
Транскрипция вирусной днк это
Обновлено: 23.04.2024
Транскрипция вируса в клетке. Трансляция вирусов.
Транскрипция +РНК-содержащих вирусов. Функции мРНК выполняет геном (+РНК), поэтому у таких вирусов для синтеза вирусных белков (трансляция) нет необходимости в процессе транскрипции. Другими словами, у +РНК-содержащих вирусов транскрипция отсутствует.
Транскрипция -РНК-содержащих вирусов и вирусов, имеющие две нити РНК. Функции мРНК выполняют транскрипты, комплементарные -РНК вириона. Поэтому у таких вирусов транскрипция существует как самостоятельный этап репродуктивного цикла. Для образования транскриптов в составе вирионов имеется собственная РНК-полимераза (транскриптаза).
Транскрипция ДНК-вирусов. Транскрипция — самостоятельный этап репродуктивного цикла, так как геном ДНК-вирусов должен транскрибироваться для образования мРНК. Вирусы, репродуцирующиеся в ядре (например, герпес- и аденовирусы) для этой цели используют клеточную ДНК-зависимую РНК-полимеразу (транскриптазу). Вирусы, репродуцирующиеся в цитоплазме (например, поксвирусы) лишены такой способности и содержат (как и вирусы с -РНК) собственную транскриптазу.
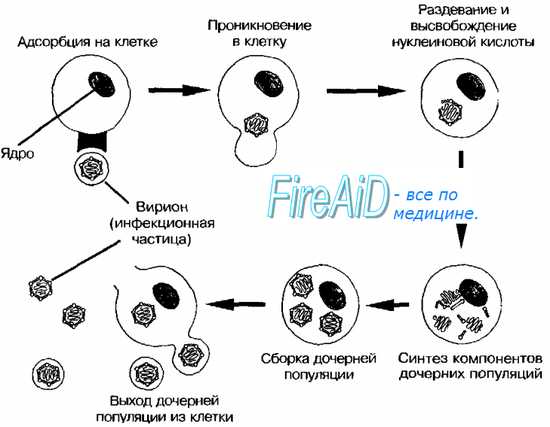
Рис. 2-3. Основные этапы репродукции вирусов.
Трансляция вирусов
Вирусные геномы кодируют синтезы двух классов белков: структурные белки входят в состав дочерних популяций, а неструктурные белки обслуживают процессы репродукции, но не входят в состав дочерних популяций (ингибиторы синтеза клеточных РНК и белков, протеазы и др.).
Трансляция РНК-содержащих вирусов. Поскольку вирусный геном кодирует несколько белков, то возможно два варианта трансляции:
1) каждый полипептид синтезируется отдельно от других (тога- и ретровирусы);
Некоторые вирусы используют оба этих механизма. Полипептиды, образующиеся при обоих вариантах трансляции, могут подвергаться посттрансляционной модификации (гликозилирование, фосфорилирование или сульфатирование).
Трансляция ДНК-содержащих вирусов. В трансляционных процессах доминирует трансляция отдельных мРНК, кодирующих индивидуальные полипептиды. В отдельных случаях (например, у аденовирусов) не менее трёх белков образуются путём нарезания общего полипептида-предшественника.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Транскрипция вируса в клетке. Трансляция вирусов.
Транскрипция +РНК-содержащих вирусов. Функции мРНК выполняет геном (+РНК), поэтому у таких вирусов для синтеза вирусных белков (трансляция) нет необходимости в процессе транскрипции. Другими словами, у +РНК-содержащих вирусов транскрипция отсутствует.
Транскрипция -РНК-содержащих вирусов и вирусов, имеющие две нити РНК. Функции мРНК выполняют транскрипты, комплементарные -РНК вириона. Поэтому у таких вирусов транскрипция существует как самостоятельный этап репродуктивного цикла. Для образования транскриптов в составе вирионов имеется собственная РНК-полимераза (транскриптаза).
Транскрипция ДНК-вирусов. Транскрипция — самостоятельный этап репродуктивного цикла, так как геном ДНК-вирусов должен транскрибироваться для образования мРНК. Вирусы, репродуцирующиеся в ядре (например, герпес- и аденовирусы) для этой цели используют клеточную ДНК-зависимую РНК-полимеразу (транскриптазу). Вирусы, репродуцирующиеся в цитоплазме (например, поксвирусы) лишены такой способности и содержат (как и вирусы с -РНК) собственную транскриптазу.
Рис. 2-3. Основные этапы репродукции вирусов.
Трансляция вирусов
Вирусные геномы кодируют синтезы двух классов белков: структурные белки входят в состав дочерних популяций, а неструктурные белки обслуживают процессы репродукции, но не входят в состав дочерних популяций (ингибиторы синтеза клеточных РНК и белков, протеазы и др.).
Трансляция РНК-содержащих вирусов. Поскольку вирусный геном кодирует несколько белков, то возможно два варианта трансляции:
1) каждый полипептид синтезируется отдельно от других (тога- и ретровирусы);
Некоторые вирусы используют оба этих механизма. Полипептиды, образующиеся при обоих вариантах трансляции, могут подвергаться посттрансляционной модификации (гликозилирование, фосфорилирование или сульфатирование).
Трансляция ДНК-содержащих вирусов. В трансляционных процессах доминирует трансляция отдельных мРНК, кодирующих индивидуальные полипептиды. В отдельных случаях (например, у аденовирусов) не менее трёх белков образуются путём нарезания общего полипептида-предшественника.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Транскрипция вирусов. Характеристика транскрипции вирусов.
У оц(+)РНК-вирусов репликация начинается с полной или частичной трансляции генома без промежуточных этапов. У всех других классов вирусов процесс экспрессии вирусного генома начинается с транскрипции мРНК.
В случае с ДНК-вирусами, реплицирующимися в ядре, эту функцию выполняет клеточная ДНК-зависимая РНК-полимераза II. Все другие вирусы нуждаются в уникальной и специфичной транскриптазе, которую кодирует вирус, и она является структурным компонентом вириона. Двуцепочечные ДНК-вирусы, которые размножаются в цитоплазме, имеют ДНК-зависимую РНК-полимеразу, тогда как двуцепочечные РНК-вирусы имеют РНК-зависимую РНК-полимеразу, специфичную для двуцепочечных РНК, а негативно полярные односпиральные РНК-вирусы носят РНК-зависимую РНК-полимеразу, специфичную для одноцепочечных РНК.
Транскрипция вирусных ДНК и РНК контролируется сложными регуляторными механизмами и, прежде всего, продуктами экспрессии регуляторных генов.
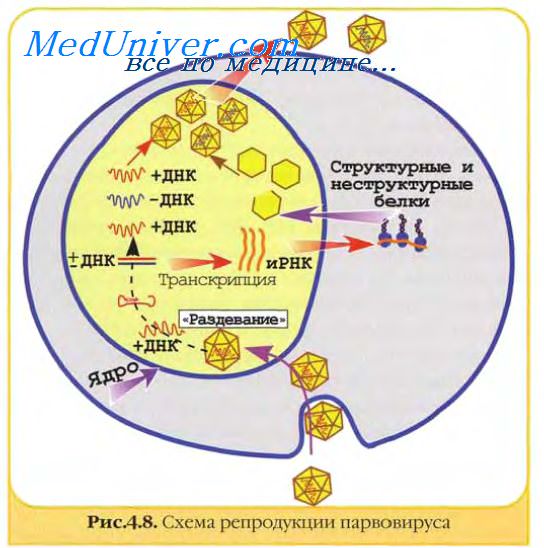
Транскрипты, которые образовались особенно в ранней стадии инфекции, считываются полностью на поздней стадии инфекции с образованием ряда длинных транскриптов с различными функциями.
Общими закономерностями транскрипции некоторых вирусов с дцДНК-геномом являются: транскрипция ранних и поздних генов с разных цепей ДНК, наличие перекрывающихся генов, разных рамок считывания и интронов.
У РНК-вирусов регуляция транскрипции в общем происходит менее сложно, чем у ДНК-вирусов. Временные различия транскрипции разных генов выражены не так отчетливо. У большинства семейств вирусов с оц(+)РНК геномная РНК служит мРНК, и для транскрипции-репликации РНК требуется только образование негативной цепи. Однако для ретровирусов и вирусов с несегментированным негативным РНК-геномом существут другие механизмы регуляции транскрипции.
Различные виды мРНК имеют различный период полураспада, что может также служить одним из возможных уровней регуляции вирусной репликации.
Первичные РНК-транскрипты, образующиеся на ДНК-геномах в ядре перед выходом в цитоплазму, претерпевают серию посттрансляционных изменений (кэпирование, аденилирование, метилирование, делетирование и сплайсинг).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.
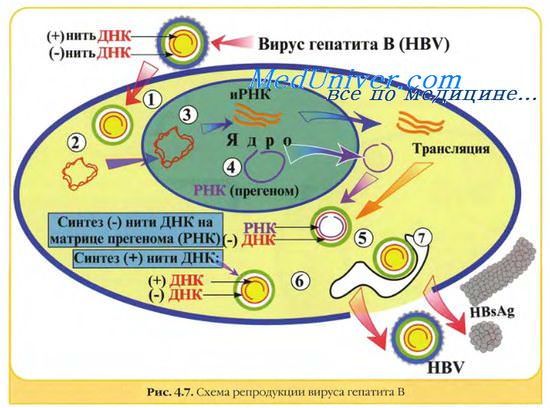
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Транскрипция – в вирусологии этот термин обозначает процессы синтеза комплементарных (пространственно-взаимодополняющих) геному РНК (рибонуклеиновых кислот), обладающих информационными функциями (иРНК). Иными словами. транскрипция в данном случае – это переписывание информации с ДНК на РНК и образование иРНК (РНК с информационной функцией) [3] [1] .
Совокупность процессов транскрипции является одним из основных этапов в системе синтеза компонентов вируса в общей структуре репродукции вируса [2] .
Отсутствие транскрипции
Наличие транскрипции в процессах репродукции вируса зависит от типа генома вириона. Она присутствует у ДНК-содержащих вирусов, РНК-содержащих вирусов с негативным геномом (минус-РНК), РНК-содержащих ретровирусов [3] .
Процессы транскрипции в реализации генетической информации отсутствуют у вирусов, содержащих плюс-РНК, поскольку в этом случае функции иРНК выполняет вирусная нуклеиновая кислота. В том числе отсутствие транскрипции отмечается и у вирусов, содержащих геном, состоящий из комплементарных друг другу плюс-РНК и минус-РНК. В этом случае функции иРНК берет на себя плюс-РНК нить [1] .
Ферменты, запускающие транскрипцию
Процессы транскрипции запускаются при помощи определенных ферментов [3] [1] .
Вирусы, содержащие негативный РНК-геном (минус-РНК), необходимую для транскрипции РНК-зависимую РНК-полимеразу привносят в составе фрагментов капсидов, поскольку при заражении клеток минус-РНК геномные вирусы полностью не раздеваются [3] [1] .
У вирусов с ДНК-геномами транскрипция осуществляется с помощью ДНК-зависимой РНК-полимеразы. Она отражает нуклеотидную последовательность ДНК в нуклеотидной последовательности иРНК. Для этого часто используется фрагмент клетки-хозяина [3] [1] .
Обратная транскрипция
Обратная транскрипция – это синтез ДНК на матрице РНК. Открытие этого процесса явилось значительной поправкой в центральную догму молекулярной биологии об одно направленности движения генетической информации от ДНК к белку. (ДНК→РНК →белок) [1] .
К обратной транскрипции способны:
- вирусы – возбудители злокачественных новообразований;
- вирус иммунодифицита человека (ВИЧ) [1] .
Эти вирусы объединяют в группу ретровирусы. Их геном представляет собой однонитевую плюс-РНК и кодирует фермент ревертазу (обратная транскриптаза). Последняя, обладает свойствами РНК-зависимой и ДНК-зависимой ДНК-полимеразы [1] .
Ревертаза транскрибирует плюс-РНК вирусов, копируя ее сначала в однонитевую минус-ДНК, а затем в двухнитевую ДНК (кДНК) [1] .
Отмечена важная особенность транскрипта кДНК – это возможность его превращения в провирус, то есть формирование способности встраиваться в хромосому клетки [1] .
Интеграция провируса (вирусной ДНК) – это уникальное событие в жизни ретровирусов абсолютно необходимое для их последующей репликации в зараженных клетках [1] .
Читайте также:
- Какой путь передачи наиболее характерен для вирусного гепатита в тесты
- Облучатель рециркулятор медицинский отзывы помогает при коронавирусе
- Циклоферон при вирусной инфекции как быстро помогает
- Результат анализа на гепатит отрицательный что это значит
- Есть ли уголовная ответственность за заражение гепатитом с