
В состав вирусов входит атф
Обновлено: 18.04.2024
Химический состав вирусов. Вирусные геномы. Виды вирусных геномов.
Химический состав вирусов отличается от других форм жизни необычайной простотой. Кроме геномной ДНК или РНК вирусы позвоночных содержат белки, масса которых составляет 57—90% массы вириона. Количество вирионных белков может колебаться в широких пределах в зависимости от сложности строения вируса. Среди белков, кодируемых вирусным геномом, различают структурные и неструктурные вирусспецифические белки. Первые входят в структуру вириона, вторые не входят. Структурными белками являются капсидные белки, белки оболочки и в некоторых случаях белки тегумента и ферменты.
У многих вирусов некоторые структурные вирусные белки не являются первичными продуктами генов, а образуются в результате посттрансляционного расщепления полипептида-предшественника вирусспецифическими и клеточными протеазами.
Неструктурными вирусными белками являются белки, кодируемые вирусным геномом, но не являющиеся составными частями вириона. Одни из них участвуют в репликации вирусных компонентов, другие — в сборке вирионов, третьи нарушают нормальный клеточный синтез. Они включают различные типы репликаз, связанные с репликацией ДНК, транскриптазы, транскрибирующие мРНК на геномной дцДНК или дцРНК или (-)оцРНК, вирусные протеазы, геликазы и лигазы. У ретро- и гепаднавирусов найдена обратная транскриптаза, транскрибирующая ДНК на матрице РНК. У ретровирусов найден уникальный фермент, участвующий в интеграции ДНК-продукта в хромосомальную ДНК клетки. Вирусы оспы, которые размножаются в цитоплазме и имеют много уникальных ферментов для процессинга РНК-транскриптов и репликации вирусной ДНК, мало зависят от метаболизма клетки. Вирусные белки имеют молекулярную массу 5-200 кД. Наиболее просто устроенные вирусы (вирусы-сателлиты, дефектные вирусы) кодируют синтез только одного белка, многие патогенные вирусы кодируют синтез 5—10 белков, крупные вирусы, такие как вирусы оспы, герпесвирусы, кодируют синтез до 200 белков. Хотя это немного по сравнению с клетками прои эукариотов (кодируют соответственно более 5000 и 100000 белков).
Разные вирусы демонстрируют различные варианты стратегии экспрессии своих генов и репликации геномов.
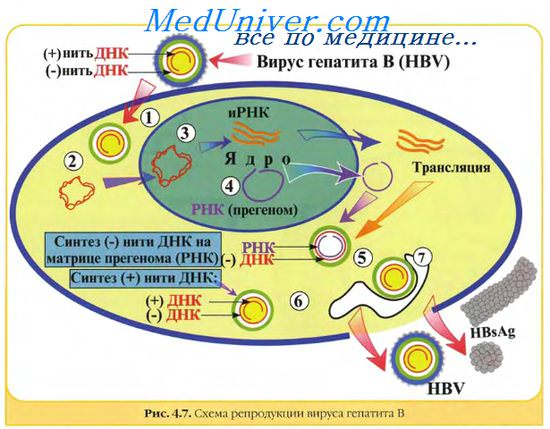
Вирусные геномы. Виды вирусных геномов.
Все вирусные геномы являются гаплоидными, т.е. содержат одну копию каждого гена. Исключение составляют ретровирусы, которые обладают диплоидным геномом. Геномы ДНК-вирусов позвоночных представлены одной двуспиральной молекулой за исключением парво- и цирковирусов.
Геномы полиома-, папиллома-, гепадна- и цирковирусов представлены кольцевой ДНК. ДНК гепаднавирусов частично двуспиральная, частично односпиральная. ДНК вирусов полиомы и папилломы является суперспиральной. Большинство линейных вирусных ДНК обладает способностью приобрести циркулярную конфигурацию, которая требуется для репликации по вращающемуся кольцевому механизму. Две цепи ДНК вируса оспы ковалентно связаны своими концами и при денатурации образуют большое одноцепочечное кольцо. У некоторых ДНК-вирусов (так же как у РНК-ретровирусов) имеются концевые повторяющиеся последовательности. Инвертированные концевые повторы обнаружены у адено- и парвовирусов. У адено-, гепадна- и парвовирусов, так же как у некоторых РНК-вирусов (пикорна- и калицивирусов), с 5'-концом генома ковалентно связан белок, играющий важную роль в его репликации.
Все РНК-вирусы позвоночных за исключением рео- и бирнавирусов имеют одноцепочечные геномы. Геном некоторых РНК-вирусов состоит из нескольких (2-12) уникальных фрагментов, каждый из которых кодирует, как правило, один белок. РНК-вирусы с односпиральным геномом могут иметь различную полярность. Если они имеют ту же полярность, что и мРНК, то они могут прямо индуцировать синтез вирусного белка и считаются положительно (+) полярными.
Если геномная нуклеотидная последовательность комплементарна мРНК, то они считаются отрицательно (—) полярными. К ним относятся: парамиксо-, рабдо-, фило-, ортомиксо-, арена- и буньявирусы. Все они имеют вирионную РНК-зависимую полимеразу (транскриптазу), которая в инфицированной клетке транскрибирует положительно-полярную РНК на матрице геномной вирусной РНК. У аренавирусов, по крайней мере, у одного рода буньявирусов, один из РНК-сегментов является двуполярным. Обычно у (+)полярных РНК-вирусов З'-конец имеет polyA-последовательность, а 5'-конец имеет кэп-структуру.
Размер геномов РНК-вирусов (одноцепочечных 1,7—21 т.н.; двуцепочечных — 18—27 т.п.н.) значительно меньше размера генома многих ДНК-вирусов. Поэтому РНК-вирусы, как правило, кодируют меньше белков, чем ДНК-вирусы. Масса генома различных вирусов находится в пределах от 1 % (орто- и пара-миксовирусы) до 32% (парвовирусы) от массы вириона.
Различные семейства вирусов позвоночных значительно различаются по структуре и функции генома. Основные типы вирусных геномов можно представить следующим образом:
1) двуцепочечной линейной молекулой ДНК с открытыми (герпесвирусы, аденовирусы, иридовирусы) или ковалентно связанными концами (вирусы оспы, асфаровирусы);
2) одноцепочечной линейной молекулой ДНК (парвовирусы);
3) одноцепочечной кольцевой молекулой ДНК (цирковирусы);
4) двуцепочечной кольцевой молекулой ДНК (папилломавирусы, полиомавирусы);
5) частично двуцепочечной кольцевой незамкнутой молекулой ДНК (гепаднавирусы);
6) одноцепочечной молекулой РНК, являющейся мРНК (положительно-геномные вирусы: пикорнавирусы, тогавирусы, флавивирусы, астровирусы, калицивирусы, коронавирусы, артеривирусы, нодавирусы);
7) одноцепочечной единой (рабдовирусы, парамиксовирусы, филовирусы, бор-навирусы) или фрагментированнои (ортомиксовирусы) линейной молекулой РНК, комплементарной мРНК — отрицательно-геномные вирусы;
8) одноцепочечной фрагментированнои кольцевой ковалентно несвязанной отрицательной или двуполярной РНК (буньявирусы, аренавирусы);
9) двуцепочечной линейной фрагментированнои молекулой РНК (реовирусы, бирнавирусы);
10) двумя идентичными линейными молекулами плюс-РНК, являющимися матрицами для синтеза ДНК (ретровирусы).
Молекулярная масса ДНК различных вирусов позвоночных варьирует в широких пределах: от 0,7—1,5 МД у цирковирусов и парвовирусов, до 150—200 МД у вирусов оспы. Молекулярная масса генома у РНК вирусов колеблется менее значительно - от 2,0 до 20,0 МД.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Оглавление
- Введение
- Обозначения и сокращения
- 1 История вирусологии, природа и происхождение вирусов
- 2 Химический состав вирусов
- 3 Морфология, морфогенез, биофизические свойства и генетика вирусов
Приведённый ознакомительный фрагмент книги Общая вирусология с основами таксономии вирусов позвоночных предоставлен нашим книжным партнёром — компанией ЛитРес.
2 Химический состав вирусов
Основными химическими соединениями, которые входят в состав всех вирусов, являются белки и нуклеиновые кислоты. В состав ряда вирусов входят липиды и углеводы.
Белки. Локализация вирусных белков. Белки, связанные с жизненным циклом вируса, разделяют на белки, детерминируемые геномом вируса и белки, имеющие клеточное происхождение. В качестве примера клеточных белков, которые обнаружены в составе некоторых вирионов, могут быть приведены белок цитоскелета — актин, и ядерные белки — гистоны. Белки клеточного происхождения, участвующие в процессе репликации вируса, будут рассмотрены в разделе взаимодействия вируса с клеткой.
По месту локализации белки, детерминируемые вирусным геномом, разделяют на две группы: 1) структурные белки — это белки, входящие в состав ВЧ, их обозначают как VP; 2) неструктурные белки — это предшественники структурных белков, регуляторные белки и ферменты, обслуживающие процесс внутриклеточной репродукции вируса и не входящие в состав ВЧ. Их обозначают как NS-белки.
Свойства вирусных белков. В состав вирионов входят белки с различной молекулярной массой (от 4 до 100 кД), состоящие из одной или нескольких полипептидных цепей. Количество этих белков также различно у разных вирусов. В состав нуклеокапсида ВТМ входит один белок. У других вирусов в состав вириона может входить несколько десятков белков, имеющих различные физико-химические свойства. Белки, формирующие капсид, нуклеокапсид и коровую оболочку, обладают одним общим свойством — способностью к самосборке.
В состав ВЧ могут входить низкомолекулярные белки, не участвующие в формировании капсида. Например, геномные белки пикорнавирусов и аденовирусов. Геномный белок ковалентно связан с нуклеиновой кислотой и участвует в ее репликации.
Сложные белки представлены гликопротеинами (обозначают как gp) и липопротеинами. Наличие гликопротеина определяет присутствие в вирионе углеводного компонента, который может быть представлен олигосахаридами маннозного типа, галактозой, N-ацетилглюкозамином или нейраминовой кислотой. Вирусные гликопротеины, как правило, экспонированы на наружной поверхности ВЧ и выполняют три основные функции: обеспечивают связывание вириона с клеточным рецептором (функция прикрепительного белка), обладают фузионной активностью (обеспечивают слияние мембран) и определяют антигенные свойства вирусов. В то же время, вирусные гликопротеины могут быть и неструктурными белками и, оставаясь в интегральной форме в мембране шероховатого эндоплазматического ретикулюма (ШЭР), выполнять функции транслоказ, обеспечивая транспорт вирусных компонентов в его просвет.
Вирусные липопротеины представлены белками, ацилированными, как правило, миристиновой (C14) кислотой. Остатки жирных кислот, соединенные с молекулой белка, выполняют функцию липофильного якоря.
Вирусные белки-ферменты могут входить в состав вирусной частицы или являться неструктурными белками и появляться в клетке после экспрессии вирусного генома. Наиболее оснащенным ферментами является вирион вируса оспы, который имеет практически полный набор энзимов, необходимых для независимой внутриклеточной репликации вируса. В то же время, мелкие просто организованные изометрические вирусы с позитивным РНК-геномом могут не иметь никаких ферментов в составе вириона.
Функционально активные белки вирусов представлены, в первую очередь, ферментами нуклеинового обмена, обеспечивающими сложные механизмы репликации/транскрипции вирусного генома; ферментами, осуществляющими посттрансляционный процессинг и модификацию белков, и ферментами, участвующими в проникновении вирионов в клетку хозяина.
Первая группа ферментов наиболее многочисленна и включает как аналоги клеточных ферментов, так и вирус-специфические ферменты.
ДНК-зависимая ДНК-полимераза — осуществляет синтез ДНК на матрице ДНК (вирус оспы).
ДНК-зависимая РНК-полимераза — осуществляет синтез мРНК на матрице ДНК (вирус оспы).
Обратная транскриптаза или ревертаза или РНК-зависимая ДНК-полимераза осуществляет синтез ДНК на матрице РНК. Впервые открыта в 1970 г. у ретровирусов Темином и Мизутани.
Хеликаза — осуществляет расплетете двухнитевой структуры ДНК. Кроме этого хеликазы обладают нуклеотидтрифосфат-зависимой РНК-хеликазной активностью, которая включает три процесса: связывание дезоксинуклеотидтрифосфата, его гидролиз и за счет этой энергии расплетение двухнитевой РНК.
мРНК-модифицирующие ферменты : поли-А-полимераза — аденилирует 3'конец РНК за счет энергии АТФ; Кэп-энзим и метилтрансферазный комплекс — катализирует образование на 5'-конце кэп-структуры.
АТФ-аза, ГТФ-аза — осуществляют гидролиз соответствующих энергетических субстратов.
Рибонуклеаза Н — разрушает РНК, находящуюся в дуплексе с ДНК. Вторая группа вирусных ферментов — ферменты белкового обмена.
Здесь мы приведем лишь некоторые из них:
Протеиназы — ферменты, участвующие в посттрансляционном процессинге полипротеинов. Являются NS-белками РНК-содержащих вирусов;
Протеинкиназы — ферменты, фосфорилирующие структурные белки вирионов. Обнаружены в составе вируса везикулярного стоматита, вируса бешенства, альфавирусов и ретровирусов. Примерами ферментов, участвующих в проникновении вирусов в клетку, являются лизоцим бактериофагов и нейраминидаза вируса гриппа.
Липиды. Все оболочечные РНК-содержащие почкующиеся вирусы имеют липиды клеточного происхождения, входящие в состав суперкапсида (от 15 % до 30 % от сухого веса). От 50 % до 60 % липидов представлены фосфолипидами, от 20 % до 30 % составляет холестерин.
У ДНК-геномных вирусов липиды содержат вирусы оспы, герпеса, гепатита B. Это непочкующиеся вирусы. У вируса оспы липиды не образуют дифференцированной оболочки, которая формируется в цитоплазме в процессе морфогенеза поксвириона. Липиды вируса гепатита B образуются путем инвагинации мембран эндоплазматического ретикулюма (ЭПР). Липидсодержащая оболочка вируса герпеса формируется при прохождении внутреннего компонента вириона через ядерную мембрану. Следовательно, в состав вирусной оболочки герпесвирусов входят липиды ядерной мембраны.
Нуклеиновые кислоты. Клетки всех живых организмов содержат два вида нуклеиновой кислоты — ДНК (двухнитевая ДНК клеточного генома) и РНК (мРНК, тРНК, рРНК). В отличие от клеток, вирионы содержат только один вид нуклеиновой кислоты — ДНК или РНК. И та и другая являются хранителями наследственной информации и выполняют функции генома. Однако следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса. В жизненном цикле вируса его геномная нуклеиновая кислота транскрибируется, то есть ДНК-содержащие вирусы образуют РНК. Ряд РНК-содержащих вирусов имеют в цикле репродукции стадию обратной транскрипции и синтезируют ДНК на матрице РНК. Примерно 20 % всех вирусов имеют ДНК-геном, 80 % — РНК-геном. Способность РНК хранить наследственную информацию — уникальное свойство вирусов. Размеры вирусных геномов (длина нуклеотидных последовательностей, выраженная в нуклеотидах) варьируют в широких пределах — от 1,7 тысяч нуклеотидов (т.н.) у цирковируса свиней до 300 т.н. у фикоднавирусов архибактерий.
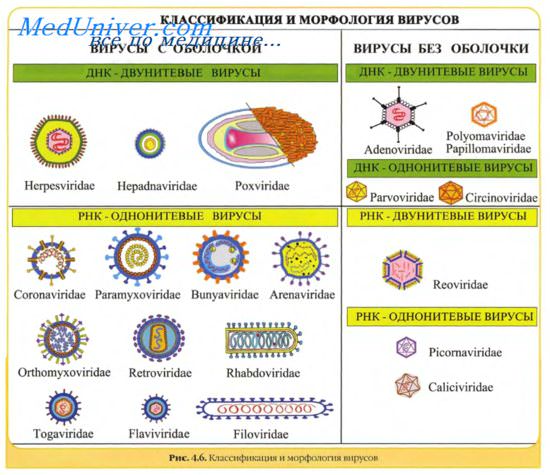
Кроме того, что геном вирусов может быть представлен или ДНК или РНК, он может находиться в разных видах — в виде двухнитевой (дн) или однонитевой (он) формы, в виде линейной или кольцевой, в виде непрерывной или сегментированной формы.
Многообразие видов РНК геномов расширяется за счет существования последовательностей, отличающихся направлением связей сахаро-фосфатного остова.
Однонитевые РНК могут иметь позитивную полярность — (+) РНК, негативную полярность — (-) РНК или могут быть представлены обоюдозначащей цепью — (+, — ) РНК (амбисенс стратегия кодирования). В свою очередь, РНК позитивной полярности могут иметь разную структурную организацию: могут, являясь матричной РНК, иметь на 5'-конце кэп (7-метилгуанозин, Сар), а на 3'-конце — поли-А (poly-A) последовательность; могут не иметь кэпа или поли-А; могут иметь на 5'-конце геномный белок; могут иметь на 3'-конце тРНК-подобную или шпильковую структуру.
Виды геномов вирусов легли в основу их классификации. Однако следует учитывать, что вид генома в настоящее время не является формальным таксоном и используется для удобства ориентации в многообразии вирусов.
Углеводы. Углеводный компонент вирусов находится в составе гликопротеидов. Количество сахаров в составе гликопротеидов может быть достаточно большим, достигая от 10 % до 13 % от массы вириона. Химическая специфичность их полностью определяется клеточными ферментами, обеспечивающими перенос и присоединение соответствующих сахарных остатков. Обычными сахарными остатками, обнаруживаемыми в вирусных белках, являются фруктоза, сахароза, манноза, галактоза, нейраминовая кислота, глюкозамин. Таким образом, подобно липидам, углеводный компонент определяется клеткой-хозяином, благодаря чему один и тот же вирус, выращенный в клетках разных видов, может значительно отличаться по составу сахаров в зависимости от специфичности клеточных гликозилтрансфераз.
Углеводный компонент гликопротеидов играет существенную роль в структуре и функции белка. Он является каркасом для локальных участков гликопротеида, обеспечивая сохранение конформации белковой молекулы, и обусловливает защиту молекулы от протеаз. Возможны и другие функции углеводов, пока достоверно не установленные.
Компоненты клетки-хозяина. В составе вирионов могут находиться компоненты клетки-хозяина. К таким компонентам могут относиться белки, и даже целые клеточные структуры. Так, например, в составе ряда оболочечных вирусов может находиться белок цитоскелета актин, в составе паповавирусов содержатся клеточные гистоны. Ряд вирусов содержит клеточные ферменты, например, протеинкиназы. В составе аренавирусов обнаружены рибосомы.
Клеточные компоненты могут включаться в вирион случайно или закономерно, В некоторых случаях они играют существенную роль в репродукции вируса, как, например, гистоны в репродукции паповавирусов.
Строение вирусов. Классификация вирусов
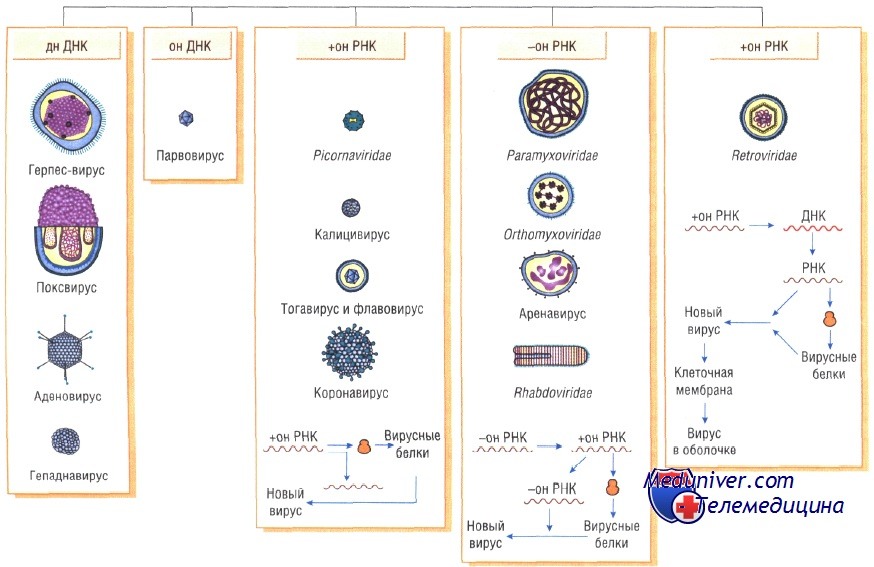
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
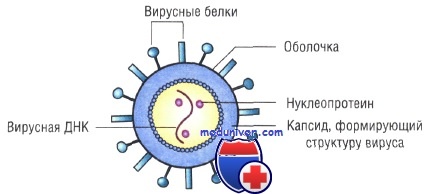
Структура и состав вирусов. Строение вирусов и вирусных частиц.
Основным структурным компонентом вирионов (полных вирусных частиц) является нуклеокапсид, т.е. белковый чехол (капсид) в котором заключен вирусный геном (ДНК или РНК). Нуклеокапсид большинства семейств вирусов окружен липопротеиновой оболочкой. Между оболочкой и нуклеокапсидом у некоторых вирусов (орто-, парамиксо-, рабдо-, фило- и ретровирусов) находится негликозилированный матриксный белок, придающий дополнительную жесткость вирионам. Вирусы большинства семейств имеют оболочку, которая играет важную роль в инфекционности. Наружный слой оболочки вирионы приобретают, когда нуклеокапсид проникает через клеточную мембрану почкованием.
Белки оболочки кодируются вирусом, а липиды заимствуются из мембраны клетки. Гликопротеины обычно в виде димеров и тримеров образуют пепломеры (выступы) на поверхности вирионов (орто-, парамиксовирусы, рабдо-, фило-, корона-, бунья-, арена-, ретровирусы). Гликозилированные белки слияния связаны с пепломерами и выполняют ключевую роль в проникновении вируса в клетку. Капсиды и оболочки вирионов образуются множеством копий одного или нескольких видов белковых субъединиц в результате процесса самосборки. Взаимодействие в системе белок-белок, благодаря слабым химическим связям, ведет к объединению симметричных капсидов.
Различия вирусов по форме и размеру вирионов зависят от формы, размера и количества структурных белковых субъединиц и природы взаимодействия между ними.
Капсид состоит из множества морфологически выраженных субъединиц (капсомеров), собранных из вирусных полипептидов строго определенным образом, в соответствии с относительно простыми геометрическими принципами. Белковые субъединицы, соединяясь друг с другом, образуют капсиды двух видов симметрии: изометрические и спиральные. Структура нуклеокапсида оболочечных вирусов сходна со структурой нуклеокапсида безоболочечных вирусов. На поверхности оболочки вирусов различают морфологически выраженные гликопротеиновые структуры — пепломеры.
В состав суперкапсидной оболочки входят липиды (до 20—35%) и углеводы (до 7—8%), имеющие клеточное происхождение. Она состоит из двойного слоя клеточных липидов и вирусспецифических белков, расположенных снаружи и изнутри липидного биослоя. Наружный слой суперкапсидной оболочки представляют пепломеры (выступы) одного или более типов, состоящие из одной или нескольких молекул гликопротеинов. Нуклеокапсид у оболочечных вирусов часто называют сердцевиной (core), а центральную часть вирионов, содержащую нуклеиновую кислоту, называют нуклеоидом.
Капсомеры (пепломеры) состоят из структурных единиц, построенных из одной либо из нескольких гомологичных или гетерологичных полипептидных цепей (белковых субъединиц).
Изометрические капсиды представляют собой не сферы, а правильные многогранники (икосаэдры). Их линейные размеры идентичны по осям симметрии. Согласно Каспару и Клугу (1962), капсомеры в капсидах расположены в соответствии с икосаэдрической симметрией.
Такие капсиды состоят из идентичных субъединиц, образующих икосаэдр. Они имеют 12 вершин (углов), 30 граней и 20 поверхностей в виде равнобедренных треугольников. В соответствии с этим правилом капсид полиовируса и вируса ящура образован 60 белковыми структурными единицами, каждая из которых состоит из четырех полипептидных цепей.
Икосаэдр оптимально решает проблему укладки повторяющихся субъединиц в строгую компактную структуру при минимальном объеме. Только некоторые конфигурации структурных субъединиц могут сформировать поверхности, образовать вершины и грани вирусного икосаэдра. Например, структурные субъединицы аденовируса на поверхностях и гранях формируют шестигранные капсомеры (гексоны), а на вершинах - пятигранные капсомеры (пептоны). У одних вирусов оба вида капсомеров образуются одними и теми же полипептидами, у других — разными полипептидами. Так как структурные субъединицы разных вирусов различаются между собой, то одни вирусы кажутся более гексагональными, другие — более сферическими.
Все известные ДНК-содержащие вирусы позвоночных, за исключением вирусов оспы, а также многие РНК-содержащие вирусы (7 семейств) имеют кубический тип симметрии капсида.
Реовирусы, в отличие от других вирусов позвоночных, имеют двойной кап-сид (наружный и внутренний), причем каждый состоит из морфологических единиц.
Вирусы, обладающие спиральным типом симметрии, имеют вид цилиндрической нитевидной структуры, их геномная РНК имеет вид спирали и находится внутри капсида. Все вирусы животных спиральной симметрии окружены липопротеиновой оболочкой.
Спиральные нуклеокапсиды характеризуются длиной, диаметром, шагом спирали и числом капсомеров, приходящихся на один оборот спирали. Так, у вируса Сендай (парамиксовирус) нуклеокапсид представляет собой спираль длиной около 1 мкм, диаметром 20 нм и шагом спирали 5 нм. Капсид состоит примерно из 2400 структурных единиц, каждая из которых является белком с молекулярной массой 60 кД. На каждый виток спирали приходится 11—13 субъединиц.
У вирусов со спиральным типом симметрии нуклеокапсида укладка белковых молекул в спираль обеспечивает максимальное взаимодействие между нуклеиновой кислотой и белковыми субъединицами. У икосаэдрических вирусов нуклеиновая кислота находится внутри вирионов в скрученном состоянии и взаимодействует с одним или несколькими полипептидами, расположенными внутри капсида.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: