
В клетке возбудителя чумы нет рибосом цитоплазмы
Обновлено: 06.05.2024
• Нуклеоид бактерий выглядит как диффузная масса ДНК, однако для него характерна высокая упорядоченность и неслучайное расположение генов
• У бактерий нет нуклеосом, однако организации ДНК способствуют различные белки, связанные с нуклеоидом
• Подобно тому как это имеет место для ядра и цитоплазмы эукариотической клетки, у бактерий транскрипция происходит по всей массе нуклеоида, трансляция — на его периферической зоне
• Важную роль в организации нуклеоида играет РНК полимераза
Фундаментальное отличие клеток прокариот от клеток эукариот заключается в отсутствии у них ядерной оболочки. Присутствие ядерной мембраны у эукариот обеспечивает существование компартментов, которые разделяют процессы транскрипции и трансляции. У прокариот эти процессы не разделены мембраной, и мРНК может транслироваться во время транскрипции. Одновременное протекание этих процессов имеет важные последствия для регуляции активности некоторых генов.
Как показано на рисунке ниже, хромосомальная ДНК бактерий имеет вид аморфной массы, нуклеоида, занимающего большую часть объема в центре цитоплазмы. Нуклеоид состоит из хромосомальной ДНК и связанных с ней белков. Бактерии не содержат нуклеосом, которые участвуют в упаковке ДНК хромосом эукариотов и архей. Однако бактериальная ДНК компактна и упакована с участием многочисленных белков, ассоциированных с нуклеоидом, которые перчислены на рисунке ниже.
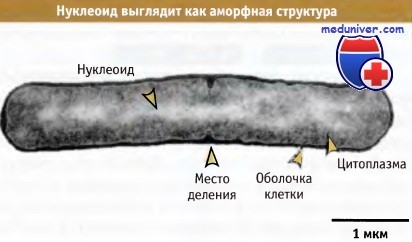
Электронная микрофотография, демонстрирующая,
что нуклеоид представляет собой диффузную массу, находящуюся внутри клетки бактерии.
К числу наиболее важных из этих белков относятся топоизомеразы. Они контролируют суперспирализацию ДНК, которая играет важную роль в ее компактизации, и обеспечивают протекание таких процессов, как репликация и транскрипция, для которых требуется раскручивание молекулы ДНК. Белки семейства SMC, поддерживающие структурную организацию хромосом, также участвуют в организации нуклеоида. Об этом свидетельствует фенотип соответствующих мутантов, однако конкретный механизм их участия остается неясным.
В клетках эукариот белки, близкие к SMC, участвуют в скреплении хромосом между собой и их конденсации в митозе и мейозе. Эти белки различной природы, связанные с нуклеоидом, участвуют в поддержании необходимого уровня его суперспирализации и компактизации. Однако предстоит еще выяснить, каким образом достигается и поддерживается такое состояние гомеостаза нуклеоида.
Хотя нуклеоид обладает аморфной структурой, отдельные гены располагаются в нем упорядоченно. Положение генов в нуклеоиде отражает их относительное расположение на карте хромосомы. По счастью, первое подтверждение этого было получено при исследовании свойств мутантов бактерий В. subtilis, дефектных по гену spoIIIE. Мутант этого организма не способен правильно сегрегировать хромосому при асимметрическом делении, которое сопровождает ранние стадии образования споры. Вместо этого септа деления замыкается вокруг одной копии хромосомы. У этого мутанта определенные гены почти всегда попадают в небольшой компартмент, поблизости от полюса, в то время как другие из него всегда исключаются.
Это наблюдение позволяет предполагать, что до деления хромосома всегда находится в определенном месте и в определенной ориентации.
Прямые данные были получены в исследованиях с использованием гибридизации in situ и флуоресцентной метки (FISH). Этот метод позволяет непосредственно отслеживать положение в клетке определенных генов. Однако при его использовании, перед гибридизацией зонда с ДНК, необходима фиксация препаратов и проведение других жестких воздействий. Еще один подход заключается в использовании конъюгата зеленого флуоресцирующего белка с белком LacI, связывающимся с ДНК. Этот конъюгат может присоединяться к сайтам связывания, находящимся в разных местах клетки. На основании всех этих экспериментов было показано, что гены не диффундируют по бактериальной клетке свободно, а локализованы в определенных, строго ограниченных местах.
Вообще говоря, область хромосомы, содержащая oriC, находится на одном конце нуклеоида, а область, содержащая terC, — на противоположном. Гены, которые на генетической карте расположены между двумя этими точками, распределены по нуклеоиду более или менее пропорционально.
У бактерий в аппарате транскрипции используется одна каталитическая основная РНК-полимераза, состоящая из двух а-, одной b- и одной b-субъединиц. Специфичность промотора определяется на начальном уровне различными сигма (а) факторами, которые также необходимы для инициации транскрипции, однако после этого отщепляются от кора. Регуляция транскрипции осуществляется большим набором дополнительных регуляторов, которые обычно связываются с ДНК вблизи от промотора, с тем чтобы активировать или подавлять инициацию транскрипции. Другие факторы регуляции действуют на уровне терминации (прекращения) транскрипции или изменения стабильности мРНК.
Большая часть молекул основной РНК-полимеразы находится в нуклеоиде в центре клетки. Поэтому, вероятно, здесь в основном происходит транскрипция. Напротив, рибосомы и различные белки, принимающие участие в трансляции, сосредоточены по периферии клетки. Таким образом, даже при отсутствии ядерной оболочки, в бактериальной клетке транскрипция и трансляция пространственно разобщены, подобно тому как это имеет место в клетке эукариот. Однако существуют различные данные в пользу того, что иногда у бактерий транскрипция и трансляция тесно сопряжены друг с другом.
Эти данные не противоречат имеющимся результатам, которые свидетельствуют о том, что РНК-полимеразы и рибосомы локализованы в разных местах клетки. Возможно, что оба процесса происходят на границе центральной, или сердцевинной, и периферийной областях клетки. Пока мы мало знаем об организации центральной, или сердцевинной, и периферийной областей нуклеоида, так же как и о деталях общей организации этой структуры.
Белки, участвующие в организации нуклеоида Escherichia coli.
У большинства других бактерий вместо белков MukB, MukE и MukF присутствуют белки SMC (белки, поддерживающие структуру хромосом),
а также связанные с ними факторы, родственные когезину и конденсинам эукариот. Сегрегация хромосом после образования полярной септы при наступлении споруляции.
В холе споруляции В. subtilis клетки делятся асимметрично, образуя материнскую клетку и небольшую преспору.
Каждая клетка получает одну копию хромосомы. Сегрегация хромосом с образованием преспоры представляет собой двухэтапный процесс.
Вначале полярная разделительная септа замыкается вокруг хромосомы,
а затем белок SpoIIIE активно транспортирует оставшиеся 2/3 хромосомы в преспоровый компартмент.
У мутантов по гену spoIIIE только 1/3 хромосомы сегрегирует в преспору.
Анализ ДНК, захваченной в небольшой компартмент клеток мутанта по гену spoIIIE, показывает, что всегда захватывается специфический участок ДНК.
Это указывает на то, что до деления хромосома должна быть строго ориентирована и упорядочена.
На фотографиях, полученных во флуоресцентном микроскопе, представлены клетки спорулирующих spoIIIE-мутантов и клетки дикого типа, окрашенные на ДНК. Несмотря на отсутствие ядерной оболочки, аппараты транскрипции и трансляции локализуются в отдельных частях бактериальной клетки.
Представлены делящиеся клетки В. subtilis.
Они экспрессируют конъюгаты белка рибосомальной субъединицы RpsB с зеленым флуоресцирующим белком (GFP)
и субъединицы РНК-полимеразы RpoC с GFP-UV, обладающие зеленой и красной флуоресценцией соответственно.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Обзор
Автор
Редактор

Один монах, странствуя по белому свету, встретил Чуму, которая направлялась в его город.
— Ты куда это направляешься, Чума? — спросил он ее.
— Иду в твой родной город, — ответила она. — Мне нужно забрать там тысячу жизней.
Через некоторое время монах снова встретил Чуму на своем пути.
— Почему ты меня обманула тогда? — спросил он ее с укором. — Ты говорила, что должна забрать тысячу жизней, а забрала пять тысяч.
— Я тогда сказала тебе правду, — ответила Чума. — Я действительно забрала тысячу жизней. Остальные умерли от страха.
Жертвы чумы исчислялись сотнями тысяч и даже миллионами человек, вымирали города, становились безлюдными целые области, и ужас пандемий чумы затмевал ужасы всех войн, какие знала история человечества. Целые тысячелетия люди не понимали, что является источником заболевания [2].
Библия — одно из древнейших дошедших до нас свидетельств эпидемий чумы (1 книга Царств, глава 5; 4 книга Царств, глава 19, стихи 35–36). В мировой истории отмечают три пандемии этой болезни:
Бубонная форма чумы является наиболее распространенной формой заболевания и при отсутствии лечения приводит к гибели 40–60% заболевших. Легочная форма возникает либо как осложнение бубонной или септической форм, либо при вдыхании воздуха, зараженного возбудителем чумы. Если лечение не начинают в первые 24 часа после появления симптомов, смерть наступает через 48 часов [8].
В природе чумной микроб встречается практически на всех континентах, исключая Австралию, Антарктиду, а также Арктику, что обусловливает ежегодно регистрируемые случаи этой болезни. Стремительная эволюция микроорганизмов приводит к появлению популяций бактерий (штаммов), устойчивых к антибиотикам [9], что в случае с возбудителем чумы особенно опасно. Кроме того, этих бактерий могут использовать в качестве агента биотерроризма. Все вышесказанное объясняет необходимость изучения чумного микроба.
Возбудитель чумы Yersinia pestis — самая опасная бактерия в мире [10]. Что делает ее столь смертоносной?
Факторы вирулентности, или вооружен и очень опасен
Со времен открытия возбудителя чумы в 1894 году французом Александром Йерсеном и японцем Китасато Сибасабуро ученые пытались выяснить, что определяет патогенность Y. рestis. В результате многолетней тяжелой и рискованной работы, которая продолжается и по сей день, выделили следующие факторы патогенности возбудителя:
- белки внешней мембраны (Yersinia outer proteins — называемые Yop-белками, эффекторными белками, или комплексом Yop-вирулона) [11];
- комплекс области пигментации [12];
- активатор плазминогена [13];
- капсульный антиген [14];
- пили адгезии или pH6-антиген [15].
Белки внешней мембраны, или зачем возбудителю чумы шприц?

Рисунок 1. Схема действия системы секреции III типа.
Комплекс области пигментации, или может ли стать потребность в чем-либо фактором патогенности?
Активатор плазминогена, или двуликий Янус
При вдыхании чумных микробов (и развитии легочной чумы) этот белок обеспечивает быстрое размножение бактерий в тканях легких и приводит к развитию молниеносной пневмонии и отеку легких, тогда как в отсутствии Pla инфекция не развивается в смертельную пневмонию. Установлено, что активатор плазминогена нарушает постоянство внутренней среды организма хозяина и блокирует иммунные реакции, направленные на уничтожение патогена [27].
Капсульный антиген, или скользкий тип этот возбудитель чумы
Бактерии окружены капсулой из слизистого вещества (фракция I, Fra1), которая препятствует поглощению и обезвреживанию Y. pestis иммунными клетками организма-хозяина в процессе фагоцитоза. На выявлении этого вещества-антигена основаны многие современные методы лабораторной диагностики чумы, оно входит в состав многих экспериментальных химических вакцин против чумы. Однако позднее обнаружили популяции бактерий, лишенные капсулы [28]. Кроме того, слизистая капсула есть у многих других микроорганизмов, например, возбудителя сибирской язвы, туляремии. Капсульное вещество иерсинии образуют при температуре 37 °С.
Антигены, схожие с рН6, были обнаружены у ряда возбудителей, вызывающих менее опасные болезни — кишечные инфекции (Y. pseudotuberculosis [31], Y. enterocolitica [32], Escherichia coli [8]).
Температурный фактор, или то, что действительно имеет значение
Необходимо заострить внимание на особой роли температуры в физиологии чумного микроба. Именно при температуре 37 °С у него повышаются питательные потребности [33] и синтезируются практически все известные детерминанты вирулентности (рис. 2) [34]. У других бактерий подобная зависимость выражена в меньшей степени, что позволяет говорить о ведущей роли температурного фактора в вирулентности возбудителя чумы [8].
Геном или все важное внутри
Помимо хромосомы у чумного микроба есть плазмиды — внехромосомные участки ДНК [38]. Большинство белковых факторов вирулентности закодированы на плазмидах: эффекторные белки на плазмиде pCad; капсула — pFra; активатор плазминогена — рPla (pPst, pPCP). Плазмиды pFra и рPla обнаружены только у Y. pestis (видоспецифические), pCad является общей с возбудителем псевдотуберкулеза (родоспецифическая) [20].
Заключение
В настоящее время продолжается работа по выявлению новых, еще не изученных маркеров вирулентности [39]. С использованием 2D-электрофореза, масс-спектрометрии, полногеномного секвенирования проводят сравнительный анализ отличающихся по вирулентности популяций чумного микроба для выявления различий в их белковых спектрах и геномных последовательностях. Ранее не известные белки и участки генома становятся объектом пристального внимания и изучения как потенциальные детерминанты вирулентности.
Таким образом, патогенность возбудителя чумы — это множественный (полидетерминантный) признак. Соединение многих факторов в единое целое создает страшную угрозу чумных эпидемий, с противостоянием которым, однако, прогрессивное человечество успешно справляется.

Обзор
Рибосомы обладают избирательностью
Автор
Редактор

Спонсором приза зрительских симпатий выступил медико-генетический центр Genotek.
Белки принимают участие в самых разных клеточных процессах. Например, они могут быть строительными блоками для образования клеточных структур, могут катализировать протекание химических реакций в клетке, осуществлять транспорт молекул из клетки и обратно, а также выполнять некоторые другие важные функции. Понятно, что для нормального роста и развития любой клетке необходимо синтезировать белки. А для их производства, как известно, необходимы рибосомы, ведь именно они отвечают за этот процесс.
Рибосома эукариот состоит из двух субъединиц — большой (ее принято обозначать 60S) и малой (40S). Субъединицы построены из рибосомной РНК (рРНК) и рибосомных белков (РБ). Всего в эукариотической рибосоме находятся около 80 рибосомных белков и 4 типа рРНК .
Новые вехи в истории рибосомы
Новая волна интереса к рибосомам поднялась в начале 90-х годов прошлого столетия. Возможно, толчком к этому послужили две примечательные работы, посвященные исследованию рибосом на разных стадиях жизни эукариотических организмов — малярийного плазмодия и клеточного слизевика.
Малярийный плазмодий вызывает у человека тяжелое заболевание — малярию. Заражение происходит после укуса малярийного комара, в слюне которого находится возбудитель болезни. Отличительной особенностью плазмодия является то, что его жизненный цикл состоит из двух частей (рис. 1). Одна часть протекает в теле комара, другая — в теле человека [12].

Рисунок 1. Жизненный цикл малярийного плазмодия
[12], рисунок с изменениями
После укуса спорозоиты плазмодия — мелкие червеобразные подвижные клетки — попадают в кровоток. По кровотоку они достигают печени, где размножаются бесполым путем, превращаясь в одноядерные клетки, называемые мерозоитами. Голодные мерозоиты атакуют клетки крови — эритроциты — и проникают в них. Питаясь гемоглобином, мерозоиты продолжают бесполое размножение. Через некоторое время они разрушают эритроциты и снова выходят в кровь, где набрасываются на новые эритроциты. Это происходит циклично через каждые 48 часов. При разрушении эритроцитов в кровоток попадают и токсичные продукты жизнедеятельности мерозоитов, так что у человека через равные промежутки времени наблюдаются приступы лихорадки с ознобом и высокой температурой.
На стадии выхода из эритроцитов часть мерозоитов приобретает признаки половых клеток (мужских и женских) — образуются гаметоциты. Когда это происходит, плазмодий вступает во вторую стадию жизненного цикла, которая протекает в теле комара. Если комар выпивает кровь больного малярией человека, то в его организме гаметоциты окончательно созревают и превращаются в гаметы. Именно в теле комара гаметы сливаются, образуя оплодотворенную яйцеклетку. Для дальнейшего развития яйцеклетке необходимо попасть в кишечник комара, где она, проходя несколько стадий, образует спорозоиты. Малярийный плазмодий возвращается в то состояние, когда для дальнейшего развития ему снова необходимо попасть в тело человека. Как только комар кусает свою жертву, спорозоиты попадают в кровь, и цикл повторяется.
Так вот, изучая рибосомы малярийного плазмодия, ученые выяснили, что рРНК малой субъединицы рибосомы кодируется двумя генами [13]. Оказалось, что в клетках плазмодия, живущего в москитах, образуется продукт одного рРНК-гена, а в клетках, попавших в млекопитающее или человека, — продукт другого гена рРНК. Выходило, что, во-первых, состав рибосомы может меняться, а, во-вторых, что эти изменения зависят от состояния внешней среды и от фазы жизненного цикла организма.
Похожие результаты, но уже в отношении других компонентов рибосомы — рибосомных белков — несколькими годами позже получили другие ученые.
Dictyostelium discoideum — слизевик, обитающий в верхних слоях почвы, — является излюбленным модельным организмом молекулярных биологов.

Рисунок 2. Этапы жизненного цикла слизевика Dictyostelium discoideum

Рисунок 3. Изменение клеточного состава рибосом слизевика Dictyostelium discoideum в течение жизненного цикла
[12], рисунок с изменениями
Проведенные опыты позволили ввести понятие гетерогенной популяции рибосом — то есть такой популяции, в которой один или несколько структурных компонентов (рибосомные белки или рибосомные РНК) зрелой органеллы подвергались изменениям (качественным или количественным) в течение жизненного цикла клетки.
Рибосома — молекулярный фильтр?
Дело в том, что в мРНК есть не только области, кодирующие аминокислотную последовательность белка, но и области, имеющие чисто регуляторное назначение (рис. 4). С их помощью начинать синтезировать белок рибосома может по двум разным механизмам.

рисунок автора статьи
Первый носит название кэп-зависимого, или сканирующего. На одном из концов молекулы мРНК имеется специальная структура — кэп (молекула 7-метилгуанозина), на который перед началом синтеза садится рибосома. Сев, она движется вдоль молекулы мРНК, сканируя ее на предмет старт-кодона, кодирующего первую аминокислоту белка. Найдя старт-кодон, рибосома начинает синтез.
Гипотезы приобретают подтверждение
Несомненно, двадцать первый век — век высоких технологий, позволяющих человеку совершать то, что раньше казалось выдумкой ученых-фантастов. И молекулярные биологи активно пользуются результатами научно-технического прогресса в своих научных изысканиях. Благодаря интенсивному развитию техники, несколько лет назад сразу обе гипотезы нашли свое экспериментальное подтверждение [20]. С помощью самых современных технологий установили, что рибосомы модельного объекта — стволовой клетки эмбриона мыши, не обладающей таким сложным жизненным циклом, как клетки диктиостелиума или малярийного плазмодия, также гетерогенны, то есть отличаются друг от друга составом входящих в них белков. Получается, что и клетки млекопитающих содержат неодинаковые по составу рибосомы!
Что же дальше?
Пока рано говорить о том, куда приведут нас исследования гетерогенности рибосом. Несомненно, они позволят человечеству лучше понимать механизмы многих заболеваний, в том числе онкологических.
Например, уже выявлены заболевания, именуемые рибосомопатиями, причиной которых являются генетические мутации компонентов рибосом [21].
В заключение
История рибосомы, начавшаяся более шестидесяти лет назад, — яркий пример процесса научного поиска, который привел к изменению нашего взгляда на работу и функции рибосомы. Сейчас мы не можем сказать, чем закончится эта история, к каким результатам и открытиям она приведет.
Углеводные остатки, входящие в структуру клеточной мембраны, выполняют функцию:
1) транспортную
2) сигнальную
3) пиноцитоза
4) фагоцитоза
Плазматическая мембрана
Плазматическая мембрана у эукариот НЕ выполняет функции:
1) транспорта веществ
2) защиты клетки
3) взаимодействия с другими клетками
4) синтеза белка
Цитоплазма
1) раствор минеральных веществ вместе с ядром
2) водный раствор минеральных и органических веществ клетки без ядра
3) внутреннее содержимое ядра
4) раствор органических соединений
- 2) водный раствор минеральных и органических веществ клетки без ядра
Клеточные компоненты
Немембранным компонентом нервной клетки является:
1) рибосома
2) митохондрия
3) ядро
4) эндоплазматическая сеть
Хромосомы
Клетки разных видов могут содержать одинаковое число хромосом, но отличаться:
1) химическим составом хромосом
2) видами нуклеотидов
3) последовательностью нуклеотидов
4) механизмом репликации
Форма клетки
Наиболее изменчивой формой обладает:
1) нервная клетка
2) клетка инфузории туфельки
3) сперматозоид человека
4) лейкоцит человека
Строение клетки
В клетке возбудителя чумы нет:
1) рибосом
2) цитоплазмы
3) мембраны
4) ядра
Клеточная теория
Какое из положений клеточной теории принадлежит Р. Вирхову?
1) все организмы состоят из клеток
2) всякая клетка происходит от другой клетки
3) каждая клетка есть некое самостоятельное целое
4) клетка — элементарная живая система
Собака и мухомор
У собаки и мухомора сходны:
1) способ питания
2) размножение спорами
3) строение клеточной стенки
4) наличие пластид в клетках
Хлоропласты
Хлоропласты есть в клетках:
1) корня капусты
2) гриба-трутовика
3) листа красного перца
4) почек собаки
Общее строение клетки: ядро, цитоплазматическая мембрана, цитоплазма
Клетка — основная функциональная единица организма. Ядро клетки служит хранилищем огромного объёма генетической информации и одновременно центром её активной экспрессии. Существует большое количество различных типов клеток (клетки эпителия, печени, нервных волокон и др.), особенности метаболизма которых обусловлены находящимися в их цитоплазме органеллами, а также множеством растворимых ферментов, характерных для каждого вида клеток.
Цитоплазматическая мембрана, или плазмолемма, — барьер для растворимых в воде молекул, который отделяет внутреннее содержимое клетки от внешней среды. Она состоит из двух параллельных рядов фосфолипидов, которые образуют гидрофобную липидную прослойку между двумя гидрофильными слоями из фосфатных групп.
Плазмолемма пронизана различными белками, гидрофобные части которых находятся внутри билипидного слоя, а гидрофильные — на внешней и внутренней поверхности мембра ны. Микроворсинки — удлинения на верхней (апикальной) части плазмолеммы, которые увеличивают поверхность мембраны и облегчают обмен молекулами.
Ядро клетки. Генетическая информация заключена в хромосомах, которые находятся в ядерном матриксе. Матрикс — сетчатый внутриядерный каркас, состоящий из белкового материала и тесно примыкающий к ядерной оболочке.
Ядрышком называют морфологически выраженную структуру внутри ядра, в которой происходит синтез рибосомальной РНК (рРНК). В ядре клеток человека обычно присутствует одно ядрышко, в котором во время интерфазы возникают ядрышковые организаторы акроцентрических хромосом.
Ядро окружено двойной мембраной, называемой ядерной оболочкой, которая пронизана ядерными порами.
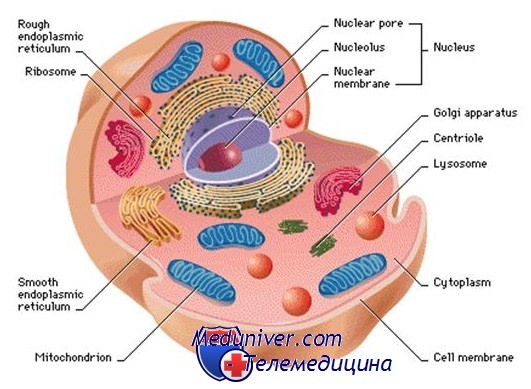
Цитоплазма клетки. Цитоплазма состоит из гелеобразного цитозоля, содержащего запасы гликогена, липидные вкрапления и свободные рибосомы, который пронизан рядами взаимосоединённых волокон и трубочек, образующих цитоскелет. Основные структурные компоненты цитоскелета — микротрубочки, микрофиламенты и промежуточные филаменты.
Микротрубочки — прямые полые цилиндры, стенки которых состоят из чередующихся молекул а- и b-тубулина. Они исходят из клеточного центра (центросомы), который имеет пару центриолей— цилиндрических структур, образованных девятью триплетами микротрубочек. Подобное строение свойственно также базальным тельцам реснитчатого эпителия.
Сеть микротрубочек играет важную роль в поддержании структуры и размера клетки, а также при расхождении хромосом во время деления и движения ресничек и сперматозоидов.
Микрофиламенты представляют собой двуспиральные полимеры белка актина и находятся в основном по периметру клетки. Они участвуют в движении клетки и изменении её формы.
Промежуточные филаменты имеют трубчатую структуру и соединяют десмосомы. В зависимости от вида клетки в их состав входит один или несколько из пяти определённых белков.
Митохондрии — самые крупные и наиболее распространённые в цитоплазме органеллы, основной функцией которых служит обеспечение организма энергией посредством синтеза АТФ. Митохондрии — самовоспроизводящиеся полуавтономные органеллы, содержащие рибосомы и до десяти и более копий кольцевых нитей митохондриальной ДНК.
Данная ДНК кодирует митохондриальные гены. В митохондриях присутствуют ферменты, необходимые для функционирования цикла трикарбоновых кислот, а также большое количество ферментов, участвующих в окислении жирных кислот.
Пероксисомы частично отвечают за детоксикацию различных веществ (в том числе этанола), однако их основная задача — окисление жирных кислот.
Эндоплазматическая сеть (ЭПС) — основной центр синтеза белков и липидов, который также служит начальным этапом секреторного пути белков. ЭПС представляет собой обширный лабиринт из связанных с мембраной каналов, который соединяется непосредственно с ядерной оболочкой.
Вблизи ядра на поверхности ЭПС есть рибосомы (гранулярная ЭПС), в то время как на участках, расположенных дальше, рибосомы отсутствуют (агранулярная или гладкая ЭПС). ЭПС играет важную роль в нейтрализации токсинов. Белки, синтезируемые в ней, затем попадают в комплекс Гольджи — ряд расположенных друг над другом сплюснутых везикул. После этого белки депонируются или попадают в секреторные везикулы для осуществления экзоцитоза, т.е. выведения из клетки в ответ на внешнее воздействие.
Эндоцитоз. Эндоцитозом называют процесс поглощения и переработки клеткой компонентов окружающей среды. При опосредованном рецепторами пиноцитозе происходит захват мелких частиц путём образования везикулы с жидкостью на поверхности цитоплазматической мембраны и её последующего поглощения клеткой. При этом образуются окаймлённые впячивания. Более крупные частицы связываются с мембраной и поглощаются в составе фагоцитарных вакуолей (фаголизосом); растворы поглощаются при помощи жидкостного пиноцитоза.
Содержимое пиноцитарных и фагоцитарных везикул, которые часто называют эндосомами, обычно обрабатывают лизосомы, содержащие разрушающие ферменты — лизоцимы.
Межклеточные соединения. В случае плотного соединения образуется непроницаемая перемычка между внешней (апикальной) и базолатеральной поверхностями эпителиальных клеток. При липких соединениях клетки связаны с помощью опоясывающих (длинные волокна) и точечных (расположены непосредственно в месте скрепления) десмосом. Гемидесмосомы (полудесмо-сомы) соединяют эпителиальные клетки через базальные мембраны (производные экстрацеллюлярного матрикса).
Причина болезни Шарко—Мари—Тута, сцепленной с Х-хромосомой, — дефект белка, участвующего в щелевом соединении клеток.
Большинство лекарственных препаратов вступают во взаимодействие с рецепторами цитоплазматической мембраны. Различные противоопухолевые препараты, такие, как винкристин или винбластин, повреждают систему микротрубочек, в то время как колхицин, применяемый для исследования хромосом, угнетает клетки во время метафа-зы митоза. Клофибрат снижает продукцию дополнительных пероксисом, его используют для снижения уровня липопротеинов в сыворотке крови.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: