
Вирус гриппа чувствителен к
Обновлено: 24.04.2024
Этиология гриппа
Грипп — острое респираторное вирусное заболевание, этиологически связанное с представителями трех родов — Influenza A virus (вирусы гриппа А), Influenza В virus (вирусы гриппа В) и Influenza С virus (вирусы гриппа С) — из семейства Orthomyxoviridae [2, 26].
Вирус гриппа А был впервые изолирован от свиней американским вирусологом Ричардом Шоупом (1901–1966) в 1930 г.; от людей — тремя годами позже группой английских ученых: Вильсоном Смитом (1897–1965), Кристофером Эндрюсом (1896–1987) и Патриком Лейдлоу (1881–1940) [26].
| Рис. 1. Структура вириона вируса гриппа А (Orthomyxoviridae, Influenza A virus) |
На поверхности вириона (вирусной частицы) вируса гриппа А имеются две функционально-важные молекулы (рис. 1): гемагглютинин (с помощью которого вирион прикрепляется к поверхности клетки-мишени); нейраминидаза (разрушающая клеточный рецептор, что необходимо при почковании дочерних вирионов, а также для исправления ошибок при неправильном связывании с рецептором) [2, 24, 26].
В настоящее время известны 16 типов гемагглютинина (обозначаемые как Н1, Н2, …, Н16) и 9 типов нейраминидазы (N1, N2, …, N9). Комбинация типа гемагглютинина и нейраминидазы (например, H1N1, H3N2, H5N1 и т. п.) называется субтипом: из 144 (16 × 9) теоретически возможных субтипов на сегодняшний день известны 115 [24].
Природным резервуаром вируса гриппа А являются дикие птицы водно-околоводного экологического комплекса (в первую очередь, речные утки, чайки и крачки), однако вирус способен преодолевать межвидовой барьер, адаптироваться к новым хозяевам и длительное время циркулировать в их популяциях [9–12]. Эпидемические варианты вируса гриппа А вызывают ежегодный подъем заболеваемости и раз в 10–50 лет — опасные пандемии [1, 11, 16].
Вирус гриппа В был открыт в 1940 г. американским вирусологом Томасом Фрэнсисом-младшим (1900–1969). Вирус гриппа В не вызывает пандемии, но является возбудителем крупных эпидемических вспышек [26].
Вирус гриппа С был открыт в 1947 г. американским вирусологом Ричардом Тейлором (1887–1981). Вирус гриппа С вызывает локальные эпидемические вспышки в детских коллективах. Наиболее тяжело инфекция протекает у детей младшего возраста [26].
Вирусы гриппа занимают важное место в структуре заболеваемости людей острыми респираторными вирусными инфекциями (ОРВИ), составляющими до 90% от всех других инфекционных болезней. По данным Всемирной Организации Здравоохранения (ВОЗ), только тяжелыми формами гриппа в мире ежегодно заболевают 3–5 млн человек. Заболевает ежегодно гриппом и другими ОРВИ в РФ — 25–35 млн, из них 45–60% — дети. Экономический ущерб РФ от сезонного эпидемического гриппа составляет до 100 млрд руб./год, или порядка 85% экономических потерь от инфекционных болезней [2–8, 20–23].
История гриппозных пандемий
В первый постпандемический эпидсезон 2010–2011 гг. пандемический грипп А(H1N1) swl стал причиной более 70% случаев ОРВИ в мире, грипп А(H3N2) — 1–5%, грипп В — 10–20%.
Состав противогриппозных вакцин в эпидсезоне 2011–2012 гг. (как и в 2010–2011 гг.): A/California/07/2009 (H1N1) swl; A/Perth/16/2009 (H3N2); B/Brisbane/60/2008.
Пандемический вирус гриппа А(H1N1) swl резистентен к Ремантадину и Амантадину, но чувствителен к Тамифлю, Релензе, Ингавирину, Арбидолу и Рибавирину [13–16, 18, 19, 23].
Высоковирулентный грипп А(H5N1) птиц — возможный возбудитель очередной пандемии.
Вероятность преодоления вирусом гриппа А межвидового барьера и проникновения в человеческую популяцию с опасными последствиями резко увеличивается в период эпизоотий 3 . Поэтому высоковирулентный вирус гриппа А(H5N1) птиц, ставший причиной современной масштабной эпизоотии среди диких и домашних птиц Старого Света и имеющий повышенную способность репродуцироваться в клетках млекопитающих, рассматривается как наиболее вероятный возбудитель очередной пандемии гриппа [10, 11, 17]. Дальнейшее распространение этого вируса может иметь катастрофические последствия в случае появления у него эпидемического потенциала (способности передаваться от человека к человеку), так как, во-первых, у человечества отсутствует коллективный иммунитет к вирусам гриппа А (Н5), а во-вторых, из 563 лабораторно подтвержденных случаев заболевания людей в 15 странах мира в результате заражения вирусом гриппа А(H5N1) птичьего происхождения за 2003–2011 гг. 330 умерли, т. е. летальность приближается к 60% [11, 24].
Патогенез гриппа
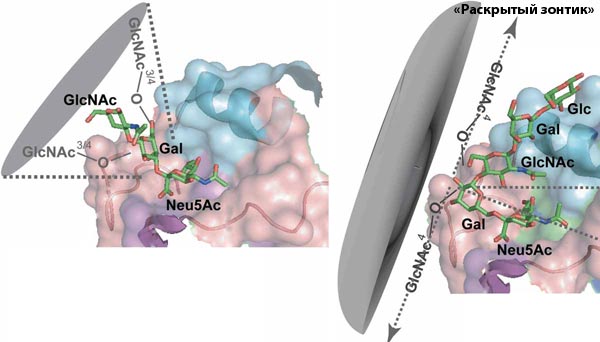
У человека вирусы гриппа поражают эпителиальные клетки слизистой оболочки респираторного тракта, а также бокаловидные клетки (секретирующие слизь), альвеолоциты и макрофаги [3, 4, 7]. Все эти клетки имеют на своей поверхности рецептор, с которым связывается вирусный гемагглютинин (рис. 1), — концевой остаток сиаловой, или N-ацетилнейраминовой, кислоты (Neu5Ac) (рис. 2), в составе полисахаридных цепочек, входящих в состав ганглиозидов и гликопротеинов. Концевой остаток сиаловой кислоты может связываться со следующим моносахаридом двумя способами: с помощью альфа2-3- или альфа2-6-связи (рис. 2) [14, 18].
Рис. 2. Структурные формулы сиаловой, или N-ацетилнейраминовой кислоты (Neu5Ac) и двух способов ковалентной связи со следующим моносахаридом (в данном случае — галактозой, Gal): альфа2-3- или альфа2-6-связью
Клетки эпителия верхних отделов респираторного тракта человека содержат, в основном, альфа2-6-сиалозиды; нижних отделов — альфа2-3-сиалозиды (рис. 3). Поэтому эпидемические штаммы вирусов гриппа, имея альфа2-6-специфичность, легко репродуцируются в верхних отделах респираторного тракта человека, активно выделяются в окружающую среду при речи, чихании, кашле и эффективно заражают других людей капельно-воздушным путем.
Варианты вируса гриппа А, адаптированные к птицам, имеют альфа2-3-специфичность (рис. 3). Концевые альфа2-3-сиалозиды содержатся у птиц, в основном, на поверхности эпителиальных клеток слизистой кишечника, поэтому у птиц грипп протекает в форме энтерита; вирус выделяется во внешнюю среду с фекалиями, а заражение происходит алиментарным путем. Альфа2-3-специфичность птичьих вариантов вируса гриппа А объясняет их неспособность эффективно поражать эпителий верхних отделов респираторного тракта человека и, как следствие, — передаваться капельно-воздушным путем в человеческой популяции. Вместе с тем, если высоковирулентный вирус гриппа А птиц каким-либо образом сумел вызвать продуктивную инфекцию в человеческом организме, то он будет эффективно поражать нижние отделы респираторного тракта, становясь причиной тяжелой первичной вирусной пневмонии (по данным ВОЗ, в 60% случаев — летальной).
Эпителиоциты свиней одновременно содержат и альфа2-6-, и альфа2-3-сиалозиды (рис. 3), поэтому в организме могут одновременно циркулировать и эпидемические, и птичьи варианты вируса гриппа А. Вследствие этого в свиных популяциях могут, во-первых, формироваться реассортанты 4 человеческих и птичьих штаммов с новыми биологическими свойствами; во-вторых, селектироваться штаммы со смешанной альфа2-6/альфа2-3-специфичностью. Именно такой смешанной альфа2-6/альфа2-3-специфичностью обладают штаммы пандемического вируса гриппа А(H1N1) swl, и, как следствие, они обладают способностью распространяться капельно-воздушным путем и вызывать тяжелые пневмонии [13–15, 18, 19, 23].
Рис. 3. Сиалозиды-рецепторы вирусов гриппа А на поверхности эпителиоцитов людей (альфа2-6 — на слизистой верхних, альфа2-3 — на слизистой нижних отделов респираторного тракта), свиней (альфа2-6/альфа2-3-смесь на слизистой респираторного тракта) и птиц (альфа2-3 — на слизистой кишечника)
Инфицирование эпителиоцитов имеет следствием быстрый рост вирусной нагрузки, апоптоз, дегенерацию и некроз этого типа клеток с последующим развитием токсических и токсико-аллергические реакций. У людей характерно повреждение клеток цилиндрического эпителия трахеи и бронхов. Главным звеном в патогенезе гриппа А является поражение сосудистой и нервной систем, возникающее вследствие токсического действия вируса. При этом одним из основных механизмов влияния вируса гриппа А на сосудистую систему является образование активных форм кислорода, которые взаимодействуют с фосфолипидами клеточных мембран, вызывая в них процесс перекисного окисления липидов, нарушение мембранного транспорта и барьерных функций, способствуя дальнейшему развитию вирусной инфекции. Лизосомальные ферменты дополнительно повреждают эпителий капилляров, базальную мембрану клеток, что способствует распространению гриппозной инфекции и виремии. Повышение проницаемости сосудов, ломкость их стенок, нарушение микроциркуляции является причиной возникновения геморрагических проявлений — от носовых кровотечений до геморрагического отека легких и кровоизлияний в вещество головного мозга. Циркуляторные расстройства, в свою очередь, вызывают поражения ЦНС: патоморфологическая картина характеризуется наличием лимфомоноцитарных инфильтратов вокруг мелких и средних вен, гиперплазией глиальных элементов и очаговой демиелинизацией, что свидетельствует о токсико-аллергической природе патологического процесса в ЦНС при гриппе [3–8, 23].
Важным фактором патогенеза при гриппе является продукция вирусного белка PB1-F2, который вызывает апоптоз тканевых макрофагов легких и тем самым способствует развитию вторичных бактериальных пневмоний (у современного пандемического варианта вируса гриппа А(H1N1) swl продукция PB1-F2, к счастью, отсутствует, что снижает — но не отменяет! — вероятность развития вторичных пневмоний, оставляя в силе опасность первичных вирусных пневмоний — см. далее) [2, 26].
Клиническая картина гриппа у людей
Начало острое, с озноба, быстрого повышения температуры до высоких цифр, резкого нарастания симптомов интоксикации. Температура достигает максимальных значений (39,0–40,0 °С) в первые сутки заболевания. В этот же период нарастают признаки интоксикации: озноб, сильная головная боль, головокружение, миалгии, артралгии, выраженная слабость. При внешнем осмотре: лицо гиперемировано, одутловато, сосуды склер инъецированы, определяется гиперемия конъюнктив, цианоз губ и слизистой оболочки ротоглотки, возможны точечные геморрагии на мягком небе. Цианоз вообще является важным симптомом при гриппе: следует обращать внимание не только на цианоз губ, но и на цианотичный оттенок язычка, миндалин, небных дужек на фоне яркой гиперемии слизистой ротоглотки; слизистая оболочка мягкого неба также имеет цианотичный оттенок, хорошо видна мелкая зернистость, инъекция сосудов и мелкоточечные геморрагические элементы; на задней стенке глотки — умеренная гиперплазия лимфоидной ткани.
Локализация головной боли: в лобно-височной области и в глазных яблоках (при легком надавливании на них или при их движении). Нередко определяются менингеальные знаки, которые постепенно исчезают с уменьшением интоксикации и снижением температуры тела. Диапазон клинических проявлений со стороны нервной системы достаточно широкий: от функциональных расстройств до серозных менингитов и тяжелых менингоэнцефалитов.
При развитии вторичных пневмоний на фоне вирусной инфекции определяются признаки бактериального воздействия, подтверждаемого обнаружением в мокроте бактерий Streptococcus pneumoniae, Staphylococcus aureus и др. Как правило, вторичная пневмония развивается после 5–7 сут гриппа и характеризуется повторным подъемом температуры до фебрильных значений, усилением кашля, появлением слизисто-гнойной мокроты, часто с прожилками крови, рентгенологически — очаговыми и очагово-сливными инфильтратами, нередко с признаками деструкции и абсцедирования. Позже 10 сут пневмония имеет, как правило, бактериальную этиологию и чаще всего связана с грамотрицательной микрофлорой.
Одним из главных факторов, способствующих тяжелому течению гриппа, является сопутствующая патология. В частности, у пациентов, умерших в период двух последних эпидсезонов 2009–2011 гг., преобладали болезни сердца и сосудов, сахарный диабет, метаболический синдром (ожирение), алкоголизм и табакокурение. Особую группу риска составляют беременные, у которых пневмония может развиваться стремительно, а потому они требуют особого внимания клиницистов и безотлагательной терапии.
Литература
М. Ю. Щелканов, доктор биологических наук, доцент
Л. В. Колобухина, доктор медицинских наук, профессор
Д. К. Львов, доктор медицинских наук, профессор, академик РАМН
1 Для сравнения: в результате военных действий за 5 лет Первой мировой войны (1914–1918 гг.) погибли 8,3 млн человек.
3 Эпизоотия — процесс распространения инфекционного заболевания в популяциях животных.
4 Вирус гриппа А имеет геном, состоящий из 8 отдельных молекул РНК. Реассортацией называется формирование штамма, у которого источником различных генетических сегментов стали различные родительские штаммы, одновременно инфицировавшие одну и ту же клетку.
Эпидемиология гриппа. Распространенность гриппа. Морфология вирусов гриппа.
Грипп [от фр. gripper, хватать], или инфлюэнца [от итал. influenza di freddo, влияние холода], — острая инфекция, проявляющаяся поражениями дыхательного тракта, непродолжительной лихорадкой, упадком сил, головной болью, миалгиями и др.
Резервуар гриппа — инфицированный человек (больные и бессимптомные носители). Больной становится заразным за 24 ч до появления основных симптомов и представляет эпидемическую опасность в течение 48 ч после их исчезновения.
Грипп регистрируют повсеместно, рост заболеваемости наблюдают в холодные месяцы. Эпидемии гриппа развиваются с интервалом 2-3 года.
Передача возбудителя гриппа происходит воздушно-капельным путём. Наиболее восприимчивы дети и лица преклонного возраста. Вирусы гриппа чувствительны к действию высоких температур, высушиванию, инсоляции и УФ-облучению. Также они лабильны к действию эфира, фенола, формальдегида и других веществ, денатурирующих белки.

Морфология вирусов гриппа
Суперкапсид вируса гриппа образован липидным бислоем, который пронизывают гликопротеиновые шипы (спикулы), определяющие гемагглютинирующую (Н) либо нейраминидазную (N) активность. Репликация ортомиксовирусов первично реализуется в цитоплазме инфицированной клетки; синтез вирусной РНК происходит в ядре.
• Гемагглютинин обусловливает проникновение вирусов гриппа в клетки в результате слияния с мембраной клетки и мембранами лизосом. AT к нему обеспечивают защитный эффект. Нейраминидаза распознаёт и взаимодействует с рецепторами, содержащими N-ацетилнейраминовую кислоту, то есть приводит к проникновению вируса, а также, отщепляя нейраминовую кислоту от дочерних вирионов и клеточной мембраны, к выходу вирусов из клеток.
• Семь сегментов вирусного генома кодируют структурные белки, восьмой — неструктурные белки NS1 и NS2 вируса гриппа, существующие только в инфицированных клетках. Основные из них — матриксный (М) и нуклеопротеидный (NP) белки. В меньших количествах присутствуют внутренние белки (P1, P2, Р3), участвующие в этапах транскрипции и репликации вируса гриппа.
• М-белок вирусов гриппа играет важную роль в морфогенезе вирусов и защищает геном, окружая нуклеокапсид. Белок NP выполняет регуляторные и структурные функции. Внутренние белки являются ферментами: Р1 — транскриптаза, Р2 — эндонуклеаза, Р3 — репликаза.
Штаммы вируса гриппа. Антигены вируса гриппа. Строение вируса гриппа.
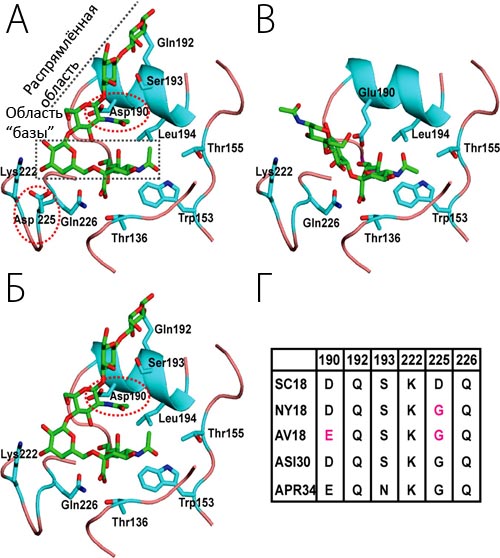
Вирулентные штаммы вируса гриппа, в отличие от невирулентных, обязательно активизируются за счет расщепления протеазами. Гемагглютинины вируса гриппа расщепляются внутриклеточно и потому способны инфицировать клетки различных тканей и вызывать системную инфекцию. Ортомиксовирусы с нерасщепленным НА имеют низкую инфекционную активность. Инфекционная форма вируса с расщепленным НА в организме млекопитающих образуется только в клетках, выстилающих дыхательный тракт. При отсутствии трипсина в культуре клеток расщепляется НА только вирулентных штаммов вируса. Для вируса гриппа птиц установлена строгая корреляция между расщепляемостью НА, способностью размножаться в культуре клеток и патогенностью для кур. Расщепление НА на субъединицы не является обязательным для сборки и выхода вирионов из клетки и проявления гемагглютинирующей активности. Полагают, что фактором, детерминирующим патогенность вируса гриппа птиц, является чувствительность НА к протеолитическому расщеплению.
В сайте расщепления НА у вирулентных штаммов вируса гриппа содержится несколько основных аминокислот, а у невирулентных — только один остаток аргинина. Таким образом, только участок из нескольких основных аминокислот у С-конца НА образует сайт узнавания для ферментов, ответственных за расщепление.

Основные функции НА: гемагглютинирующая активность вируса; прикрепление вирионов к клеткам-мишеням путем связывания с сиалосодержащими рецепторами; слияние вирусной и клеточной мембран. НА является основным специфическим антигеном вируса, определяющим (наряду с NA) подтип и вызывающим образование антител, нейтрализующих инфекционность вируса и его ГА-активность. НА играет главную роль в индукции протективного иммунитета при гриппе. В молекуле НА имеются 3-4 антигенных домена, изменения в которых определяют антигенный дрейф. NA является ферментом, катализирующим отщепление сиаловой кислоты от субстрата. Удаление сиаловой кислоты с НА облегчает его расщепление клеточными протеазами. Антитела к белку NA имеют вспомогательное значение в защите и нейтрализуют вирус лишь при высоком титре. Матриксный белок М - самый низкомолекулярный структурный белок. Он принимает участие в морфогенезе вириона и стабилизации его структуры. Нуклеопротеид (NP) - основной внутренний белок, формирующий субъединицы капсида. Белки NP и М являются типоспецифическими антигенами, общими для всех вирусов гриппа одного типа, и в этом отношении резко отличаются от высокодивергентных поверхностных белков. Они не вызывают образования протективных антител. Однако нуклеопротеин вируса гриппа — основной антиген, узнаваемый цитотоксическими Т-лимфоцитами. Аминокислотные последовательности 260—283 нуклеопротеина вируса гриппа А являются индукторами Т-клеточного ответа. Изменение антигенных свойств вируса гриппа — результат двух генетических процессов: антигенных дрейфа и шифта. Антигенный дрейф происходит в основном через накопление аминокислотных замен (точечных мутаций) в НА1. Замена одного аминокислотного остатка в эпитопе нарушает его связывание с соответствующими МАТ. Основные механизмы антигенного шифта — реассортация отдельных генов, возвращение в популяцию старых генов, прямые мутации, изменяющие специфичность к хозяину. NA может измениться независимо от НА. Вариабельность НА вируса гриппа А значительно выше вариабельности НА вируса гриппа В.
В процессе адаптации вируса гриппа к различным системам наблюдают изменение антигенной структуры НА, в основе которой лежит селекция мутантов с измененной рецепторсвязывающей специфичностью. Вирусы гриппа А и В человека, размноженные в КЭ, могут изменяться антигенно и претерпевать мутации в ГА (замена трех аминокислот в ГА). Вирус, размноженный в КЭ или в культуре клеток, защищает против культурального вируса более эффективно, чем против полевого вируса, выделенного от человека.
Один и тот же клинический изолят вируса гриппа при размножении в культуре клеток животных (МДСК) или в КЭ имеет разные антигенные свойства, что связано с заменой одной-двух аминокислот в разных участках молекулы НА. Особенно заметные изменения антигенности в молекуле НА наблюдали при замене аминокислотных остатков 187 и 189 в процессе адаптации вируса к КЭ. Изменение сайта гликозилирования может сопровождаться изменением антигенности и вирулентности вируса. Состав углеводов гемагглютинина вирусов и гриппа птиц может изменяться в различных хозяйских клетках даже в течение одного пассажа. У вирусов гриппа птиц типа А идентифицировано девять нейраминидазных N-антигенов, обозначенных 1-9, и 13 гемагглютинирующих антигенов Н, обозначенных 1-13. Состав Н- и N-антигенов полевых изолятов зависит от вида птиц, места и времени вспышек гриппа. Свиньи восприимчивы к различным подтипам вируса гриппа А и, возможно, вместе с водоплавающей птицей являются главным резервуаром вируса гриппа в природе.
В пермиссивных клетках вирусный НА активируется путем расщепления на две части НА1 и НА2, которые остаются связанными дисульфидными связями. Вирионы прикрепляются к клеткам, активированным НА, соединяются с рецепторами сиаловой кислоты плазматической мембраны и входят в клетку эндоцитозом. После сплавления оболочки вируса и эндоплазматической мембраны транскрипционный комплекс освобождается и транспортируется в ядро, где происходит транскрипция и репликация РНК.
Так же, как у всех других вирусов с негативно-полярным РНК-геномом, геном ортомиксовирусов выполняет две функции: 1) матричную для синтеза мРНК и 2) матричную для синтеза позитивно-полярной промежуточной формы РНК, которая служит матрицей для синтеза потомства геномной РНК. Из 8 первичных транскриптов, синтезированных на 8 генных сегментах вирусов гриппа А и В, 6 являются моноцисторными и транслируются прямо в белки. Два других — подвергаются сплайсингу с образованием двух мРНК, которые транслируются с различных рамок считывания с образованием двух белков. Вирусные белки синтезируются, используя клеточный механизм трансляции. Ортомиксовирусы используют несколько механизмов для увеличения кодирующей способности: сплайсинг мРНК, спаренную стоп-старт-трансляцию тандемных генов и сдвиг рамки считывания.
Репликация геномных РНК сегментов требует синтеза полноразмерных, положительной полярности РНК посредников, которые, в отличие от соответствующих мРНК транскриптов, не имеют КЭП-структуры на 5'-конце и поли (А) последовательности на З'-конце. Вновь синтезированный нуклеопротеин присоединяется к этим РНК, облегчая их использование в качестве матрицы для синтеза геномной РНК. Вирионы формируются почкованием, включая М белок и нуклеокапсид, которые встроены на плазматической мембране, в которую включены НА и NA. Механизмы копирования каждого РНК сегмента и включение их в каждый вирион неизвестны.
Хотя в результате реассортации генов теоретически могут возникать вирусы с любой комбинацией Н и N генов, однако только ограниченное число вирусов с определенной комбинацией Н и N генов оказались важными патогенами, имеющими право на существование. В том числе вызывающие респираторную патологию у людей: H1N1, H2N2, H3N2, H5N1 и, возможно, H3N8; у лошадей H7N7 и H3N8; у норок H10N4; у котиков H7N7 и H4N5; вирусы, часто выделяемые от свиней - H1N1 и H3N2. Главными возбудителями гриппа птиц являются вирусы с H5N2 и H7N1, хотя встречаются и другие варианты вируса гриппа А. Так как установлена резистентность диких видов птиц к вирусу гриппа А, им отводили роль резервуара вируса, прежде всего для домашней птицы. При экспериментальном заражении вирусом H1N5 дикие утки, как правило, не проявляли клинических признаков болезни, хотя вирус в их организме размножался и выделялся во внешнюю среду. Латентное инфицирование вирусом гриппа может иметь место у других домашних и диких животных.

Обзор
Такие маски в принудительном порядке носили в 1918—1919-х годах, во время крупнейшей пандемии гриппа, по современным подсчетам унесшей до 50 млн. человеческих жизней
Автор
Редакторы

Грипп
Среди наиболее трагических событий первой половины XX века — наряду с двумя мировыми войнами — выделяется пандемия гриппа 1918 года, ставшая не менее смертоносной, чем крупнейшие конфликты между людьми: в течение 18 месяцев около 50 млн. людей погибли от вирусной инфекции.
Гонки с гриппом
Ежегодно гриппом переболевает 5–15% всей мировой популяции. Наибольшему риску подвергаются дети до двух лет, пожилые люди от 65, а также больные астмой, диабетом или хроническими заболеваниями сердца вне зависимости от возраста. В то же время от болезни не застрахован никто — как показала пандемия 2009-го, когда больше всего пострадали как раз молодые люди с превосходной иммунной системой [4].
Гриппозный зоопарк
Это эстафета, которую ученые и органы здравоохранения уже так долго и с переменным успехом стараются прервать.

Ловец вирусов

— Что является самым главным в экологии вируса гриппа?
— Что еще мы узнали после встречи с птичьим гриппом?
— Болеют ли дикие птицы — природные носители вируса — гриппом?
— Как грипп передается между дикими птицами?
— Это неизвестно. Один из способов это выяснить — исследование природных очагов гриппа. Яркий пример, который наблюдаем мы с коллегами — залив Делавэр на востоке США. Птицы побережья во время сезонной миграции на север в мае останавливаются там, чтобы покормиться яйцами мечехвостов. По-видимому, в этом месте собирается критическая масса птиц, многие из которых еще не являются носителями гриппа, — что и становится определяющим фактором, позволяющим инфекции быстро распространяться. Есть и другие подобные места — в Австралии, Канаде, в других регионах. Потенциально подходит любое место массового скопления и кормления этой водной братии.
— Сколько разновидностей вируса гриппа циркулирует в природе?
— Над чем вы сейчас работаете?
Это исследование, в высшей степени ориентированное на практику. Используя наши диагностические методы, можно будет оперативно объявлять о появлении новых штаммов вируса и проводить грамотные кампании по вакцинации.

Рисунок 3. Во время пандемии поцелуи возможны только с защитой
— До сих пор птичий вирус гриппа H5N1 был не слишком активен, заражая людей. Какова вероятность, что в будущем эта его способность значительно возрастет?
— Вероятность пандемии вируса H5N1 значительно меньше, чем любого из H1, H2 или H3, потому что H5 далеко не так заразен для людей. Но если назвать вирус, который нам хотелось бы видеть пандемичным меньше всего, это будет как раз H5 из-за высокой смертности, причиной которой он становится. Одна из отличительных черт высокопатогенного вируса — молекулярные особенности строения гемагглютинина, позволяющие развиться системной инфекции, а не инфекции только дыхательных путей, как обычно [2]. Такая опасная способность числится только за вирусами H5 и H7.
У H5N1 было десять лет, чтобы переменить хозяина, а значит, это не одна-две и не три-четыре замены, которые должны произойти в нем, чтобы стать патогеном человека, а значительно больше. Есть примеры, когда для перемены хозяина вирусу требуется не одно десятилетие, несмотря на плотный контакт между видами, однако сейчас понятно хотя бы, что H5N1 не может передаться людям легко. Впрочем, совсем исключить эту вероятность нельзя, и если что-то и мешает нам спать спокойно по ночам, — то это вирус H5.
— Что, на ваш взгляд, наиболее удивительно в вирусе гриппа?
— Не устаю удивляться, как мало мы про него знаем, несмотря на то, что так давно и так активно изучаем. Это же сравнительно простые вирусы, — но до сих пор так и не понятно, что позволяет им периодически преодолевать межвидовые барьеры.

Обзор
Автор
Редактор
Вирус гриппа

Рисунок 1. Вирус гриппа. Слева — электронная микрофотография вирионов гриппа, увеличение ≈100 000 раз. Справа — молекулярное строение вируса гриппа; из 11 вирусных белков, закодированных в геноме, состоящем из нескольких фрагментов РНК, подписаны только гликопротеины оболочки гемагглютинин ( HA ) и нейраминидаза ( NA ).
Впервые появившись в XVI веке, грипп вызывает регулярные эпидемии и пандемии, причём с серотипом А связывают эпидемии с промежутками 2–3 года (наиболее крупные эпидемии и пандемии сведены в таблицу), а с серотипом B — каждые 4–6 лет. Серотип С эпидемий не вызывает; исключение составляют единичные вспышки у детей и ослабленных людей.
| Год | Серотип | Распространение |
|---|---|---|
| 1889–1890 | H2N8 | Тяжёлая эпидемия |
| 1900–1903 | H3N8 | Умеренная эпидемия |
| 1918–1919 | H1N1 | Тяжёлая пандемия [1] |
| 1933–1935 | H1N1 | Средняя эпидемия |
| 1946–1947 | H1N1 | Средняя эпидемия |
| 1957–1958 | H2N2 | Тяжёлая пандемия |
| 1968–1969 | H3N2 | Умеренная пандемия |
| 1977–1978 | H1N1 | Средняя пандемия |
| 1995–1996 | H1N1 и H3N2 | Тяжёлая пандемия |
Читайте также: