
В составе вирусов имеются нуклеиновые кислоты сколько видов
Обновлено: 13.05.2024
Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных.
Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
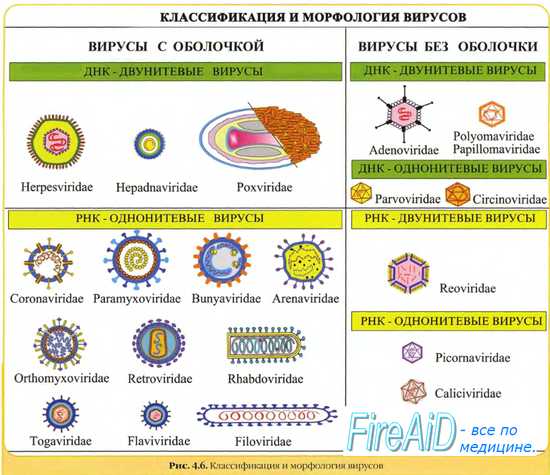
Морфология вирусов. Размеры вирусов.
Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки.
Нуклеиновые кислоты вирусов
Вирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине.
Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180") повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
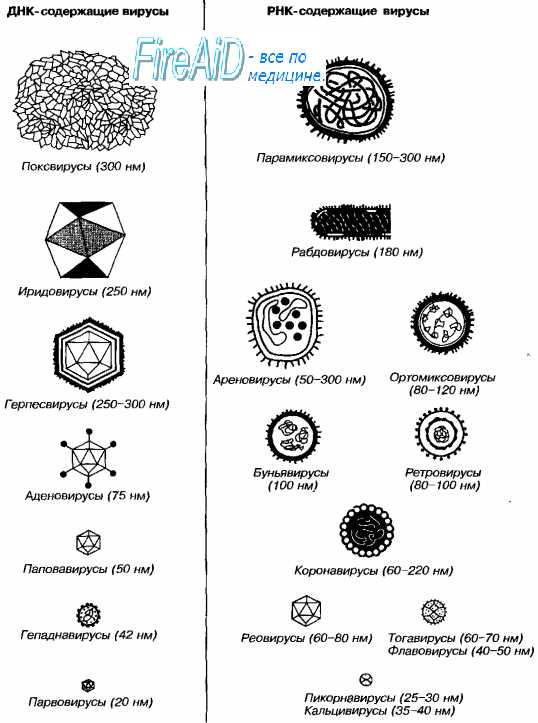
Рис. 2-1. Размеры и морфология основных возбудителей вирусных инфекций человека.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК.
Инфекционность нуклеиновых кислот вирусов
Многие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Химический состав вирусов. Вирусные геномы. Виды вирусных геномов.
Химический состав вирусов отличается от других форм жизни необычайной простотой. Кроме геномной ДНК или РНК вирусы позвоночных содержат белки, масса которых составляет 57—90% массы вириона. Количество вирионных белков может колебаться в широких пределах в зависимости от сложности строения вируса. Среди белков, кодируемых вирусным геномом, различают структурные и неструктурные вирусспецифические белки. Первые входят в структуру вириона, вторые не входят. Структурными белками являются капсидные белки, белки оболочки и в некоторых случаях белки тегумента и ферменты.
У многих вирусов некоторые структурные вирусные белки не являются первичными продуктами генов, а образуются в результате посттрансляционного расщепления полипептида-предшественника вирусспецифическими и клеточными протеазами.
Неструктурными вирусными белками являются белки, кодируемые вирусным геномом, но не являющиеся составными частями вириона. Одни из них участвуют в репликации вирусных компонентов, другие — в сборке вирионов, третьи нарушают нормальный клеточный синтез. Они включают различные типы репликаз, связанные с репликацией ДНК, транскриптазы, транскрибирующие мРНК на геномной дцДНК или дцРНК или (-)оцРНК, вирусные протеазы, геликазы и лигазы. У ретро- и гепаднавирусов найдена обратная транскриптаза, транскрибирующая ДНК на матрице РНК. У ретровирусов найден уникальный фермент, участвующий в интеграции ДНК-продукта в хромосомальную ДНК клетки. Вирусы оспы, которые размножаются в цитоплазме и имеют много уникальных ферментов для процессинга РНК-транскриптов и репликации вирусной ДНК, мало зависят от метаболизма клетки. Вирусные белки имеют молекулярную массу 5-200 кД. Наиболее просто устроенные вирусы (вирусы-сателлиты, дефектные вирусы) кодируют синтез только одного белка, многие патогенные вирусы кодируют синтез 5—10 белков, крупные вирусы, такие как вирусы оспы, герпесвирусы, кодируют синтез до 200 белков. Хотя это немного по сравнению с клетками прои эукариотов (кодируют соответственно более 5000 и 100000 белков).
Разные вирусы демонстрируют различные варианты стратегии экспрессии своих генов и репликации геномов.
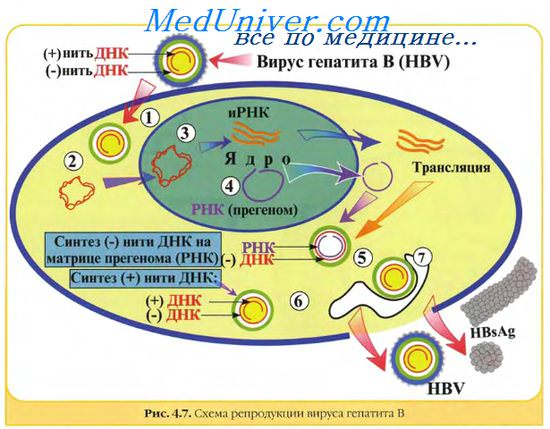
Вирусные геномы. Виды вирусных геномов.
Все вирусные геномы являются гаплоидными, т.е. содержат одну копию каждого гена. Исключение составляют ретровирусы, которые обладают диплоидным геномом. Геномы ДНК-вирусов позвоночных представлены одной двуспиральной молекулой за исключением парво- и цирковирусов.
Геномы полиома-, папиллома-, гепадна- и цирковирусов представлены кольцевой ДНК. ДНК гепаднавирусов частично двуспиральная, частично односпиральная. ДНК вирусов полиомы и папилломы является суперспиральной. Большинство линейных вирусных ДНК обладает способностью приобрести циркулярную конфигурацию, которая требуется для репликации по вращающемуся кольцевому механизму. Две цепи ДНК вируса оспы ковалентно связаны своими концами и при денатурации образуют большое одноцепочечное кольцо. У некоторых ДНК-вирусов (так же как у РНК-ретровирусов) имеются концевые повторяющиеся последовательности. Инвертированные концевые повторы обнаружены у адено- и парвовирусов. У адено-, гепадна- и парвовирусов, так же как у некоторых РНК-вирусов (пикорна- и калицивирусов), с 5'-концом генома ковалентно связан белок, играющий важную роль в его репликации.
Все РНК-вирусы позвоночных за исключением рео- и бирнавирусов имеют одноцепочечные геномы. Геном некоторых РНК-вирусов состоит из нескольких (2-12) уникальных фрагментов, каждый из которых кодирует, как правило, один белок. РНК-вирусы с односпиральным геномом могут иметь различную полярность. Если они имеют ту же полярность, что и мРНК, то они могут прямо индуцировать синтез вирусного белка и считаются положительно (+) полярными.
Если геномная нуклеотидная последовательность комплементарна мРНК, то они считаются отрицательно (—) полярными. К ним относятся: парамиксо-, рабдо-, фило-, ортомиксо-, арена- и буньявирусы. Все они имеют вирионную РНК-зависимую полимеразу (транскриптазу), которая в инфицированной клетке транскрибирует положительно-полярную РНК на матрице геномной вирусной РНК. У аренавирусов, по крайней мере, у одного рода буньявирусов, один из РНК-сегментов является двуполярным. Обычно у (+)полярных РНК-вирусов З'-конец имеет polyA-последовательность, а 5'-конец имеет кэп-структуру.
Размер геномов РНК-вирусов (одноцепочечных 1,7—21 т.н.; двуцепочечных — 18—27 т.п.н.) значительно меньше размера генома многих ДНК-вирусов. Поэтому РНК-вирусы, как правило, кодируют меньше белков, чем ДНК-вирусы. Масса генома различных вирусов находится в пределах от 1 % (орто- и пара-миксовирусы) до 32% (парвовирусы) от массы вириона.
Различные семейства вирусов позвоночных значительно различаются по структуре и функции генома. Основные типы вирусных геномов можно представить следующим образом:
1) двуцепочечной линейной молекулой ДНК с открытыми (герпесвирусы, аденовирусы, иридовирусы) или ковалентно связанными концами (вирусы оспы, асфаровирусы);
2) одноцепочечной линейной молекулой ДНК (парвовирусы);
3) одноцепочечной кольцевой молекулой ДНК (цирковирусы);
4) двуцепочечной кольцевой молекулой ДНК (папилломавирусы, полиомавирусы);
5) частично двуцепочечной кольцевой незамкнутой молекулой ДНК (гепаднавирусы);
6) одноцепочечной молекулой РНК, являющейся мРНК (положительно-геномные вирусы: пикорнавирусы, тогавирусы, флавивирусы, астровирусы, калицивирусы, коронавирусы, артеривирусы, нодавирусы);
7) одноцепочечной единой (рабдовирусы, парамиксовирусы, филовирусы, бор-навирусы) или фрагментированнои (ортомиксовирусы) линейной молекулой РНК, комплементарной мРНК — отрицательно-геномные вирусы;
8) одноцепочечной фрагментированнои кольцевой ковалентно несвязанной отрицательной или двуполярной РНК (буньявирусы, аренавирусы);
9) двуцепочечной линейной фрагментированнои молекулой РНК (реовирусы, бирнавирусы);
10) двумя идентичными линейными молекулами плюс-РНК, являющимися матрицами для синтеза ДНК (ретровирусы).
Молекулярная масса ДНК различных вирусов позвоночных варьирует в широких пределах: от 0,7—1,5 МД у цирковирусов и парвовирусов, до 150—200 МД у вирусов оспы. Молекулярная масса генома у РНК вирусов колеблется менее значительно - от 2,0 до 20,0 МД.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
НУКЛЕИНОВЫЕ КИСЛОТЫ — класс биополимеров, ответственных за хранение, передачу и воплощение генетической информации; универсальные компоненты всех живых организмов.
Имеется два типа Н. к.: дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК), углеводные компоненты к-рых представлены дезоксирибозой (см.) и рибозой (см.) соответственно. Биол, роль этих типов Н. к. заключается в том, что у большинства живых организмов ДНК несет функцию хранения и воспроизведения наследственной информации, в то время как РНК отвечает за воплощение этой информации в процессе синтеза белков (см. Дезоксирибонуклеиновые кислоты, Рибонуклеиновые кислоты).
Различают следующие основные виды РНК: информационная, или матричная (иРНК, или мРНК), к-рая служит матрицей для синтеза белков; рибосомная (рРНК), являющаяся компонентом белоксинтезирующего аппарата клетки — рибосомы (см.); транспортная (тРНК), функция к-рой состоит в переносе активированных аминокислот в место синтеза белка — рибосому. РНК служит генетическим материалом многих вирусов.
Основная часть ДНК эукариотов содержится в хромосомах. Кроме того, ДНК присутствует в хлоропластах растений и митохондриях животных и растений. В нек-рых клетках, напр, спермиях, содержание ДНК может составлять около половины веса всей клетки. Удобным источником для получения ДНК являются лимфоциты вилочковой железы. РНК преимущественно находится в цитоплазме. Содержание РНК в клетках сильно варьирует в зависимости от интенсивности белкового синтеза в них. Много РНК содержится, напр., в поджелудочной железе, а также в быстрорастущих эмбриональных и опухолевых клетках. Высокоспециализированные мышечная и нервная ткани содержат относительно мало РНК.
Н. к. были открыты в 1868 г. швейцарским химиком Мише ром (F. Miescher), к-рый показал, что они локализованы в ядрах клетки, обладают кислотными свойствами и в отличие от белков содержат фосфор.
Химически Н. к. представляют собой полинуклеотиды, состоящие из мономерных звеньев — так наз. мононуклеотидов (нуклеотидов). Каждый нуклеотид содержит одно из четырех типов азотистых оснований: пурины — аденин (А) и гуанин (Г) и пиримидины — цитозин (Ц) и тимин (Т). В нуклеотидах РНК вместо тимина присутствует урацил (У). Основание, соединенное с углеводом — пентозой, образует так наз. нуклеозид, фосфорилированные производное к-рого называется нуклеотидом. Нуклеотиды в Н. к. соединены посредством фосфодиэфирной связи, где R — H (водород) для ДНК и — OH (гидроксил) для РНК.
Специфичность Н. к. определяется порядком чередования азотистых оснований, что определяет так наз. первичную структуру Н. к. Первичная структура установлена для многих тРНК, рРНК, иРНК, а также ряда РНК- и ДНК-содержащих вирусов и бактериофагов. Пространственная структура Н. к. определяется нековалентными взаимодействиями: водородными связями между основаниями, гидрофобными взаимодействиями между плоскостями пар оснований, электростатическими взаимодействиями с участием отрицательно заряженных фосфатных групп и противоионов.
Важнейшим достижением молекулярной биологии (см.) было открытие в 1953 г. Дж. Уотсоном и Ф. Криком двойной спирали ДНК, в молекуле к-рой две антипараллельно расположенные сахарофосфатные цепи удерживаются водородными связями между аденином и тимином или гуанином и цитозином. Последовательность азотистых оснований в одной цепи определяет последовательность оснований в другой. Размеры комплементарных пар А — T и Г — Ц одинаковы. Это позволяет нуклеотидной цепи, состоящей из комплементарных пар оснований в произвольной последовательности, свернуться в правильную двойную спираль (вторичная структура Н. к.). В физиол, условиях двойная спираль ДНК близка к так наз. B-форме, в к-рой пары оснований, лежащие одна над другой (стопкой), перпендикулярны оси спирали.
На один виток спирали приходится 10 пар оснований. В солевом растворе Н. к. число пар азотистых оснований, приходящихся на один виток спирали, варьирует, уменьшаясь при увеличении концентрации соли в р-ре и понижении температуры.
Синтез Н. к. в клетке осуществляется по принципу копирования молекулы-матрицы белками-полимеразами, при этом происходит реакция поликонденсации нуклеозидтрифосфа-тов с отщеплением пирофосфата. Последовательность азотистых оснований в молекуле продукта этого ферментативного процесса определяется последовательностью азотистых оснований в молекуле-матрице. Синтез ДНК называется репликацией (см.) и осуществляется комплексом белков, состоящим из ДНК-полимеразы, белка, разделяющего нити ДНК, нуклеазы, лигазы и т. д. Синтез РНК — транскрипция (см.) происходит по матрице ДНК, причем РНК-полимеразы считывают одну, так наз. значащую, нить двойной спирали. В процессе транскрипции образуется РНК-копия гена (см.). В последовательности оснований мРНК заключена информация для синтеза специфических белков. Каждой аминокислоте белка соответствуют определенные тройки нуклеотидов — триплеты, образующие генетический код (см.). Поэтому изменения нуклеотидной последовательности в ДНК — мутации (см.) сказываются на структуре синтезируемых белков, влияют на их функцию и на наследственную информацию. Искажение наследственной информации может быть причиной наследственных болезней (см.), напр, серповидно-клеточной анемии, фенилкетонурии, или наследственных аномалий, напр, альбинизма. Мутации могут подхватываться естественным отбором и служат фактором эволюции. Естественная частота мутирования очень низка, но она значительно повышается при воздействии ионизирующего, ультрафиолетового излучений или хим. мутагенов. В ходе эволюции живые организмы выработали эффективные механизмы репарации (восстановления) Н. к., устраняющие повреждения в их молекулах (см. Репарация генетических повреждений).
В соответствии с хим. строением полинуклеотидной цепи существуют три группы методов количественного определения Н. к.: по содержанию азотистых оснований [обычно пользуются определением величины поглощения в ультрафиолетовой части спектра, т. е. спектрофотометрией (см.)], по содержанию углеводного компонента [всевозможные цветные реакции (см. Углеводы)], по количеству фосфора. Спектрофотометрические методы дают удовлетворительные результаты только при незначительном количестве примесей в исследуемом образце. Методы второй группы специфичны к типу Н. к. и позволяют отличать ДНК от РНК.
Библиография: Органическая химия нуклеиновых кислот, под ред. Н. К. Кочеткова и Э. И. Будовского, М., 1970; Химия и биохимия нуклеиновых кислот, под ред. И. Б. Збарского и С. С. Дебова, JI., 1968; Ш а б а р о в а 3. А. и Богданов А. А. Химия нуклеиновых кислот и их компонентов, М., 1978; Chromatin, Cold Spr. Harb. Symp. quant. Biol., v. 42, pt 2, 1978; Handbook of biochemistry and molecular biology, nucleic acids, ed. by G. D. Fasman, v. 1, Cleveland, 1975.
Читайте также: