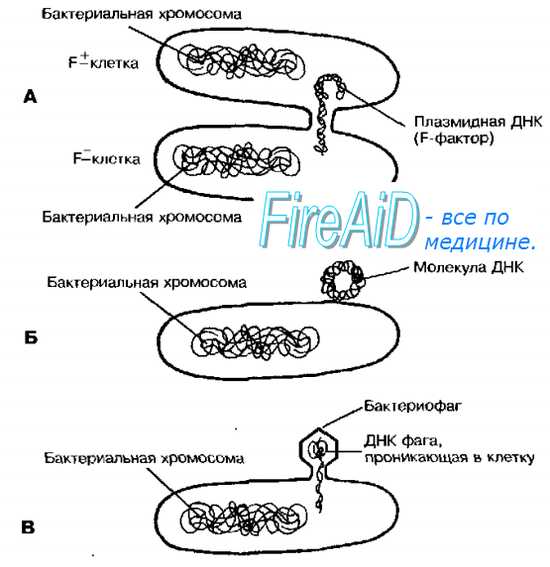
Рис. 4-16. Механизмы перенос бактериальной ДНК. Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б) трансдукция с помощью фагов (В).
Конъюгация бактерий. F-фактор бактерии.
Конъюгация — прямой перенос фрагмента ДНК от донорских бактериальных клеток к реципиентным при непосредственном контакте этих клеток. Биологическая значимость этого процесса стала проясняться после внедрения в медицинскую практику антибиотиков. Устойчивость к антибиотикам можно получить в результате мутации, что происходит один раз на каждые 106 клеточных делений. Однако, однажды изменившись, генетическая информация может быстро распространяться среди сходных бактерий благодаря конъюгации, поскольку каждая третья из близкородственных бактерий способна именно к этому типу генетического переноса. Для реализации процесса необходим F-фактор — плазмида, кодирующая информацию, необходимую для конъюгации.
Конъюгация требует наличия двух типов клеток: доноров (F + ), обладающих F-фактором, и реципиентов (F - ), не обладающих им. При скрещивании клеток F - и F + фактор фертильности передаётся с частотой, близкой к 100%.
Фактор переноса содержит гены специальных и необходимых при конъюгации структур — F-пилей и ряд других генов, вовлечённых в процесс взаимодействия с F - -клетками.
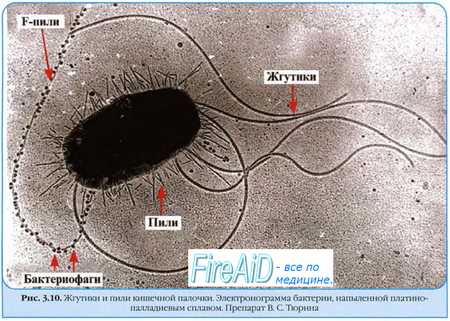
Первый этап конъюгации — прикрепление клетки-донора к реципиенту с помощью F-пилей. Затем между клетками формируется конъюгационный мостик, через который передаётся F-фактор, а также и другие плазмиды, автономно пребывающие в цитоплазме донора. При попадании F-фактора в реципиентную клетку она становится F+ и приобретает способность передавать фактор фертильности другим F - -клеткам. Подобный механизм обеспечивает приобретение популяционно. устойчивости к антибактериальным агентам.
В популяции клеток, содержащих F-плазмиду, только те, в которых она интегрирована в бактериальную хромосому (Hft + -клетки), способны быть донорами хромосомной ДНК. При перенос генетического материала бактериальная ДНК реплицируется, начиная от места включения F-фактора, одна цепь ДНК переносится в реципиентную F'-клетку двигаясь 5'-концом вперёд тогда как другая остаётся в Hfr + -клетке, то есть донор сохраняет своё генетическое постоянство. После начала конъюгации хромосомный материал переносится, начиная от генов, близких к начальной точке транспорта (рис. 4-17).
В бактерии-реципиенты обычно попадают первые из переносимых генов, размер которых зависит от времени, в течение которого проходила конъюгация, и очень редко — все гены. Позже всех переносится участок плазмиды, содержащий ген переноса кодирующий F-пили. Поскольку полная трансмиссия — явление редкое, реципиентная клетка при Hfr-конъюгации обычно остаётся F-. Вслед за процессом переноса в клетке-реципиенте происходит гомологичная рекомбинация между донорской ДНК и собственной ДНК реципиента.
Процесс конъюгации может происходить только при соблюдении ряда условий.
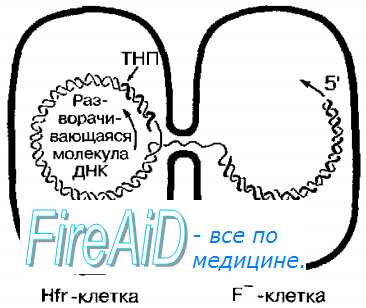
Рис. 4-17. Hfr-конъюгация. Hfr-бактерии функционируют при конъюгации как доноры. Перенос ДНК осуществляется линейно и начинается с удвоения места включения F-фактора (то есть с удвоения точки начала переноса — ТНП).
• На поверхности реципиентных бактерий должны быть рецепторы пилей, имеющие существенное сродство ( к F-пилям, что позволяет образовать стабильную связь между пилями и рецепторами.
• Для эффективной конъюгации у F-фактора должна быть точка начала репликации, распознаваемая репликативными системами хозяина.
• Эффективность Hfr-конъюгации зависит от величины гомологии ДНК. Перенос негомологичного хромосомного материала донора не приведёт к его интеграции с ДНК реципиента.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Мигрирующие генетические элементы бактерий. Транспозоны. Бактериофаги, как мигрирующие генетические элементы.
Мигрирующие генетические элементы — отдельные участки ДНК, способные осуществлять собственный перенос (транспозицию) внутри генома. Транспозиция связана со способностью мигрирующих элементов кодировать специфический фермент рекомбинации — транспозазу.
Вставочные (инсерционные) последовательности [IS-элементы (англ. insertion, вставка, + sequence, последовательность)] — простейший тип мигрирующих элементов (рис. 4-15, А); их величина не превышает 1500 пар оснований (в среднем 800-1400). IS-элементы самостоятельно не реплицируются и не кодируют распознаваемых фенотипических признаков. Содержащиеся в них гены обеспечивают только их перемещение из одного участка в другой.
Основные функции IS-последовательностей — регуляция активности генов бактериальной клетки (могут инактивировать гены, в которые включились, или, встраиваясь в хромосому, проявлять эффект промотора, включающего либо выключающего транскрипцию соответствующих генов), индукция мутаций типа делеций или инверсий (при перемещении) и дупликаций (при встраивании в хромосому), координация взаимодействий плазмид, траспозонов и профа-гов (как между собой, так и бактериальной хромосомой).
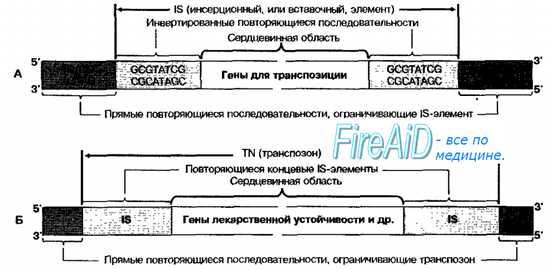
Рис. 4-15. Инсерционная последовательность (А), транспозон (Б).
Транспозоны (Tn-элементы) состоят из 2000-25 000 пар нуклеотидов, содержат фрагмент ДНК, несущий специфические гены, и два концевых IS-элемента (рис. 4-15, Б). При включении в ДНК бактерий транспозоны вызывают дупликации, при выходе из определённого участка ДНК— делеций, при выходе и включении обратно с поворотом фрагмента на 180 градусов— инверсии. Транспозоны не способны к самостоятельной репликации и размножаются только в составе бактериальной хромосомы.
Каждый транспозон обычно содержит гены, привносящие важные для бактерии характеристики типа множественной устойчивости к антибактериальным агентам. Поскольку транспозоны содержат гены, определяющие фенотипически выраженные признаки (например, устойчивость к антибиотикам), то их легче обнаружить, чем IS-элементы. В общем, для транспозонов характерны те же гены, что и для плазмид (гены устойчивости к антибиотикам, токсинообразования, дополнительных ферментов метаболизма).
Умеренные и дефектные бактериофаги также могут быть факторами изменчивости, напоминая по своим свойствам интегрированные плазмиды. Встраиваясь в бактериальную хромосому в форме профага (провируса), они вызывают лизогенизацию бактерий, которые могут приобретать новые свойства.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Отличается от неспецифической тем, что в этом случае трансдуцирующие фаги всегда переносят только определенные гены, а именно, те из них, которые располагаются в хромосоме лизогенной клетки слева от attL или справа от attR. Специфическая трансдукция всегда связана с интеграцией умеренного фага в хромосому клетки-хозяина. При выходе (исключении) из хромосомы профаг может захватить ген с левого или правого фланга, например или gal, или bio. Но в этом случае он должен лишиться такого же размера своей ДНК с противоположного конца, чтобы ее общая длина оставалась неизменной (иначе она не может быть упакована в головку фага). Поэтому при такой форме исключения образуются дефектные фаги: ?dgal или ?dbio.
Специфическую трансдукцию у E. coli осуществляет не только фаг лямбда, но и родственные ему лямбдоидные и другие фаги. В зависимости от места расположения сайтов attB на хромосоме они при своем исключении могут включать различные бактериальные гены, сцепленные с профагом, и трансдуцировать их в другие клетки. Встраивающийся в геном материал может замещать до 1 /3 генетического материала фага.
Трансдуцирующий фаг в случае инфицирования реципиентной клетки интегрируется в ее хромосому и привносит в нее новый ген (новый признак), опосредуя не только лизогенизацию, но и лизогенную конверсию.
Таким образом, если при неспецифической трансдукции фаг является только пассивным переносчиком генетического материала, то при специфической фаг включает этот материал в свой геном и передает его, лизогенизируя бактерии, реципиенту. Однако лизогенная конверсия может произойти и в том случае, если геном умеренного фага содержит такие собственные гены, которые у клетки отсутствуют, но отвечают за синтез существенно важных белков. Например, способностью вырабатывать экзотоксин обладают только те возбудители дифтерии, в хромосому которых интегрирован умеренный профаг, несущий оперон tox. Он отвечает за синтез дифтерийного токсина. Иначе говоря, умеренный фаг tox вызывает лизогенную конверсию нетоксигенной дифтерийной палочки в токсигенную.
Фаги размножаются только за счет паразитирования в микробной клетке. Их размножение в бульонной культуре приводит к тому, что культура, бывшая перед добавлением фага мутной, через несколько часов инкубации при температуре 37 °C становится прозрачной. На плотных средах фаги обнаруживают либо с помощью спот-теста (англ. spot – пятно), либо методом агаровых слоев, предложенным А. Грациа (1936). В первом случае на поверхность агара в чашке засевают культуру, а затем на нее наносят каплю содержащего фаг материала. Если в нем содержится много вирионов, то на месте нанесения капли будет большое стерильное пятно (рис. 84, 1).
Метод агаровых слоев заключается в следующем. Вначале в чашку наливают слой питательного агара. После застывания на этот слой добавляют 2 мл расплавленного и охлажденного до 45 °C агара 0,7 %-ного, в который предварительно добавляют каплю концентрированной суспензии бактерий и определенный объем суспензии фага. После того как верхний слой застынет, чашку помещают в термостат. Бактерии размножаются внутри мягкого слоя агара, образуя сплошной непрозрачный фон, на котором хорошо видны колонии фага в виде стерильных пятен (рис. 84, 2). Каждая колония образуется за счет размножения одного исходного фагового вириона. Применение этого метода позволяет: а) путем подсчета колоний точно определить количество жизнеспособных фаговых вирионов в данном материале;
б) по характерным признакам (размер, прозрачность и др.) изучать наследственную изменчивость у фагов.
По спектру своего действия на бактерии фаги подразделяются на поливалентные (лизируют родственные бактерии, например поливалентный сальмонеллезный фаг лизирует почти все сальмонеллы), монофаги (лизируют бактерии только одного вида, например фаг Vi-I лизирует только возбудителей брюшного тифа) и типоспецифические фаги, которые избирательно лизируют отдельные варианты бактерий внутри вида. С помощью таких фагов производится наиболее тонкая дифференциация бактерий внутри вида, с разделением их на фаговарианты. Например, с помощью набора фагов Vi-II возбудитель брюшного тифа делится более чем на 100 фаговариантов. Поскольку чувствительность бактерий к фагам является относительно стабильным признаком, связанным с наличием соответствующих рецепторов, фаготипирование имеет важное диагностическое и эпидемиологическое значение.
Читайте также: